

Study on the Cellular Anti-Inflammatory Effect of Torularhodin Produced by Sporidiobolus pararoseus ZQHL Isolated from Vinegar Fungus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Origin and Distribution of Sporidiobolus pararoseus
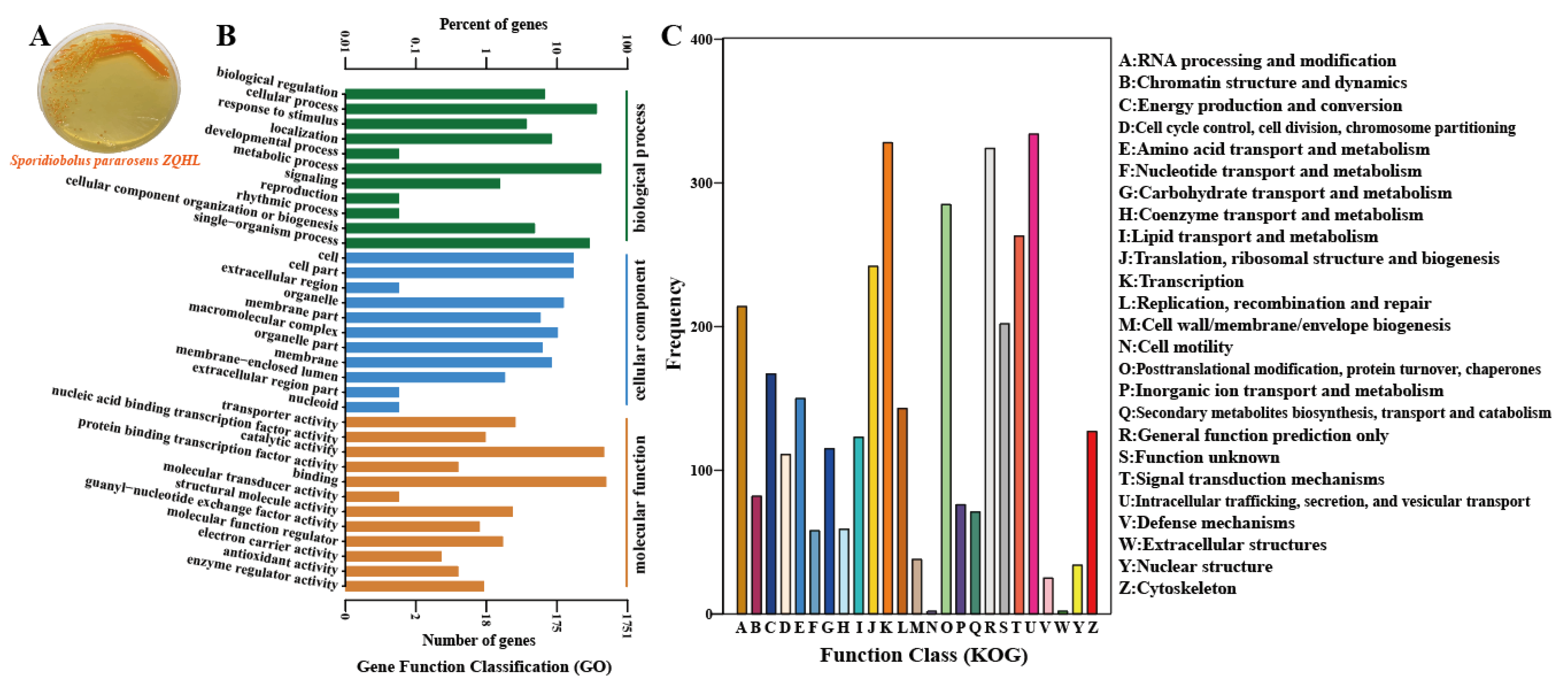
2.2. Genome-Wide and Functional Annotation of Sporidiobolus pararoseus ZQHL
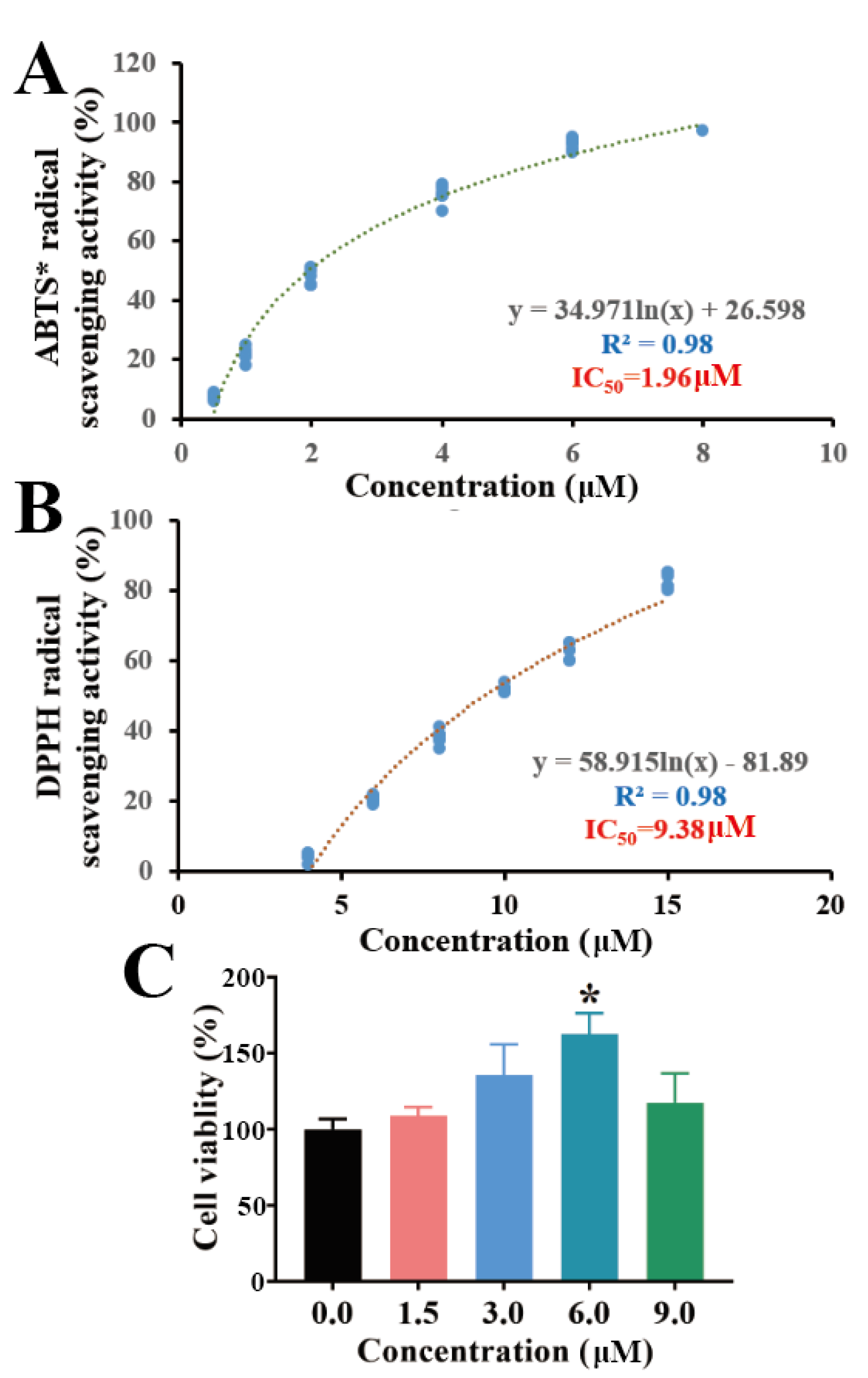
2.3. Antioxidant Capacity and Cytotoxicity of Torularhodin in Sporidiobolus pararoseus ZQHL
2.4. Cellular Anti-Inflammatory Activity of Torularhodin in Sporidiobolus pararoseus ZQHL
3. Materials and Methods
3.1. Materials and Reagents
3.2. Sample Collection and ITS Sequencing
3.3. Isolation, Purification, and Cultivation of Fungi
3.4. Whole Genome Sequencing and Assembly
3.5. Gene Function Annotation Analysis
3.6. Isolation and Purification of Torularhodin
3.7. Free Radical Scavenging Ability of Torularhodin In Vitro
3.8. Effect of Torularhodin on LPS-Induced Inflammation in RAW 264.7 Cells
3.9. Real-Time Quantitative PCR Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Mussagy, C.U.; Ribeiro, H.F.; Santos-Ebinuma, V.C.; Schuur, B.; Pereira, J.F.B. Rhodotorula sp.-based biorefinery: A source of valuable biomolecules. Appl. Microbiol. Biotechnol. 2022, 106, 7431–7447. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar]
- Fazili, A.B.A.; Shah, A.M.; Zan, X.; Naz, T.; Nosheen, S.; Nazir, Y.; Ullah, S.; Zhang, H.; Song, Y. Mucor circinelloides: A model organism for oleaginous fungi and its potential applications in bioactive lipid production. Microb. Cell Factories 2022, 21, 29. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids and Their Biosynthesis in Fungi. Molecules 2022, 27, 1431. [Google Scholar] [CrossRef]
- Zhang, Q.W.; Kong, C.L.; Tao, Y.S. Fate of carotenoids in yeasts: Synthesis and cleavage. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef]
- Kanamoto, H.; Nakamura, K.; Misawa, N. Carotenoid Production in Oleaginous Yeasts. Adv. Exp. Med. Biol. 2021, 1261, 153–163. [Google Scholar]
- Wang, H.; Hu, B.; Liu, J.; Qian, H.; Xu, J.; Zhang, W. Co-production of lipid, exopolysaccharide and single-cell protein by Sporidiobolus pararoseus under ammonia nitrogen-limited conditions. Bioprocess Biosyst. Eng. 2020, 43, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Innocenti, M.; Turchetti, B.; Libkind, D.; van Broock, M.; Mulinacci, N. Carotenoid profiles of yeasts belonging to the genera Rhodotorula, Rhodosporidium, Sporobolomyces, and Sporidiobolus. Can. J. Microbiol. 2007, 53, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Lv, T.; Liu, Q.; Cheng, Y.; Liu, C.; Han, M.; Zhang, W.; Qian, H. Carotenoids in Sporidiobolus pararoseus ameliorate diabetic nephropathy in mice through attenuating oxidative stress. Biol. Chem. 2021, 402, 785–794. [Google Scholar] [CrossRef]
- Li, X.; Cheng, Y.; Li, J.; Liu, C.; Qian, H.; Zhang, G. Torularhodin Alleviates Hepatic Dyslipidemia and Inflammations in High-Fat Diet-Induced Obese Mice via PPARalpha Signaling Pathway. Molecules 2022, 27, 6398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Hua, H.; Guo, Y.; Cheng, Y.; Pi, F.; Yao, W.; Xie, Y.; Qian, H. Torularhodin from Sporidiobolus pararoseus Attenuates d-galactose/AlCl(3)-Induced Cognitive Impairment, Oxidative Stress, and Neuroinflammation via the Nrf2/NF-kappaB Pathway. J. Agric. Food Chem. 2020, 68, 6604–6614. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Pi, F.; Guo, Y.; Cheng, Y.; Qian, H. Torularhodin Ameliorates Oxidative Activity in Vitro and d-Galactose-Induced Liver Injury via the Nrf2/HO-1 Signaling Pathway in Vivo. J. Agric. Food Chem. 2019, 67, 10059–10068. [Google Scholar] [CrossRef]
- Mussagy, C.U.; Gonzalez-Miquel, M.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Microbial torularhodin—A comprehensive review. Crit. Rev. Biotechnol. 2022, 1–19. [Google Scholar] [CrossRef]
- Yuan, R.; Huang, L.; Du, L.J.; Feng, J.F.; Li, J.; Luo, Y.Y.; Xu, Q.M.; Yang, S.L.; Gao, H.; Feng, Y.L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking TLR4 dimerization. Pharmacol. Res. 2019, 142, 102–114. [Google Scholar] [CrossRef]
- Troy, N.M.; Strickland, D.; Serralha, M.; de Jong, E.; Jones, A.C.; Read, J.; Galbraith, S.; Islam, Z.; Kaur, P.; Mincham, K.T.; et al. Protection against severe infant lower respiratory tract infections by immune training: Mechanistic studies. J. Allergy Clin. Immunol. 2022, 150, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.J. Antioxidant Potential and Capacity of Microorganism-Sourced C(30) Carotenoids-A Review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, V.; Flora, G.; Sevanan, M.; Sripriya, R.; Chen, W.H.; Park, J.H.; Rajesh Banu, J.; Kumar, G. Technological advances in the production of carotenoids and their applications—A critical review. Bioresour. Technol. 2023, 367, 128215. [Google Scholar] [CrossRef]
- Szotkowski, M.; Holub, J.; Simansky, S.; Hubacova, K.; Hladka, D.; Nemcova, A.; Marova, I. Production of Enriched Sporidiobolus sp. Yeast Biomass Cultivated on Mixed Coffee Hydrolyzate and Fat/Oil Waste Materials. Microorganisms 2021, 9, 1848. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, F.; Liu, T.; Zhao, C.; Gu, F.; Du, H.; Wang, F.; Zheng, J.; Xiao, H. Carotenoid fates in plant foods: Chemical changes from farm to table and nutrition. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Beydoun, M.A.; Chen, X.; Jha, K.; Beydoun, H.A.; Zonderman, A.B.; Canas, J.A. Carotenoids, vitamin A, and their association with the metabolic syndrome: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 32–45. [Google Scholar] [CrossRef]
- Vitucci, D.; Amoresano, A.; Nunziato, M.; Muoio, S.; Alfieri, A.; Oriani, G.; Scalfi, L.; Frusciante, L.; Rigano, M.M.; Pucci, P.; et al. Nutritional Controlled Preparation and Administration of Different Tomato Purees Indicate Increase of beta-Carotene and Lycopene Isoforms, and of Antioxidant Potential in Human Blood Bioavailability: A Pilot Study. Nutrients 2021, 13, 1336. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Zhuang, C.; Yuan, J.; Du, Y.; Zeng, J.; Sun, Y.; Wu, Y.; Gao, X.H.; Chen, H.D. Effects of Oral Carotenoids on Oxidative Stress: A Systematic Review and Meta-Analysis of Studies in the Recent 20 Years. Front. Nutr. 2022, 9, 754707. [Google Scholar] [CrossRef]
- Wolf, G. Retinoids and carotenoids as inhibitors of carcinogenesis and inducers of cell-cell communication. Nutr. Rev. 1992, 50, 270–274. [Google Scholar] [CrossRef]
- Richard, K.; Vogel, S.N.; Perkins, D.J. Quantitation of TLR4 Internalization in Response to LPS in Thioglycollate Elicited Peritoneal mouse Macrophages by Flow Cytometry. Bio-Protocol 2019, 9, e3369. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Oh, S.; Kum, E.; Seo, S.; Park, Y.; Kim, G. Immunomodulatory Effects of an Aqueous Extract of Black Radish on Mouse Macrophages via the TLR2/4-Mediated Signaling Pathway. Pharmaceuticals 2022, 15, 1376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Munch, G.; Wohlmuth, H.; Afzal, S.; Kao, M.T.; Al-Khazaleh, A.; Low, M.; Leach, D.; Li, C.G. Synergistic Inhibition of Pro-Inflammatory Pathways by Ginger and Turmeric Extracts in RAW 264.7 Cells. Front. Pharmacol. 2022, 13, 818166. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Sun, C.; Liu, X.; Wang, X.; Li, W.; Wei, X.; Li, Q.; Ju, J. Upregulation of a marine fungal biosynthetic gene cluster by an endobacterial symbiont. Commun. Biol. 2020, 3, 527. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheng, Y.; Du, C.; Lv, T.; Guo, Y.; Han, M.; Pi, F.; Zhang, W.; Qian, H. Study on the wall-breaking method of carotenoids producing yeast Sporidiobolus pararoseus and the antioxidant effect of four carotenoids on SK-HEP-1 cells. Prep. Biochem. Biotechnol. 2019, 49, 767–774. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, H.; Du, C.; Zhang, W.; Qian, H. Tentative identification of torulene cis/trans geometrical isomers isolated from Sporidiobolus pararoseus by high-performance liquid chromatography-diode array detection-mass spectrometry and preparation by column chromatography. Anal. Sci. 2013, 29, 997–1002. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Y.; Cheng, Y.; Qian, H. Bilosomes: A controlled delivery system for the sustained release of torularhodin during digestion in the small intestine both in vitro and in vivo. Colloids Surf. A Physicochem. Eng. Asp. 2022, 654, 5. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Y.; Chen, S.; Lin, J.; Yang, X.; Huang, D. Structure-Activity Relationship (SAR) of Flavones on Their Anti-Inflammatory Activity in Murine Macrophages in Culture through the NF-kappaB Pathway and c-Src Kinase Receptor. J. Agric. Food Chem. 2022, 70, 8788–8798. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Han, M.; Lv, F.; Gao, Y.; Wang, X.; Zhang, X.; Guo, Y.; Cheng, Y.; Qian, H. Study on the Cellular Anti-Inflammatory Effect of Torularhodin Produced by Sporidiobolus pararoseus ZQHL Isolated from Vinegar Fungus. Molecules 2023, 28, 1436. https://doi.org/10.3390/molecules28031436
Liu C, Han M, Lv F, Gao Y, Wang X, Zhang X, Guo Y, Cheng Y, Qian H. Study on the Cellular Anti-Inflammatory Effect of Torularhodin Produced by Sporidiobolus pararoseus ZQHL Isolated from Vinegar Fungus. Molecules. 2023; 28(3):1436. https://doi.org/10.3390/molecules28031436
Chicago/Turabian StyleLiu, Chang, Mei Han, Fuqiang Lv, Yaobin Gao, Xiaoyun Wang, Xujiao Zhang, Yahui Guo, Yuliang Cheng, and He Qian. 2023. "Study on the Cellular Anti-Inflammatory Effect of Torularhodin Produced by Sporidiobolus pararoseus ZQHL Isolated from Vinegar Fungus" Molecules 28, no. 3: 1436. https://doi.org/10.3390/molecules28031436
APA StyleLiu, C., Han, M., Lv, F., Gao, Y., Wang, X., Zhang, X., Guo, Y., Cheng, Y., & Qian, H. (2023). Study on the Cellular Anti-Inflammatory Effect of Torularhodin Produced by Sporidiobolus pararoseus ZQHL Isolated from Vinegar Fungus. Molecules, 28(3), 1436. https://doi.org/10.3390/molecules28031436