
Glucosinolates in Wild-Growing Reseda spp. from Croatia



Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
{kind=link}
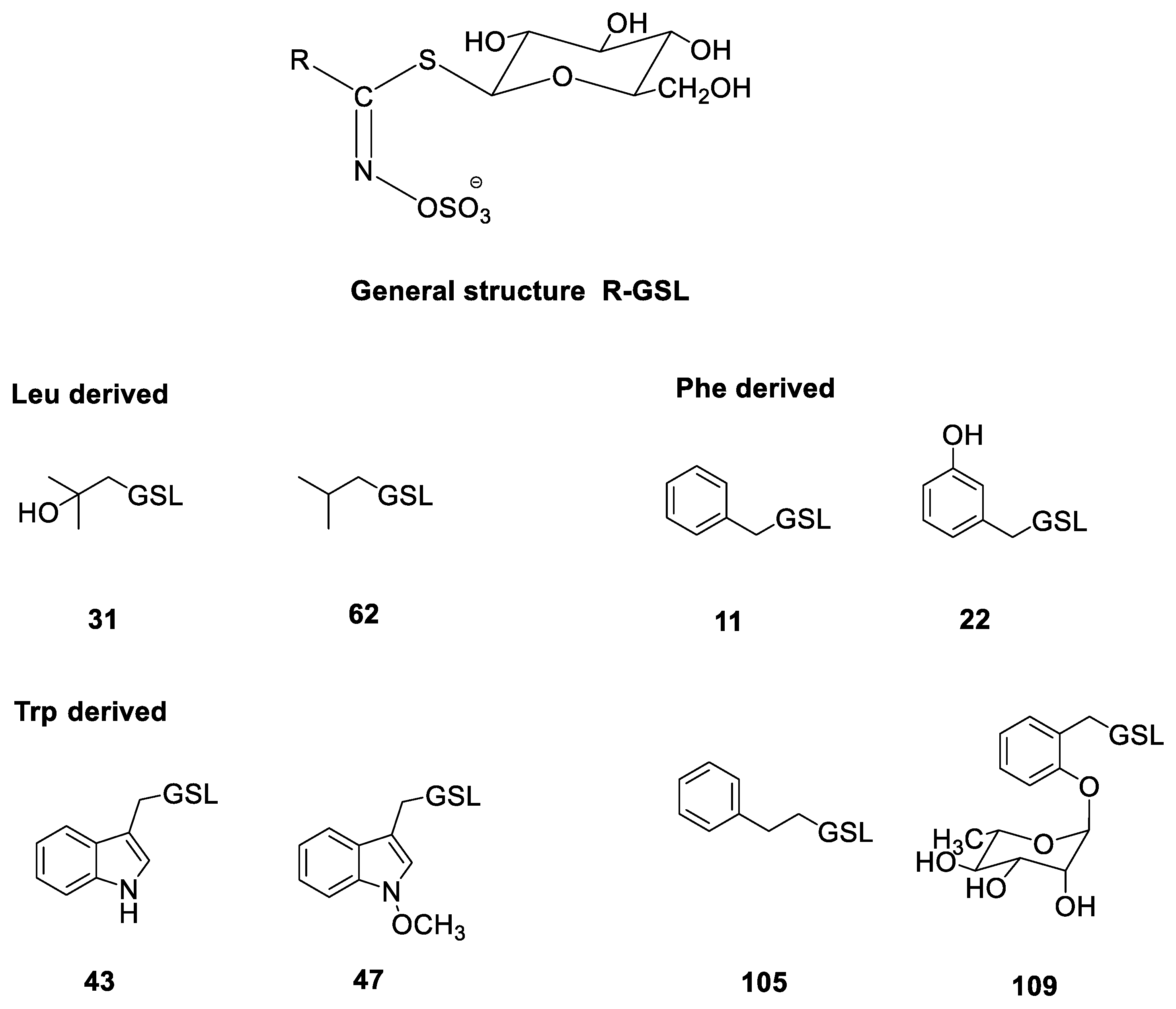
| No. * | Identified Glucosinolate | tR (min) | [M + Na]+ | Plant Tissue (μmol g−1 DW) | |||
|---|---|---|---|---|---|---|---|
| Reseda alba | Flower | Leaf | Stem | Root | |||
| Leu-derived | |||||||
| 31 | Glucoconringiin | 1.64 | 334 | 52.72 ± 2.22 | 6.48 ± 0.51 | 25.29 ± 1.13 | 19.70 ± 1.89 |
| 62 | Isobutyl GSL | 5.30 | 318 | 1.00 ± 0.24 | 0.13 ± 0.04 | 0.13 ± 0.05 | 1.13 ± 0.22 |
| Phe-derived | |||||||
| 105 | Gluconasturtiin | 7.93 | 366 | n.d. | 0.10 ± 0.03 | 1.20 ± 0.37 | 107.77 ± 2.83 |
| Trp-derived | |||||||
| 43 | Glucobrassicin | 7.21 | 391 | 1.64 ± 0.32 | 3.50 ± 0.11 | 0.85 ± 0.10 | 1.69 ± 0.67 |
| 47 | Neoglucobrassicin | 9.34 | 421 | 0.32 ± 0.08 | 0.23 ± 0.02 | 0.55 ± 0.11 | 2.74 ± 0.38 |
| Reseda lutea | Flower | Leaf | Stem | Root | |||
| Phe-derived | |||||||
| 22 | Glucolepigramin | 5.22 | 368 | 8.66 ± 1.00 | n.d. | n.d. | n.d. |
| 11 | Glucotropaeolin | 6.51 | 352 | 1.64 ± 0.76 | 0.6 ± 0.09 | 5.67 ± 0.75 | 106.69 ± 3.04 |
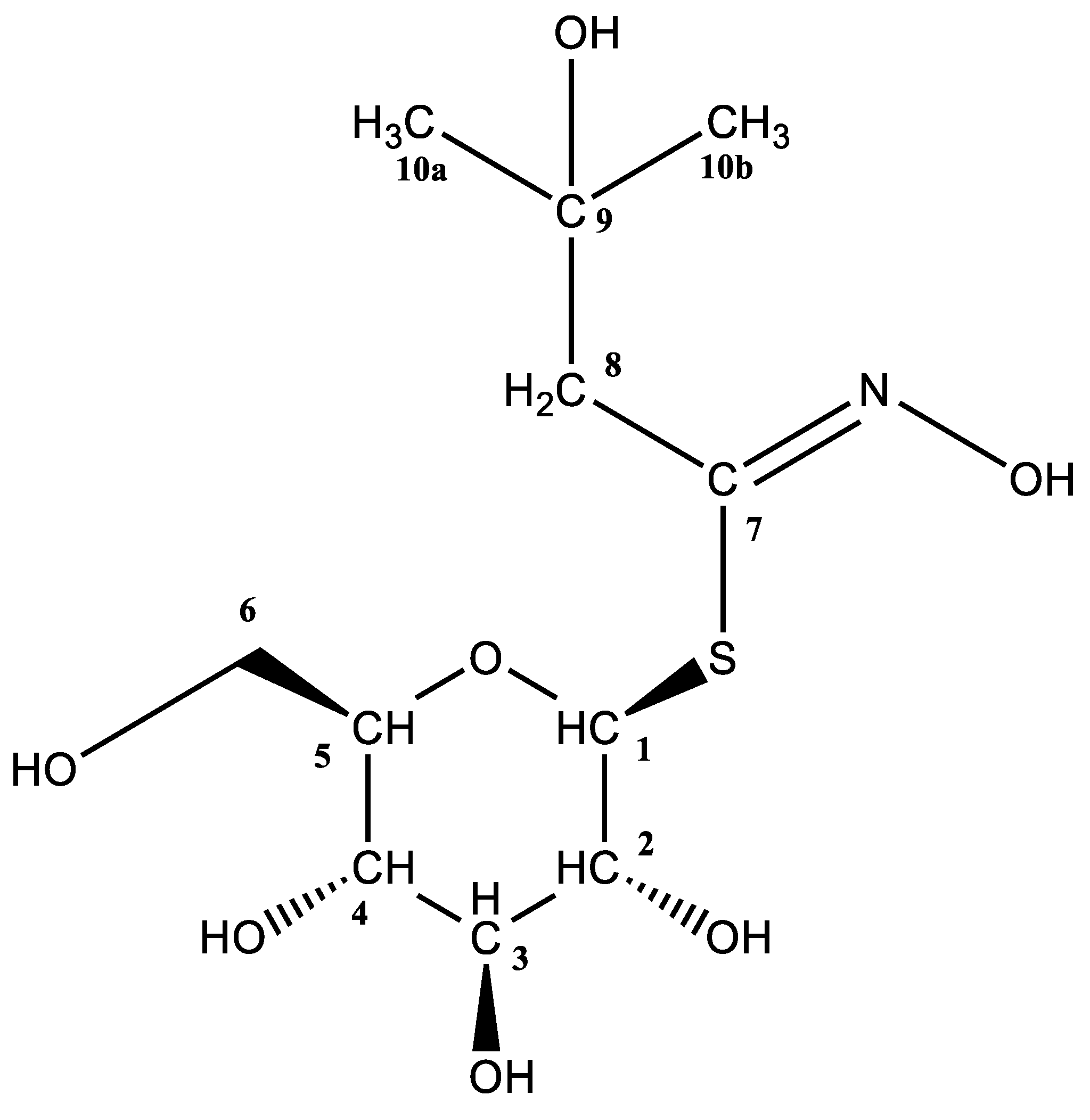
| 109 | 2-(α-L-Ramnopyranosyloxy)-benzyl GSL | 6.78 | 514 | 57.89 ± 3.19 | 20.50 ± 1.50 | 14.86 ± 1.86 | 8.10 ± 1.06 |
| Trp-derived | |||||||
| 43 | Glucobrassicin | 7.21 | 391 | 5.92 ± 0.34 | 0.76 ± 0.15 | 2.45 ± 0.25 | 3.26 ± 0.17 |
| Reseda phyteuma | Flower | Leaf | Stem | Siliquae | |||
| Phe-derived | |||||||
| 11 | Glucotropaeolin | 6.51 | 352 | n.d. | n.d. | tr | n.d. |
| 109 | 2-(α-L-Ramnopyranosyloxy)-benzyl GSL | 6.78 | 514 | 150.84 ± 3.52 | 164.37 ± 3.72 | 50.79 ± 1.23 | 123.93 ± 2.64 |
| 105 | Gluconasturtiin | 7.93 | 366 | n.d. | n.d. | 1.97 ± 0.21 | n.d. |
| Trp-derived | |||||||
| 43 | Glucobrassicin | 7.21 | 391 | 8.47 ± 0.18 | 4.79 ± 0.73 | 11.26 ± 0.77 | 3.13 ± 0.12 |
3. Materials and Methods
3.1. Materials and Reagents
3.2. Isolation and Chemical Analysis
3.2.1. Isolation of Desulfoglucosinolates
3.2.2. UHPLC-DAD-MS/MS Analysis
3.2.3. NMR Mesurements
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Stevens, P.F. Angiosperm Phylogeny Website. Version 14 July 2017. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 23 December 2022).
- Çilden, E.; Yıldırımlı, Ş. The impact of seed micromorphology in the subgeneric classification of the genus Reseda L. (Resedaceae) in Turkey. Microsc. Res. Tech. 2021, 84, 1992–2003. [Google Scholar] [CrossRef]
- Martín-Bravo, S.; Meimberg, H.; Luceño, M.; Märkl, W.; Valcárcel, V.; Bräuchler, C.; Vargas, P.; Heubl, G. Molecular systematics and biogeography of Resedaceae based on ITS and trnL-F sequences. Mol. Phylogenet. Evol. 2007, 44, 1105–1120. [Google Scholar] [CrossRef]
- Martín-Bravo, S.; Vargas, P.; Luceño, M. Is oligomeris (Resedaceae) indigenous to North America? Molecular evidence for a natural colonization from the Old World. Am. J. Bot. 2009, 96, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, T. (Ed.) Flora Croatica Database; Faculty of Science, University of Zagreb: Zagreb, Croatia, 2005; Available online: http://hirc.botanic.hr/fcd (accessed on 18 December 2022).
- Agerbirk, N.; Hansen, C.C.; Olsen, C.E.; Kiefer, C.; Hauser, T.P.; Christensen, S.; Jensen, K.R.; Ørgaard, M.; Pattison, D.I.; Lange, C.B.A.; et al. Glucosinolate profiles and phylogeny in Barbarea compared to other tribe Cardamineae (Brassicaceae) and Reseda (Resedaceae), based on a library of ion trap HPLC-MS/MS data of reference desulfoglucosinolates. Phytochemistry 2021, 185, 112658. [Google Scholar] [CrossRef]
- Bell, L. The biosynthesis of glucosinolates: Insights, inconsistencies, and unknowns. Ann. Plant Rev. Online 2019, 2, 969–1000. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E.; et al. Phytochemicals of Brassicaceae in plant protection and human health—Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Edger, P.P.; Heidel-Fischer, H.M.; Bekaert, M.; Rota, J.; Glöckner, G.; Platts, A.E.; Heckel, D.G.; Der, J.P.; Wafula, E.K.; Tang, M.; et al. The butterfly plant arms-race escalated by gene and genome duplications. Proc. Natl. Acad. Sci. USA 2015, 112, 8362–8383. [Google Scholar] [CrossRef]
- Leonti, M.; Nebel, S.; Rivera, D.; Heinrich, M. Wild gathered food plants in the European Mediterranean: A comparative analysis. Econ. Bot. 2006, 60, 130–142. [Google Scholar] [CrossRef]
- Gilbert, K.G.; Cooke, D.T. Dyes from plants: Past usage, present understanding and potential. Plant Growth Regul. 2001, 34, 57–69. [Google Scholar] [CrossRef]
- Abdallah, M.S.; De Witt, H.D.C. The Resedaceae—A taxonomical revision of the family (final instalment). Meded. Landbouwhogeschoo Wagening. 1978, 14, 308. Available online: https://edepot.wur.nl/287583 (accessed on 18 December 2022).
- Radulović, N.S.; Zlatković, D.B.; Ilić-Tomić, T.; Senerović, L.; Nikodinovic-Runic, J. Cytotoxic effect of Reseda lutea L.: A case of forgotten remedy. J. Ethnopharmacol. 2014, 153, 125–132. [Google Scholar] [CrossRef]
- Kumarasamy, Y.; Cox, P.J.; Jaspars, M.; Nahar, L.; Sarker, S.D. Screening seeds of Scottish plants for antibacterial activity. J. Ethnopharmacol. 2002, 83, 73–77. [Google Scholar] [CrossRef]
- Benmerache, A.; Berrehal, D.; Khalfallah, A.; Kabouche, A.; Semra, Z.; Kabouche, Z. Antioxidant, antibacterial activities and flavonoids of Reseda phyteuma L. Der Pharm. Lett. 2012, 4, 1863–1867. [Google Scholar]
- Susplugas, P.; Mongold, J.J.; Taillade, C.; Serrano, J.J. Anti-inflammatory and analgesic activity of Reseda phyteuma. Plantes Med. Phytother. 1993, 26, 375–382. [Google Scholar]
- Moghaddam, N.S.; Eryılmaz, M.; Altanlar, N.; Yıldırım, O. Antimicrobial screening of some selected Turkish medicinal plants. Pak. J. Pharm. Sci. 2019, 32, 947–951. [Google Scholar] [PubMed]
- Asadi-Samani, M.; Khaledi, M.; Khaledi, F.; Samarghandian, S.; Gholipour, A. Phytochemical properties and antibacterial effects of Salvia multicaulis Vahl., Euphorbia microsciadia Boiss., and Reseda lutea on Staphylococcus aureus and Acinetobacter baumanii. Jundishapur J. Nat. Pharm. Prod. 2019, 14, e63640. [Google Scholar] [CrossRef]
- Badridze, G.; Kacharava, N.; Chkhubianishvili, E.; Rapava, L.; Kikvidze, M.; Chigladze, L.; Chanishvili, S. Content of antioxidants in leaves of some plants of Tbilisi environs. Bull. Georg. Natl. Acad. Sci. 2013, 7, 105–111. [Google Scholar]
- Abdalrahman, K.S.; Güneş, M.G.; Shomali, N.; Işgör, B.S.; Yildirim, Ö. Screening effects of methanol extracts of Diplotaxis tenuifolia and Reseda lutea on enzymatic antioxidant defense systems and aldose reductase activity. Turk. J. Pharm. Sci. 2018, 15, 97–102. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel sources and biological potential. In Glucosinolates; Mérillon, J.-M., Ramawat, G.K., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–60. [Google Scholar]
- Burčul, F.; Generalić Mekinić, I.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzyme Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Gmelin, R.; Kjær, A. 2-Hydroxy-2-methylpropyl glucosinolate in Reseda alba. Phytochemistry 1970, 9, 599–600. [Google Scholar] [CrossRef]
- Olsen, O.; Sorensen, H. Isolation of glucosinolates and the identification of O-(α-L-rhamnopyranosyloxy)benzylglucosinolate from Reseda odorata. Phytochemistry 1979, 18, 1547–1552. [Google Scholar] [CrossRef]
- Ludwig-Müller, J.; Bennett, R.N.; Kiddle, G.; Ihmig, S.; Ruppel, M.; Hilgenberg, W. The host range of Plasmodiophora brassicae and its relationship to endogenous glucosinolate content. New Phytol. 1999, 141, 443–458. [Google Scholar] [CrossRef]
- Pagnotta, E.; Montaut, S.; Matteo, R.; Rollin, P.; Nuzillard, J.-M.; Lazzeri, L.; Bagatta, M. Glucosinolates in Reseda lutea L.: Distribution in plant tissues during flowering time. Biochem. Syst. Ecol. 2020, 90, 104043. [Google Scholar] [CrossRef]
- Mithen, R.; Bennett, R.; Marquez, J. Glucosinolate biochemical diversity and innovation in the Brassicales. Phytochemistry 2010, 71, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Kjær, A.; Gmelin, R. An isothiocyanate glucoside (glucobarbarin) of Reseda luteola L. Acta Chem. Scand. 1958, 12, 1693–1694. [Google Scholar] [CrossRef]
- Griffiths, D.W.; Deighton, N.; Birch, A.N.; Patrian, B.; Baur, R.; Städler, E. Identification of glucosinolates on the leaf surface of plants from the Cruciferae and other closely related species. Phytochemistry 2001, 57, 693–700. [Google Scholar] [CrossRef]
- Agerbirk, N.; Matthes, A.; Erthmann, P.Ø.; Ugolini, L.; Cinti, S.; Lazaridi, E.; Nuzillard, J.-M.; Müller, C.; Bak, S.; Rollin, P.; et al. Glucosinolate turnover in Brassicales species to an oxazolidin-2-one, formed via the 2-thione and without formation of thioamide. Phytochemistry 2018, 153, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Olsen, O.; Rasmussen, K.W.; Sørensen, H. Glucosinolates in Sesamoides canescens and S. pygmaea: Identification of 2-(α-L-arabinopyranosyloxy)-2-phenylethylglucosinolate. Phytochemistry 1981, 20, 1857–1861. [Google Scholar] [CrossRef]
- Olsen, O.; Sorensen, H. Glucosinolates and amines in Reseda media. Phytochemistry 1980, 19, 1783–1787. [Google Scholar] [CrossRef]
- Trabelcy, B.; Chinkov, N.; Samuni-Blank, M.; Merav, M.; Izhaki, I.; Carmeli, S.; Gerchman, Y. Investigation of glucosinolates in the desert plant Ochradenus baccatus (Brassicales: Resedaceae). Unveiling glucoochradenin, a new arabinosylated glucosinolate. Phytochemistry 2021, 187, 112760. [Google Scholar] [CrossRef] [PubMed]
- Đulović, A.; Popović, M.; Burčul, F.; Čikeš Čulić, V.; Marijan, S.; Ruščić, M.; Anđelković, N.; Blažević, I. Glucosinolates of Sisymbrium officinale and S. orientale. Molecules 2022, 27, 8431. [Google Scholar] [CrossRef]
- Montaut, S.; Read, S.; Blažević, I.; Nuzillard, J.-M.; Harakat, D.; Rollin, P. Glucosinolates of Lepidium graminifolium L. (Brassicaceae) from Croatia. Nat. Prod. Res. 2021, 35, 494–498. [Google Scholar] [CrossRef]
- Agerbirk, N.; Hansen, C.C.; Kiefer, C.; Hauser, T.P.; Ørgaard, M.; Lange, C.B.A.; Cipollini, D.; Koch, M.A. Comparison of glucosinolate diversity in the crucifer tribe Cardamineae and the remaining order Brassicales highlights repetitive evolutionary loss and gain of biosynthetic steps. Phytochemistry 2021, 185, 112668. [Google Scholar] [CrossRef] [PubMed]
- Agerbirk, N.; Pattison, D.I.; Mandáková, T.; Lysak, M.A.; Montaut, S.; Staerk, D. Ancient biosyntheses in an oil crop: Glucosinolate profiles in Limnanthes alba and its relatives (Limnanthaceae, Brassicales). J. Agric. Food Chem. 2022, 70, 1134–1147. [Google Scholar] [CrossRef]
- Wathelet, J.-P.; Iori, R.; Leoni, O.; Quinsac, A.; Palmieri, S.; Rollin, P. Guidelines for glucosinolate analysis in green tissues used for biofumigation. Agroindustria 2004, 3, 257–344. [Google Scholar]
- European Community. Oil seeds–determination of glucosinolates High Performance Liquid Chromatography. Off. J. Eur. Communities 1990, 170, 28. [Google Scholar]
| Amino Acid Precursor | Met | Leu | Phe/Tyr | Trp | References | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. * | 24S | 31 | 4 | 11 | 22 | 23 | 40S | 40R | 105 | 109 | 158 | 43 | 47 | |
| Caylusea abyssinica |  |  | | | [24,25] | |||||||||
| Reseda alba | |  | | | | | | [24,26,27,28] | ||||||
| R. complicata | | | [24] | |||||||||||
| R. crystallina | | [24] | ||||||||||||
| R. lutea | | | | | | [14,24,25,29] | ||||||||
| R. luteola | | | | | | [6,24,25,27,30,31,32,33] | ||||||||
| R. media | | | | | [24,34] | |||||||||
| R. odorata | | | | | [6,24,25,27,30] | |||||||||
| R. phyteuma 1 | | | | | | | | [24,30,35] | ||||||
| R. stricta | | | | [24,30] | ||||||||||
| R. suffruticosa | | [30] | ||||||||||||
| Sesamoides interrupta 2 | | | | | [24,35] | |||||||||
| Ochradenus baccatus | | | | | | | [36] | |||||||
-“Circumstantial evidence”—reasonable but not conclusive evidence of qualitative analysis; -“Present”—qualitative analysis performed using relevant analytic methods (standards, MS, and NMR); -Qualitative and quantitative analyses performed.Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đulović, A.; Tomaš, J.; Blažević, I. Glucosinolates in Wild-Growing Reseda spp. from Croatia. Molecules 2023, 28, 1753. https://doi.org/10.3390/molecules28041753
Đulović A, Tomaš J, Blažević I. Glucosinolates in Wild-Growing Reseda spp. from Croatia. Molecules. 2023; 28(4):1753. https://doi.org/10.3390/molecules28041753
Chicago/Turabian StyleĐulović, Azra, Josip Tomaš, and Ivica Blažević. 2023. "Glucosinolates in Wild-Growing Reseda spp. from Croatia" Molecules 28, no. 4: 1753. https://doi.org/10.3390/molecules28041753
APA StyleĐulović, A., Tomaš, J., & Blažević, I. (2023). Glucosinolates in Wild-Growing Reseda spp. from Croatia. Molecules, 28(4), 1753. https://doi.org/10.3390/molecules28041753