
Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
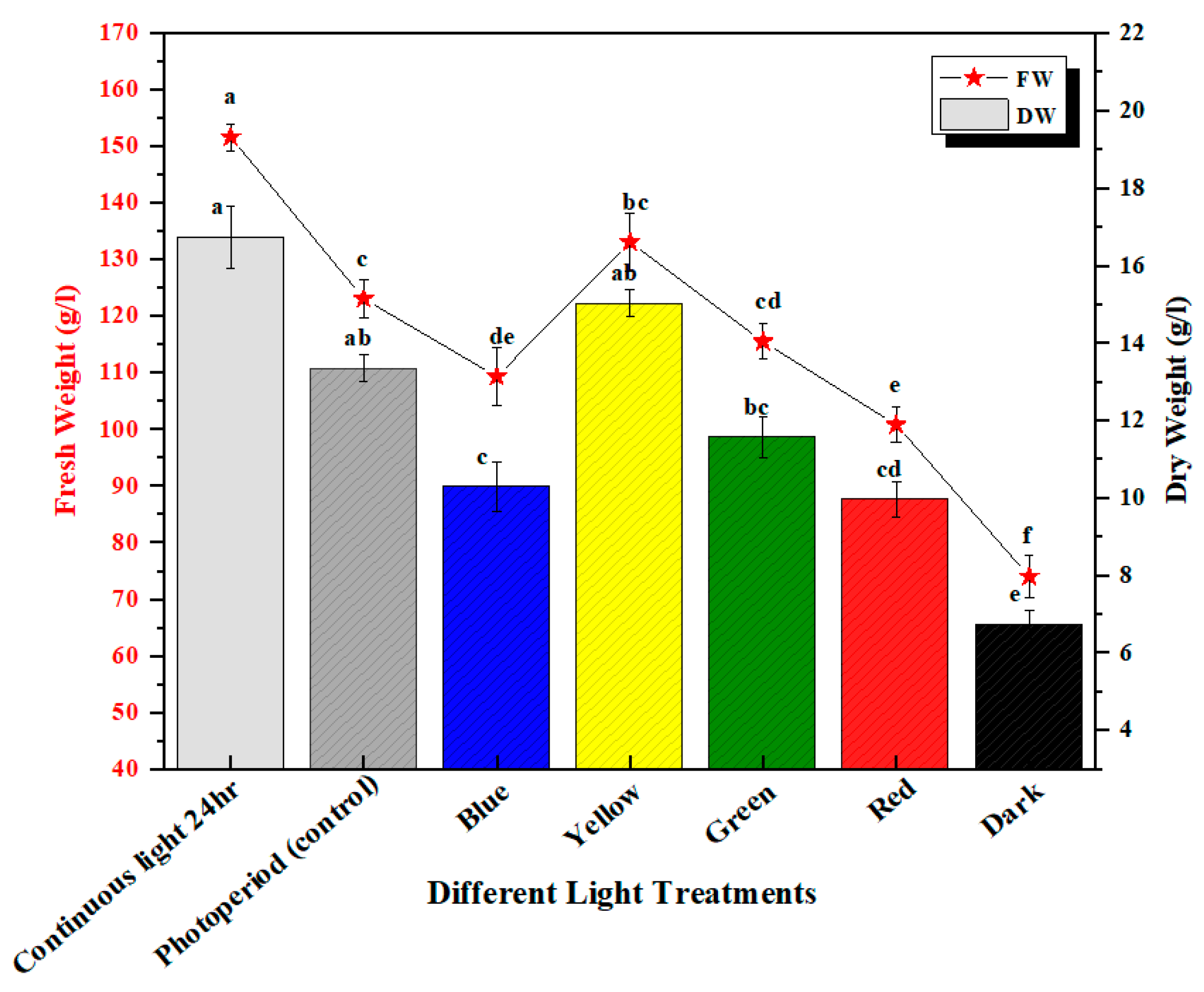
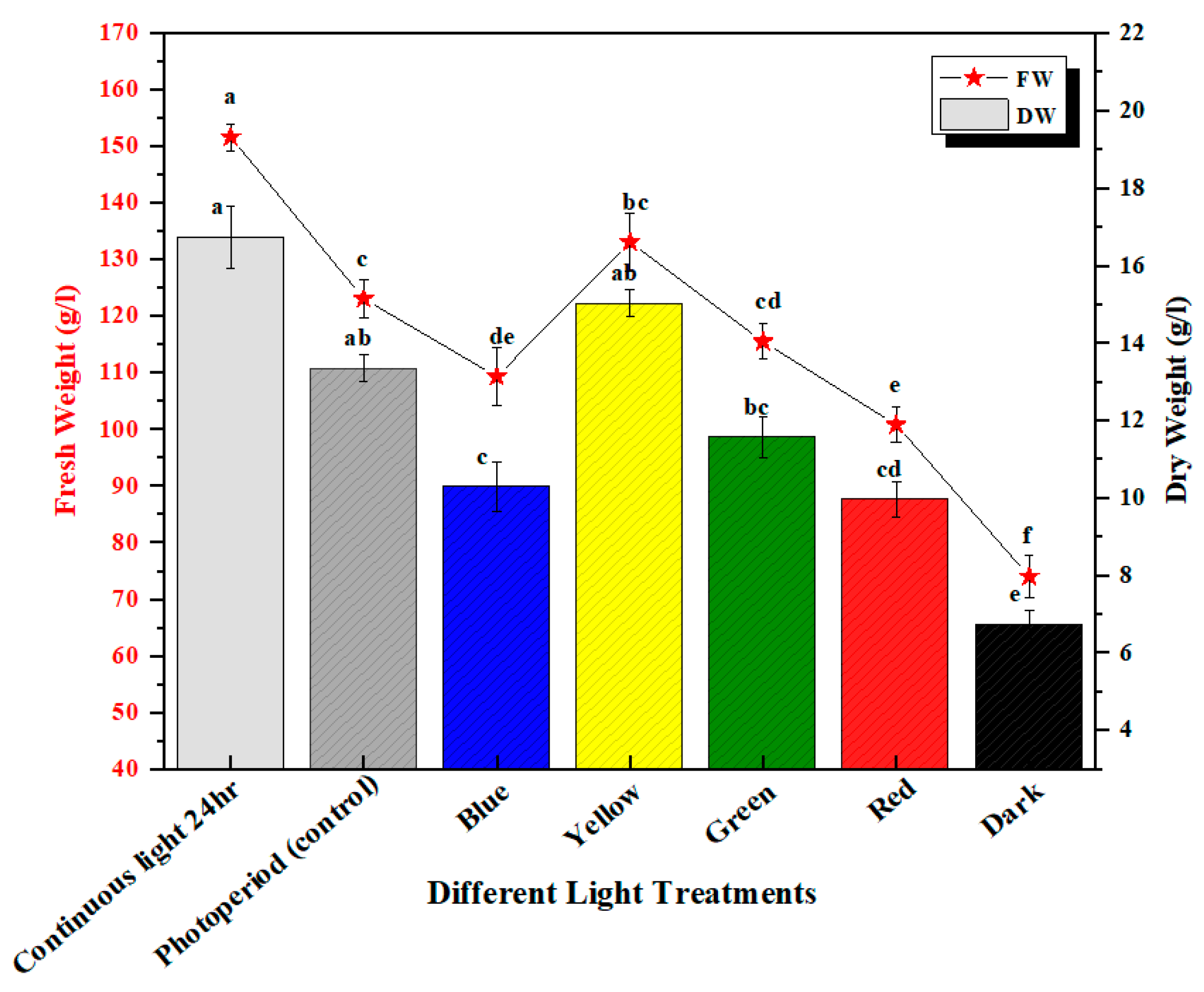
2.1. Trend in Biomass Accumulation
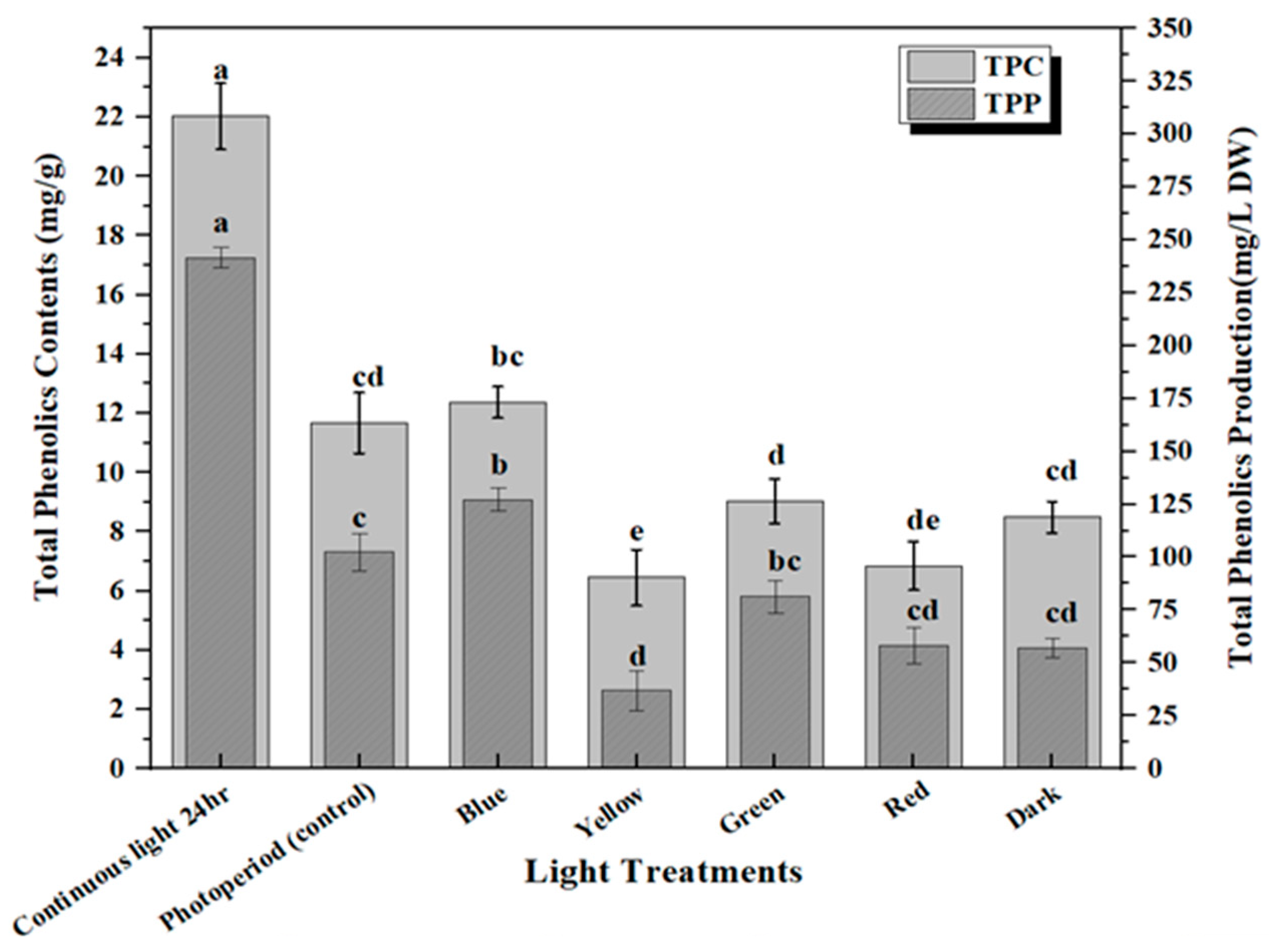
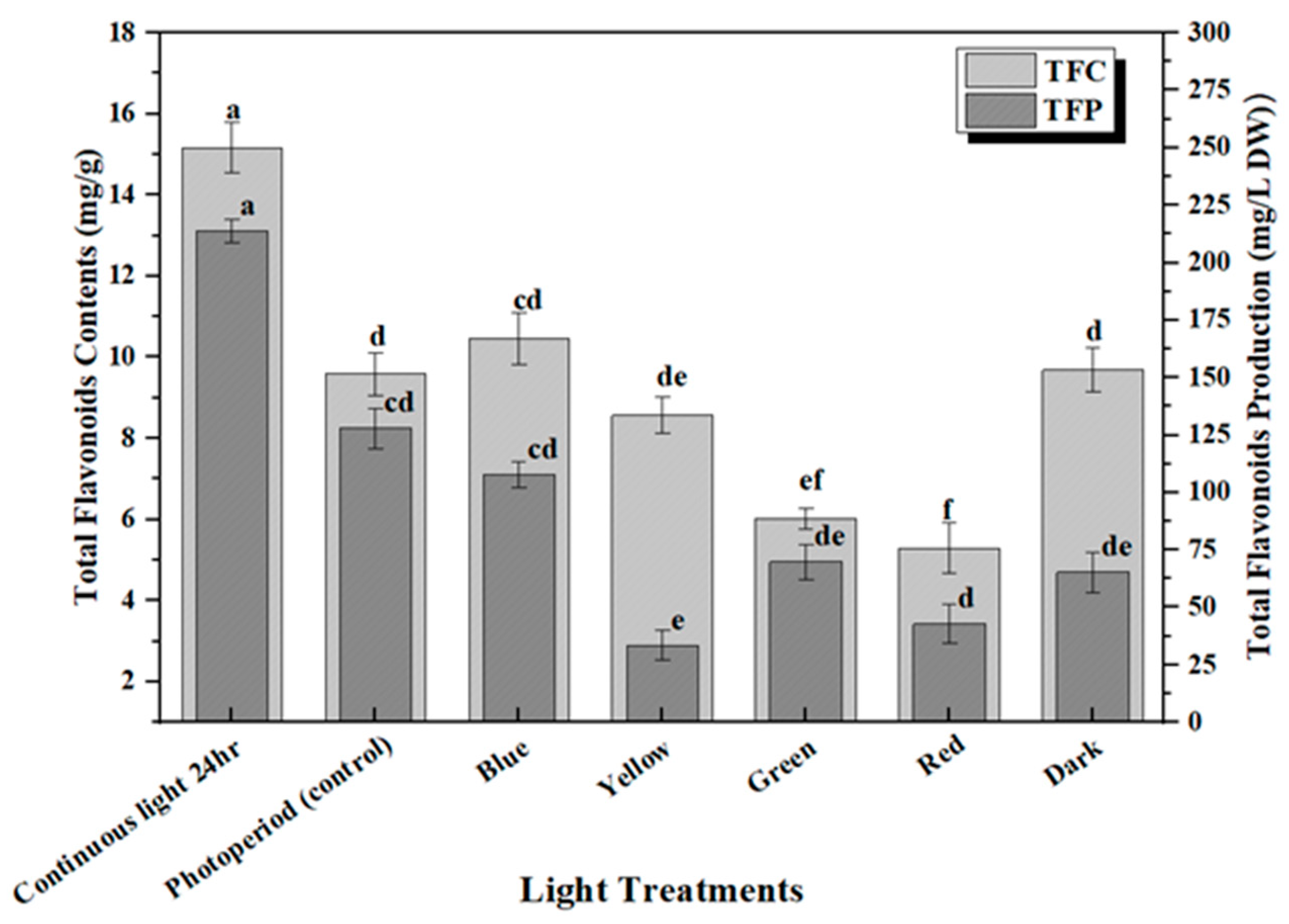
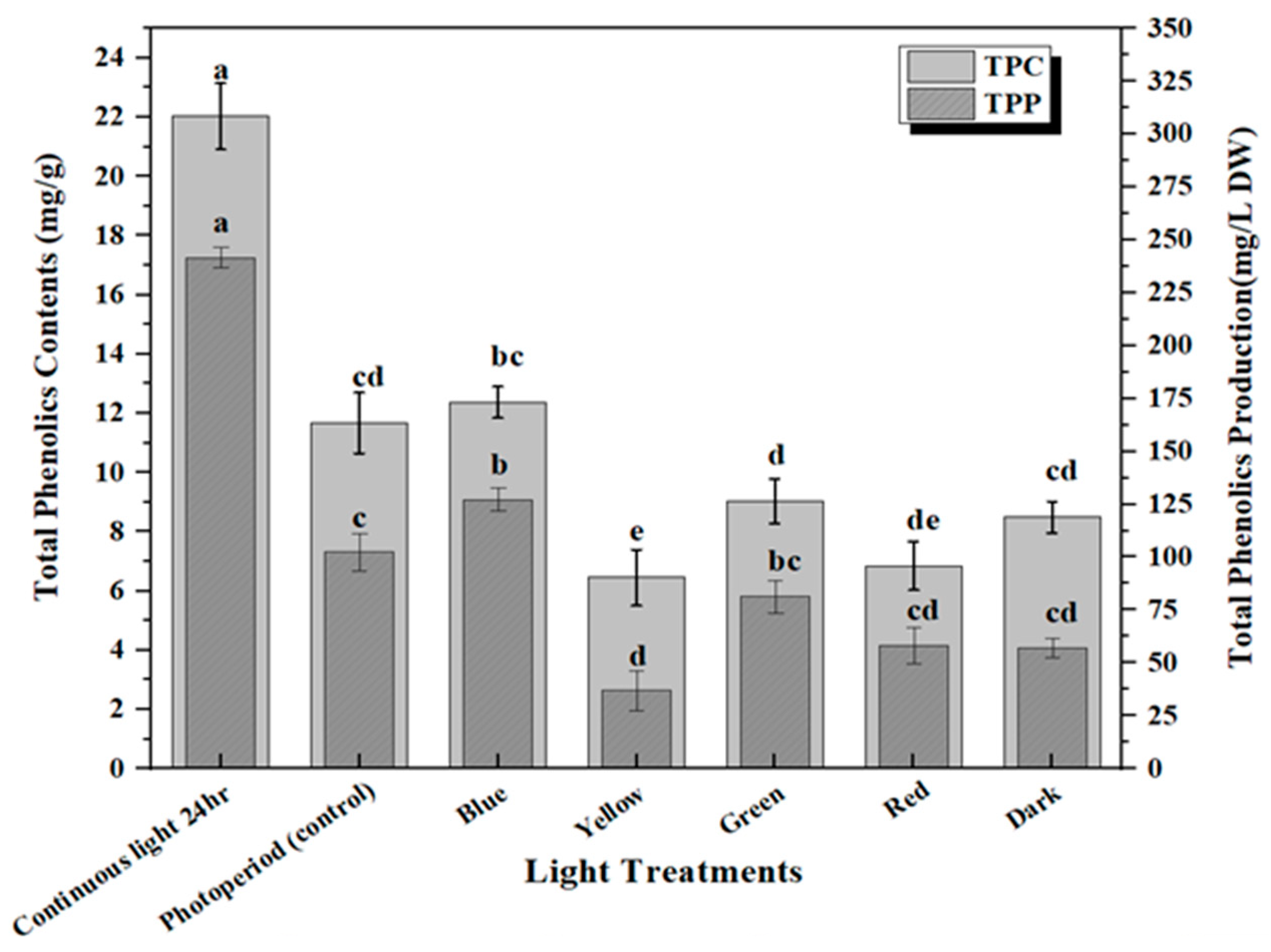
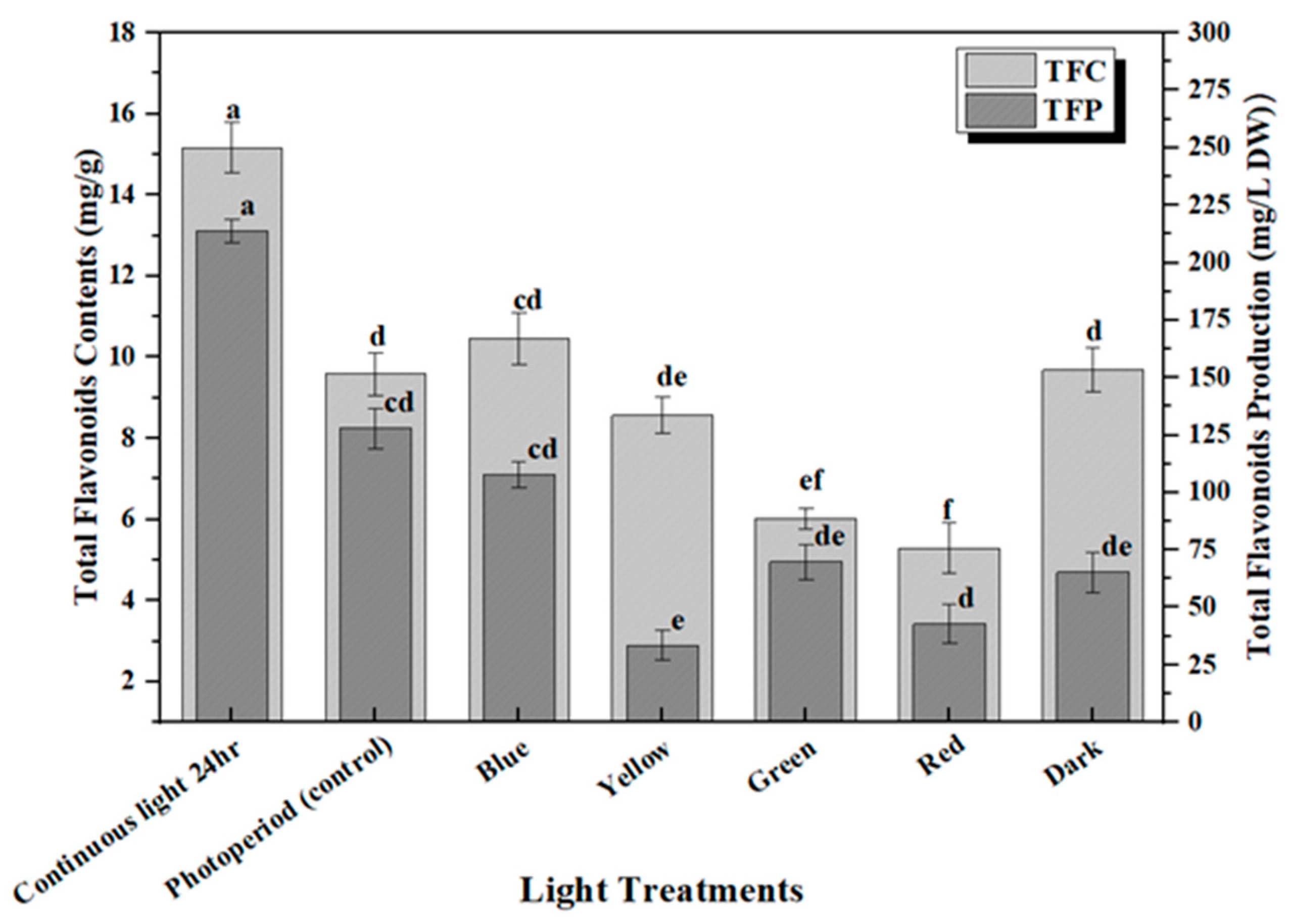
2.2. Accumulation of Phenolic and Flavonoid Contents
2.3. HPLC-Based Metabolite Quantificaton
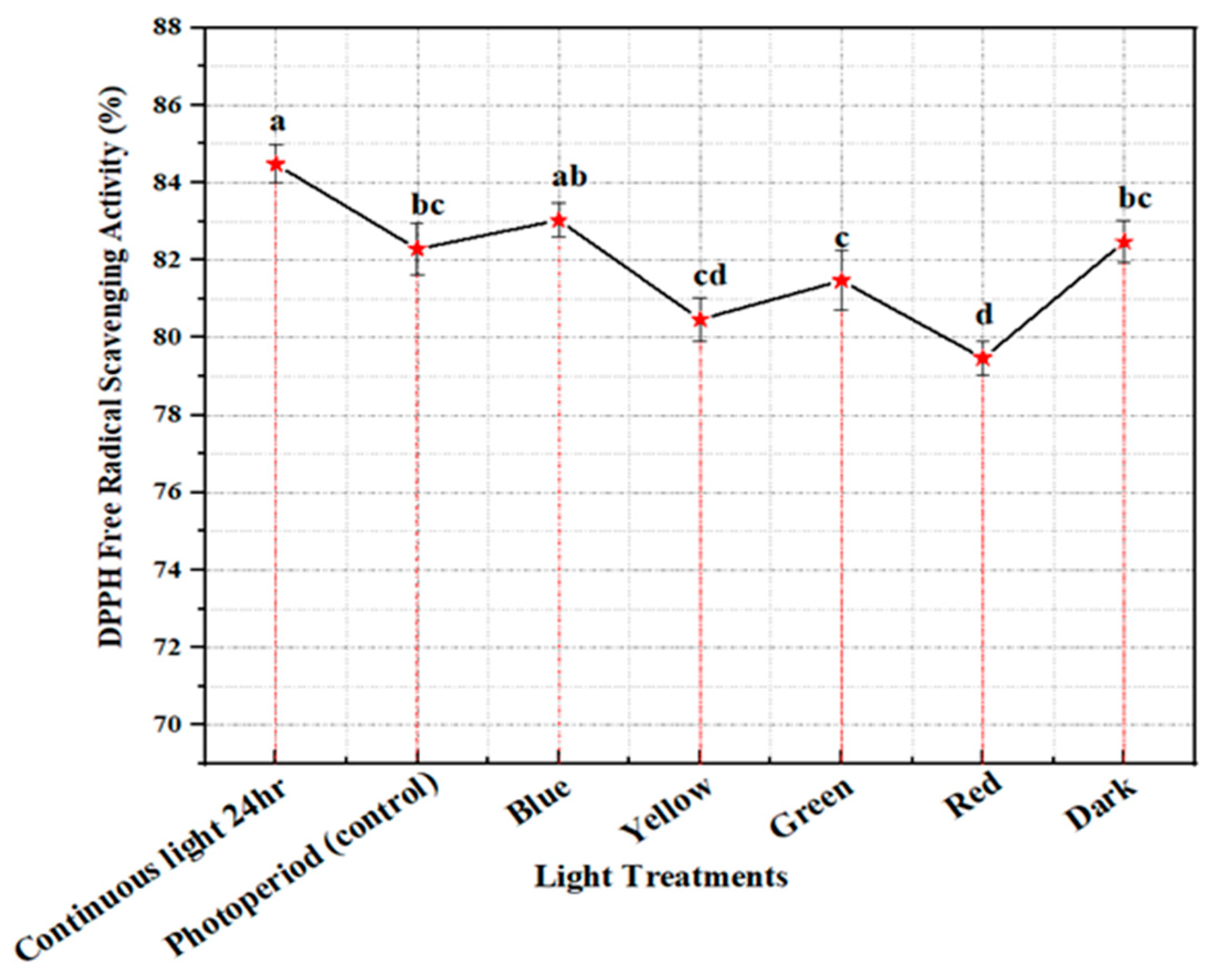
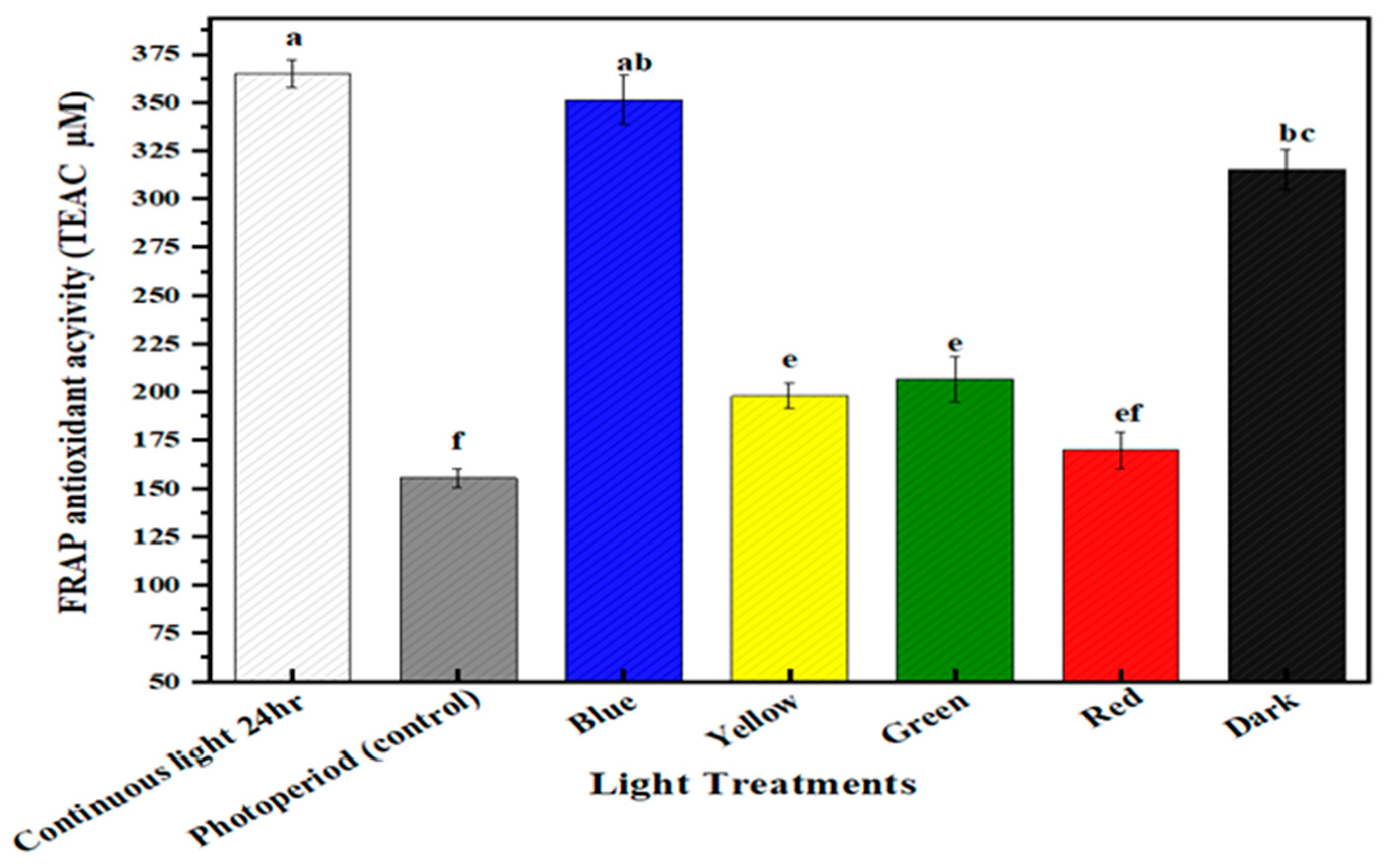
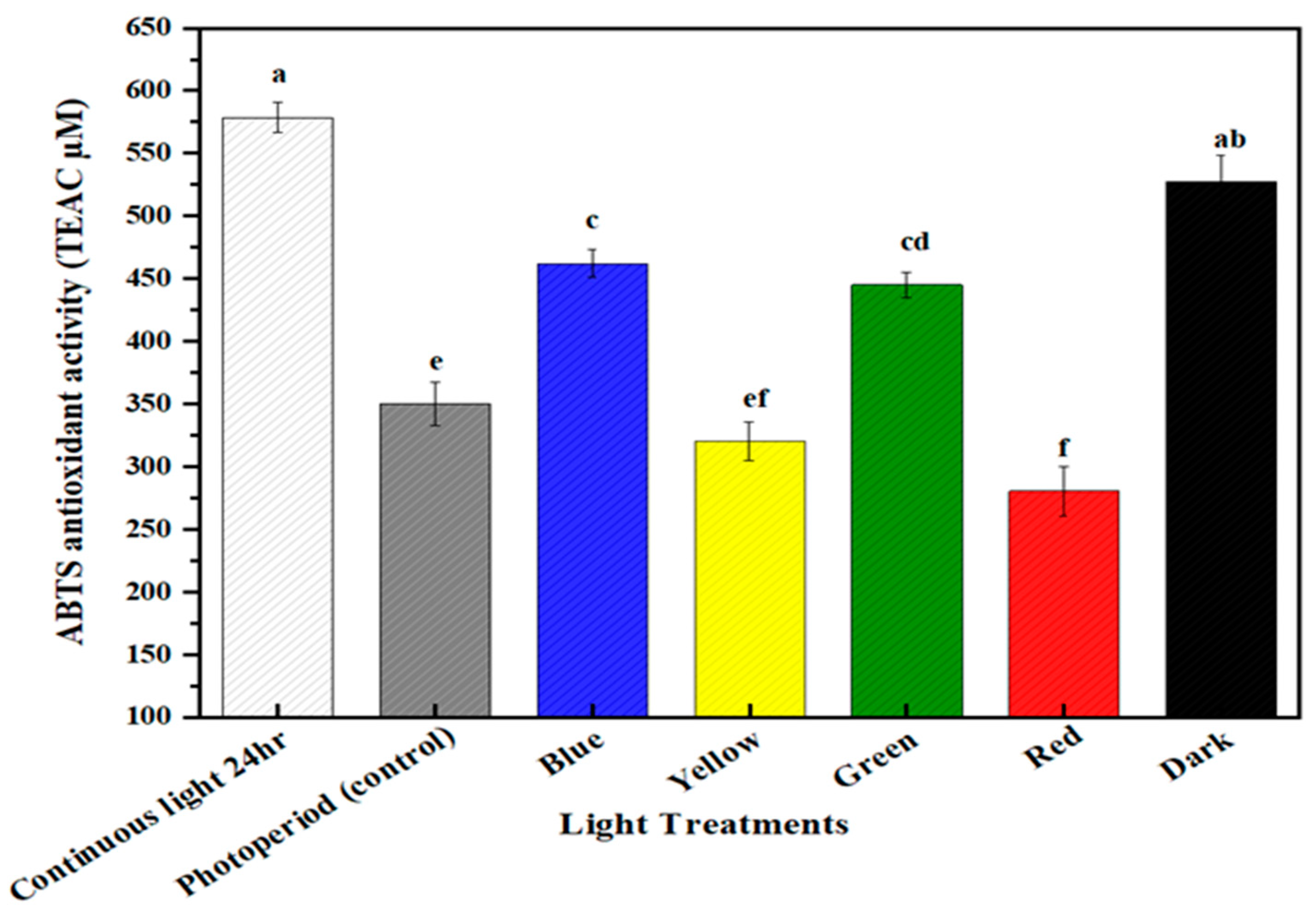
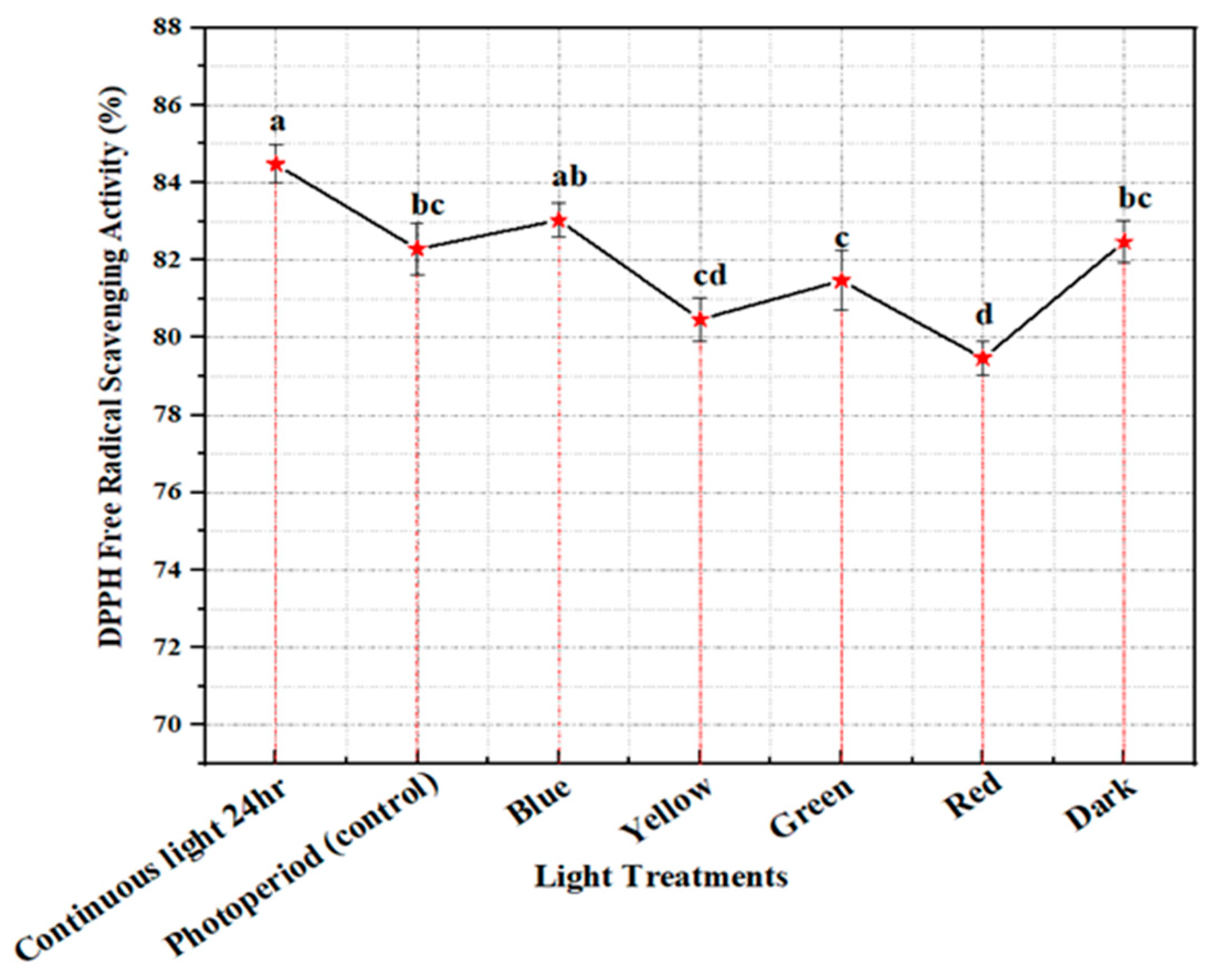
2.4. Free Radical Scavenging Activity
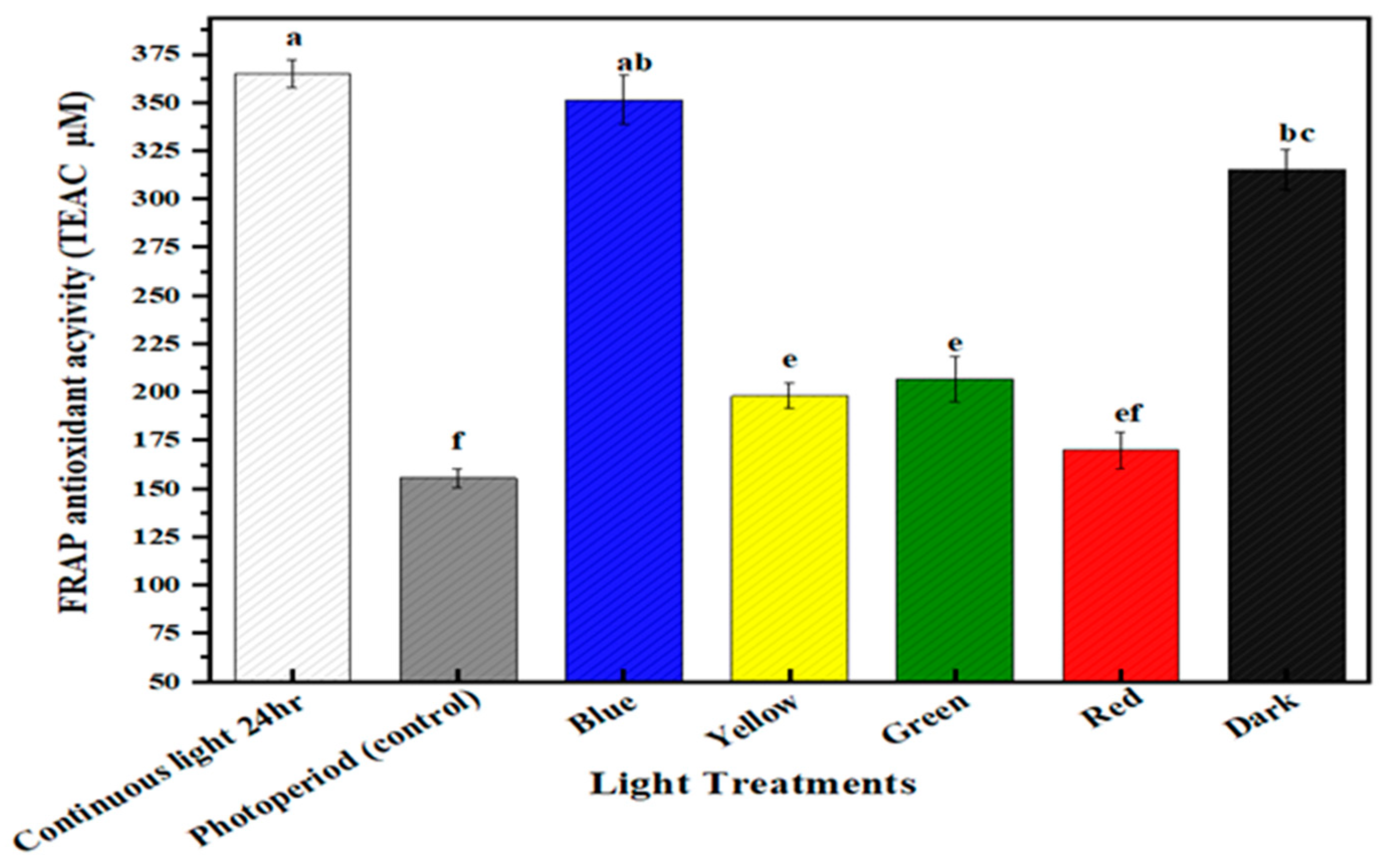
2.5. Multispectral Light Effects on the Antioxidant Potential of the Callus Culture of M. oleifera
3. Material and Methods
3.1. In Vitro Seed Germination
3.2. In Vitro Establishment of Callus Culture
3.3. Callus Exposure to Differential Lights
3.4. Harvesting and Extraction of Callus Culture Samples
3.5. Phytochemical Analysis
3.5.1. Phenolic and Flavonoid Contents
3.5.2. HPLC Quantification of Metabolites
3.6. Ferric-Reducing Antioxidant Power (FRAP) Assay
3.7. Antioxidant ABTS Assay
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Krasteva, G.; Georgiev, V.; Pavlov, A. Recent applications of plant cell culture technology in cosmetics and foods. Eng. Life Sci. 2021, 21, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Muyonga, J.; Nansereko, S.; Ilona Steenkamp, M.M.; Okoth, J. Linking agricultural universities with civil society, the private sector, governments and other stakeholders in support of agricultural development in Africa. In Potential Role of Traditional African Foods in Food Security, Nutrition and Health, Proceedings of the Fifth African Higher Education Week and Ruforum Biennial Conference 2016, Cape Town, South Africa, 17–21 October 2016; Ruforum: Kampala, Uganda, 2016; pp. 311–318. [Google Scholar]
- Khan, T.; Ullah, M.A.; Garros, L.; Hano, C.; Abbasi, B.H. Synergistic effects of melatonin and distinct spectral lights for enhanced production of anti-cancerous compounds in callus cultures of Fagonia indica. J. Photochem. Photobiol. B Biol. 2019, 190, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.T.; Mugal, T.; Haq, I.U. Moringa oleifera: A natural gift-A review. J. Pharm. Sci. Res. 2010, 2, 775. [Google Scholar]
- Lalas, S.; Tsaknis, J. Extraction and identification of natural antioxidant from the seeds of the Moringa oleifera tree variety of Malawi. J. Am. Oil Chem. Soc. 2002, 79, 677–683. [Google Scholar] [CrossRef]
- Maizuwo, A.I.; Hassan, A.S.; Momoh, H.; Muhammad, J.A. Phytochemical constituents, biological activities, therapeutic potentials and nutritional values of Moringa oleifera (Zogale): A review. J. Drug Des. Med. Chem. 2017, 3, 60. [Google Scholar]
- Dangi, S.; Jolly, C.I.; Narayanan, S. Antihypertensive activity of the total alkaloids from the leaves of Moringa oleifera. Pharm. Biol. 2002, 40, 144–148. [Google Scholar] [CrossRef]
- Panda, D.S.; Ansari, S.A. Preformulation study on the gum of Moringa oleifera. Malays. J. Pharm. Sci. 2013, 11, 41. [Google Scholar]
- Jaiswal, D.; Rai, P.K.; Kumar, A.; Mehta, S.; Watal, G. Effect of Moringa oleifera Lam. leaves aqueous extract therapy on hyperglycemic rats. J. Ethnopharmacol. 2009, 123, 392–396. [Google Scholar] [CrossRef]
- Santos, A.F.; Argolo, A.C.; Paiva, P.M.; Coelho, L.C. Antioxidant activity of Moringa oleifera tissue extracts. Phytother. Res. 2012, 26, 1366–1370. [Google Scholar] [CrossRef]
- Spandana, U.; Srikanth, P. A Review on Meracle tree: Moringa oleifera. J. Pharmacogn. Phytochem. 2016, 5, 189. [Google Scholar]
- Bhattacharya, A.; Tiwari, P.; Sahu, P.K.; Kumar, S. A review of the phytochemical and pharmacological characteristics of Moringa Oleifera. J. Pharm. Bioallied Sci. 2018, 10, 181. [Google Scholar]
- Cabardo, D.E., Jr.; Portugaliza, H.P. Anthelmintic activity of Moringa oleifera seed aqueous and ethanolic extracts against Haemonchus contortus eggs and third stage larvae. Int. J. Vet. Sci. Med. 2017, 5, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Mukherjee, P.K.; Saha, B. Studies on the antiulcer activity of Moringa oleifera leaf extract on gastric ulcer models in rats. Phytother. Res. 1995, 9, 463–465. [Google Scholar] [CrossRef]
- Sutar, N.G.; Bonde, C.; Patil, V.; Narkhede, S.; Patil, A.; Kakade, R. Analgesic activity of seeds of Moringa oleifera Lam. Int. J. Green Pharm. 2008, 2. [Google Scholar] [CrossRef]
- Peter, A.; Walter, A.; Wagai, S.; Joseph, O. Antibacterial activity of Moringa oleifera and Moringa stenopetala methanol and n-hexane seed extracts on bacteria implicated in water borne diseases. Afr. J. Microbiol. Res. 2011, 5, 153–157. [Google Scholar]
- Chollom, S.; Agada, G.; Gotep, J.; Mwankon, S.; Dus, P.; Bot, Y.; Nyango, D.; Singnap, C.; Fyaktu, E.; Okwori, A. Investigation of aqueous extract of Moringa oleifera lam seed for antiviral activity against newcastle disease virus in ovo. J. Med. Plants Res. 2012, 6, 3870–3875. [Google Scholar] [CrossRef]
- Padayachee, B.; Baijnath, H. An updated comprehensive review of the medicinal, phytochemical and pharmacological properties of Moringa oleifera. S. Afr. J. Bot. 2020, 129, 304–316. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Zhong, J.-J. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. Plant Cells 2001, 72, 1–26. [Google Scholar]
- Zaman, G.; Farooq, U.; Bajwa, M.N.; Jan, H.; Shah, M.; Ahmad, R.; Andleeb, A.; Drouet, S.; Hano, C.; Abbasi, B.H. Effects of yeast extract on the production of phenylpropanoid metabolites in callus culture of purple basil (Ocimum Basilicum L. var purpurascens) and their in-vitro evaluation for antioxidant potential. Plant Cell Tissue Organ Cult. 2022, 150, 543–553. [Google Scholar] [CrossRef]
- Ahmed, K.; Tariq, I.; Mudassir, S.U.S.M. 11. Green synthesis of cobalt nanoparticles by using methanol extract of plant leaf as reducing agent. Pure Appl. Biol. 2021, 5, 453–457. [Google Scholar]
- Bajwa, M.N.; Bibi, A.; Idrees, M.Z.; Zaman, G.; Farooq, U.; Bhatti, T.T. Elicitation, a mechanistic approach to change the metabolic pathway of plants to produce pharmacological important compounds in in-vitro cell cultures. Glob. J. Eng. 2021, 8, 1–7. [Google Scholar]
- Ullah, M.A.; Gul, F.Z.; Khan, T.; Bajwa, M.N.; Drouet, S.; Tungmunnithum, D.; Giglioli-Guivarc’h, N.; Liu, C.; Hano, C.; Abbasi, B.H. Differential induction of antioxidant and anti-inflammatory phytochemicals in agitated micro-shoot cultures of Ajuga integrifolia Buch. Ham. ex D. Don with biotic elicitors. AMB Express 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Nazir, M.; Tungmunnithum, D.; Bose, S.; Drouet, S.; Garros, L.; Giglioli-Guivarc’h, N.; Abbasi, B.H.; Hano, C. Differential production of phenylpropanoid metabolites in callus cultures of Ocimum basilicum L. with distinct in vitro antioxidant activities and in vivo protective effects against UV stress. J. Agric. Food Chem. 2019, 67, 1847–1859. [Google Scholar] [CrossRef] [PubMed]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Shama, G.; Alderson, P. UV hormesis in fruits: A concept ripe for commercialisation. Trends Food Sci. Technol. 2005, 16, 128–136. [Google Scholar] [CrossRef]
- Turtoi, M. Ultraviolet light treatment of fresh fruits and vegetables surface: A review. J. Agroaliment. Process. Technol. 2013, 19, 325–337. [Google Scholar]
- Anjum, S.; Abbasi, B.H.; Doussot, J.; Favre-Réguillon, A.; Hano, C. Effects of photoperiod regimes and ultraviolet-C radiations on biosynthesis of industrially important lignans and neolignans in cell cultures of Linum usitatissimum L.(Flax). J. Photochem. Photobiol. B Biol. 2017, 167, 216–227. [Google Scholar] [CrossRef]
- Yang, Q.; Pan, J.; Shen, G.; Guo, B. Yellow light promotes the growth and accumulation of bioactive flavonoids in Epimedium pseudowushanense. J. Photochem. Photobiol. B Biol. 2019, 197, 11155057. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H. Light-induced fluctuations in biomass accumulation, secondary metabolites production and antioxidant activity in cell suspension cultures of Artemisia absinthium L. J. Photochem. Photobiol. B Biol. 2014, 140, 223–227. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, S.S.; Akbar, F.; Kanwal, F. Correlation of different spectral lights with biomass accumulation and production of antioxidant secondary metabolites in callus cultures of medicinally important Prunella vulgaris L. J. Photochem. Photobiol. B Biol. 2016, 159, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Abbasi, B.H.; ul Haq, I. Red light controlled callus morphogenetic patterns and secondary metabolites production in Withania somnifera L. Biotechnol. Rep. 2019, 24, e00380. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Hano, C.; Abbasi, B.H. Monochromatic lights-induced trends in antioxidant and antidiabetic polyphenol accumulation in in vitro callus cultures of Lepidium sativum L. J. Photochem. Photobiol. B Biol. 2019, 196, 111505. [Google Scholar] [CrossRef] [PubMed]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Ali, M.B.; Singh, N.; Shohael, A.M.; Hahn, E.J.; Paek, K.-Y. Phenolics metabolism and lignin synthesis in root suspension cultures of Panax ginseng in response to copper stress. Plant Sci. 2006, 171, 147–154. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Tian, C.-L.; Murch, S.J.; Saxena, P.K.; Liu, C.-Z. Light-enhanced caffeic acid derivatives biosynthesis in hairy root cultures of Echinacea purpurea. Plant Cell Rep. 2007, 26, 1367–1372. [Google Scholar] [CrossRef]
- Wu, J.; Zhong, J.-J. Production of ginseng and its bioactive components in plant cell culture: Current technological and applied aspects. J. Biotechnol. 1999, 68, 89–99. [Google Scholar] [CrossRef]
- Sarkar, J.; Ghosh, P.; Adil, A. A review on hybrid nanofluids: Recent research, development and applications. Renew. Sustain. Energy Rev. 2015, 43, 164–177. [Google Scholar] [CrossRef]
- Younas, M.; Drouet, S.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Differential accumulation of silymarin induced by exposure of Silybum marianum L. callus cultures to several spectres of monochromatic lights. J. Photochem. Photobiol. B Biol. 2018, 184, 61–70. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H.; Szewczyk, A.; Fugas, E. Production of bioactive phenolic acids and furanocoumarins in in vitro cultures of Ruta graveolens L. and Ruta graveolens ssp. divaricata (Tenore) Gams. under different light conditions. Plant Cell Tissue Organ Cult. 2012, 110, 329–336. [Google Scholar]
- Ekiert, H.; Kubica, P.; Kwiecień, I.; Jafernik, K.; Klimek-Szczykutowicz, M.; Szopa, A. Cultures of Medicinal Plants In Vitro as a Potential Rich Source of Antioxidants. Plant Antioxid. Health 2020, 1–44. [Google Scholar] [CrossRef]
- Szopa, A.; Starzec, A.; Ekiert, H. The importance of monochromatic lights in the production of phenolic acids and flavonoids in shoot cultures of Aronia melanocarpa, Aronia arbutifolia and Aronia × prunifolia. J. Photochem. Photobiol. B Biol. 2018, 179, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Barnaś, M.; Ekiert, H. Phytochemical studies and biological activity of three Chinese Schisandra species (Schisandra sphenanthera, Schisandra henryi and Schisandra rubriflora): Current findings and future applications. Phytochem. Rev. 2019, 18, 109–128. [Google Scholar] [CrossRef]
- Khan, T.; Abbasi, B.H.; Khan, M.A. The interplay between light, plant growth regulators and elicitors on growth and secondary metabolism in cell cultures of Fagonia indica. J. Photochem. Photobiol. B Biol. 2018, 185, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.C.-Y.; Chen, C.-H.; Chen, S.-H.; Lu, I.; Chu, M.-J.; Huang, L.-C.; Lin, C.-Y.; Chen, C.-Y.; Lo, H.-F.; Jeng, S.-T. The effect of red light and far-red light conditions on secondary metabolism in agarwood. BMC Plant Biol. 2015, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Dajas, F. Life or death: Neuroprotective and anticancer effects of quercetin. J. Ethnopharmacol. 2012, 143, 383–396. [Google Scholar] [CrossRef]
- Nadeem, M.; Abbasi, B.H.; Younas, M.; Ahmad, W.; Zahir, A.; Hano, C. LED-enhanced biosynthesis of biologically active ingredients in callus cultures of Ocimum basilicum. J. Photochem. Photobiol. B Biol. 2019, 190, 172–178. [Google Scholar] [CrossRef]
- Middleton, E.M.; Teramura, A.H. The role of flavonol glycosides and carotenoids in protecting soybean from ultraviolet-B damage. Plant Physiol. 1993, 103, 741–752. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Moran, J.F.; Becana, M.; Iturbe-Ormaetxe, I.; Frechilla, S.; Klucas, R.V.; Aparicio-Tejo, P. Drought induces oxidative stress in pea plants. Planta 1994, 194, 346–352. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Ashry, N.A.; Mohamed, H.I. Impact of secondary metabolites and related enzymes in flax resistance and or susceptibility to powdery mildew. World J. Agric. Sci. 2011, 7, 78–85. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Urbonavičiūtė, A.; Šabajevienė, G.; Duchovskis, P. The effect of red and blue light component on the growth and development of frigo strawberries. Zemdirb.-Agric. 2010, 97, 99–104. [Google Scholar]
- Beckwith, A.G.; Zhang, Y.; Seeram, N.P.; Cameron, A.C.; Nair, M.G. Relationship of light quantity and anthocyanin production in Pennisetum setaceum cvs. Rubrum and Red Riding Hood. J. Agric. Food Chem. 2004, 52, 456–461. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Kim, D.-O.; Chun, O.K.; Kim, Y.J.; Moon, H.-Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef] [PubMed]
- Zahir, A.; Ahmad, W.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. In vitro cultures of Linum usitatissimum L.: Synergistic effects of mineral nutrients and photoperiod regimes on growth and biosynthesis of lignans and neolignans. J. Photochem. Photobiol. B Biol. 2018, 187, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, R.; Ullah, M.A.; Tungmunnithum, D.; Drouet, S.; Shah, M.; Zaeem, A.; Hameed, S.; Hano, C.; Abbasi, B.H. Lights triggered differential accumulation of antioxidant and antidiabetic secondary metabolites in callus culture of Eclipta alba L. PLoS ONE 2020, 15, e0233963. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker Compounds (µg/g DW) | Different Lights Treatment | ||||||
|---|---|---|---|---|---|---|---|
| Control | Red | Green | Blue | Dark | White Light | Yellow | |
| Apigenin | 594.65 ± 0.009 | 558.09 ± 0.03 | 574.63 ± 0.21 | 611.50 ± 0.51 | 627.60 ± 0.07 | 751.19 ± 0.44 | 459.56 ± 0.011 |
| Luteolin | 751.19 ± 0.004 | 592.29 ± 0.05 | 715.88 ± 0.33 | 540.40 ± 0.28 | 0.169 ± 0.016 | 798.29 ± 0.75 | 433.39 ± 0.077 |
| p-coumaric acid | 89.07 ± 0.006 | 79.08 ± 0.07 | 148.41 ± 0.05 | 105.43 ± 0.047 | 137.09 ± 0.63 | 159.03 ± 0.54 | 78.07 ± 0.086 |
| Neochlorogenic acid | 789.65 ± 0.064 | 611.16 ± 0.02 | 635.08 ± 0.19 | 826.75 ± 0.044 | 754.19 ± 0.65 | 998.38 ± 0.25 | 662.91 ± 0.006 |
| Chlorogenic acid | 144.92 ± 0.855 | 128.63 ± 0.82 | 86.93 ± 1.209 | 188.14 ± 1.55 | 138.07 ± 1.66 | 210.57 ± 0.99 | 144.92 ± 0.66 |
| Quercetin | 592.29 ± 1.776 | 701.07 ± 2.05 | 665.56 ± 1.99 | 721.45 ± 2.44 | 821.82 ± 3.055 | 959.92 ± 1.87 | 433.39 ± 1.363 |
| Kaempferol | 468.70 ± 1.055 | 537.87 ± 0.79 | 565.89 ± 0.622 | 721.65 ± 0.83 | 844.51 ± 1.22 | 1016.04 ± 1.22 | 405.97 ± 0.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajwa, M.N.; Khanum, M.; Zaman, G.; Ullah, M.A.; Farooq, U.; Waqas, M.; Ahmad, N.; Hano, C.; Abbasi, B.H. Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera. Molecules 2023, 28, 1497. https://doi.org/10.3390/molecules28031497
Bajwa MN, Khanum M, Zaman G, Ullah MA, Farooq U, Waqas M, Ahmad N, Hano C, Abbasi BH. Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera. Molecules. 2023; 28(3):1497. https://doi.org/10.3390/molecules28031497
Chicago/Turabian StyleBajwa, Muhammad Naeem, Mehnaz Khanum, Gouhar Zaman, Muhammad Asad Ullah, Umar Farooq, Muhammad Waqas, Nisar Ahmad, Christophe Hano, and Bilal Haider Abbasi. 2023. "Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera" Molecules 28, no. 3: 1497. https://doi.org/10.3390/molecules28031497
APA StyleBajwa, M. N., Khanum, M., Zaman, G., Ullah, M. A., Farooq, U., Waqas, M., Ahmad, N., Hano, C., & Abbasi, B. H. (2023). Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera. Molecules, 28(3), 1497. https://doi.org/10.3390/molecules28031497