
Selenium Biofortification Effect on Glucosinolate Content of Brassica oleracea var. italic and Eruca vesicaria




Abstract
:1. Introduction
2. Results and Discussion
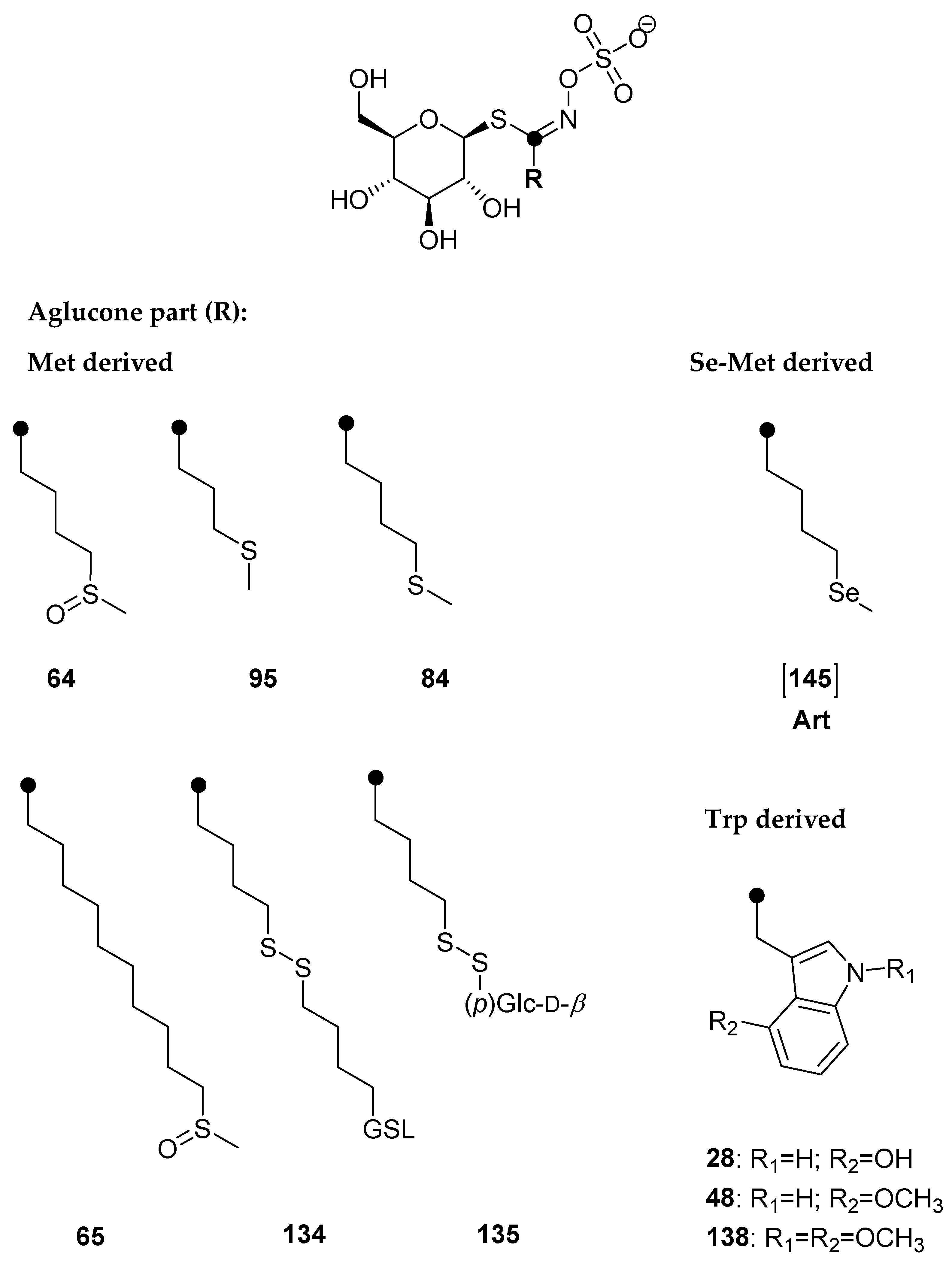
2.1. Effect of Selenium Treatment on Broccoli
2.2. Effect of Selenium Treatment on Rocket
3. Materials and Methods
3.1. Materials and Reagents
3.2. Plant Growth and Harvesting
3.3. Isolation and Chemical Analysis
3.3.1. Isolation of Desulfoglucosinolates
3.3.2. UHPLC-MS/MS Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2007. [Google Scholar] [CrossRef]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Revised Edition; Selinus, O., Ed.; British Geological Survey: Nottingham, UK, 2013; pp. 375–419. [Google Scholar] [CrossRef]
- dos Reis, A.R.; El-Ramady, H.; Santos, E.F.; Gratão, P.L.; Schomburg, L. Overview of Selenium Deficiency and Toxicity Worldwide: Affected Areas, Selenium-Related Health Issues, and Case Studies. In Selenium in Plants; Plant Ecophysiology; Pilon-Smits, E., Winkel, L., Lin, Z.Q., Eds.; Springer: Cham, Switzerland, 2017; Volume 11. [Google Scholar] [CrossRef]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Ajwa, H.A.; Banuelos, G.S.; Mayland, H.F. Selenium uptake by plants from soils amended with inorganic and organic materials. J. Environ. Qual. 1998, 27, 1218–1227. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in Brassicaceous vegetables, and their application to human health. Front. Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel sources and biological potential. In Glucosinolates, Reference Series in Phytochemistry; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 3–60. [Google Scholar] [CrossRef]
- Montaut, S.; De Nicola, G.R.; Agnaniet, H.; Issembe, Y.; Rollin, P.; Menut, C. Probing for the presence of glucosinolates in three Drypetes spp. (Drypetes euryodes (Hiern) Hutch., Drypetes gossweileri S. Moore, Drypetes laciniata Hutch.) and two Rinorea spp. (Rinorea subintegrifolia O. Ktze and Rinorea woermanniana (Büttner) Engl.) from Gabon. Nat. Prod. Res. 2017, 31, 308–313. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Montaut, S.; Read, S.; Blažević, I.; Nuzillard, J.-M.; Roje, M.; Harakat, D.; Rollin, P. Investigation of the glucosinolates in Hesperis matronalis L. and Hesperis laciniata All.: Unveiling 4′-O-β-D-apiofuranosylglucomatronalin. Carbohydr. Res. 2020, 488, 107898. [Google Scholar] [CrossRef] [PubMed]
- Trabelcy, B.; Chinkov, N.; Samuni-Blank, M.; Merav, M.; Izhaki, I.; Carmeli, S.; Gerchman, Y. Investigation of glucosinolates in the desert plant Ochradenus baccatus (Brassicales: Resedaceae). Unveiling glucoochradenin, a new arabinosylated glucosinolate. Phytochemistry 2021, 187, 112760. [Google Scholar] [CrossRef]
- Stewart, J.M.; Nigiam, S.N.; McConnel, W.B. Metabolism of Na275SeO4 in horseradish: Formation of selenosinigrin. Can. J. Biochem. 1974, 52, 144–145. [Google Scholar] [CrossRef]
- Bertelsen, F.; Gissel-Nielsen, G.; Kjær, A.; Skrydstrup, T. Selenoglucosinolates in nature: Fact or myth? Phytochemistry 1988, 27, 3743–3749. [Google Scholar] [CrossRef]
- Matich, A.J.; McKenzie, M.J.; Lill, R.E.; Brummell, D.A.; McGhie, T.K.; Chen, R.K.-Y.; Rowan, D.D. Selenoglucosinolates and their metabolites produced in Brassica spp. fertilised with sodium selenate. Phytochemistry 2012, 75, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Matich, A.J.; McKenzie, M.J.; Lill, R.E.; McGhie, T.K.; Chen, R.K.; Rowan, D.D. Distribution of selenoglucosinolates and their metabolites in Brassica treated with sodium selenate. J. Agric. Food Chem. 2015, 63, 1896–1905. [Google Scholar] [CrossRef]
- Ouerdane, L.; Aureli, F.; Flis, P.; Bierla, K.; Preud’homme, H.; Cubadda, F.; Szpunar, J. Comprehensive speciation of low-molecular weight selenium metabolites in mustard seeds using HPLC-electrospray linear trap/Orbitrap tandem mass spectrometry. Metallomics 2013, 5, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, M.; Matich, A.; Hunter, D.; Esfandiari, A.; Trolove, S.; Chen, R.; Lill, R. Selenium application during radish (Raphanus sativus) plant development alters glucosinolate metabolic gene expression and results in the production of 4-(methylseleno)but-3-enyl glucosinolate. Plants 2019, 8, 427. [Google Scholar] [CrossRef] [PubMed]
- European Economic Community, Commisssion Regulation. Oilseeds—determination of glucosinolates high performance liquid chromatography. Off. J. Eur. Comm. 1990, L170, 27–34. [Google Scholar]
- Van Hoewyk, D.; Takahashi, H.; Inoue, E.; Hess, A.; Tamaoki, M.; Pilon-Smits, E.A.H. Transcriptome analyses give insights into selenium-stress responses and selenium tolerance mechanisms in Arabidopsis. Physiol. Plant. 2008, 132, 236–253. [Google Scholar] [CrossRef]
- Lehotai, N.; Feigl, G.; Koos, A.; Molnar, A.; Ördög, A.; Peto, A.; Erdei, L.; Kolbert, Z.S. Nitric oxide-cytokinin interplay influences selenite sensitivity in Arabidopsis. Plant Cell Rep. 2016, 35, 2181–2195. [Google Scholar] [CrossRef]
- Kolbert, Z.; Lehotai, N.; Molnár, Á.; Feigl, G. “The roots” of selenium toxicity: A new concept. Plant Signal. Behav. 2016, 11, e1241935. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulou, M.; Bouzas, E.A.; Siyiannis, V.; Perouli, M.; Kokotou, M.; Chorianopoulou, S.N.; Constantinou-Kokotou, V.; Bouranis, D.L. Selenium assimilation by broccoli: Effect of Se inputs on the biosynthesis of secondary metabolites under normal or reduced S inputs. In Proceedings of the 28th International Symposium of CIEC, Fertilization and Nutrient Use Efficiency in Mediterranean Environments; Bouranis, D.L., Haneklaus, S.H., Chorianopoulou, S.N., Li, J., De Kok, L.J., Schnug, E., Ji, L., Eds.; Utopia Publishing Ltd.: Athens, Greece, 2020; pp. 169–173. [Google Scholar]
- Bouranis, D.L.; Stylianidis, G.P.; Manta, V.; Karousis, E.N.; Tzanaki, A.; Dimitriadi, D.; Bouzas, E.A.; Siyiannis, V.F.; Constantinou-Kokotou, V.; Chorianopoulou, S.N.; et al. Floret biofortification of broccoli using amino acids coupled with selenium under different surfactants: A case study of cultivating functional foods. Plants 2023, 12, 1272. [Google Scholar] [CrossRef]
- Tian, M.; Hui, M.; Thannhauser, T.W.; Pan, S.; Li, L. Selenium-induced toxicity is counteracted by sulfur in broccoli (Brassica oleracea L. var. italica). Front. Plant Sci. 2017, 8, 1425. [Google Scholar] [CrossRef] [PubMed]
- Olsen, C.E.; Huang, X.C.; Hansen, C.I.C.; Cipollini, D.; Ørgaard, M.; Matthes, A.; Geu-Flores, F.; Koch, M.A.; Agerbirk, N. Glucosinolate diversity within a phylogenetic framework of the tribe Cardamineae (Brassicaceae) unraveled with HPLC-MS/MS and NMR-based analytical distinction of 70 desulfoglucosinolates. Phytochemistry 2016, 132, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Pfalz, M.; Mukhaimar, M.; Perreau, F.; Kirk, J.; Hansen, C.I.C.; Olsen, C.E.; Agerbirk, N.; Kroymann, J. Methyl transfer in glucosinolate biosynthesis mediated by indole glucosinolate O-Methyltransferase 5. Plant Physiol. 2016, 172, 2190–2203. [Google Scholar] [CrossRef]
- Agerbirk, N.; Hansen, C.C.; Olsen, C.E.; Kiefer, C.; Hauser, T.P.; Christensen, S.; Jensen, K.R.; Ørgaard, M.; Pattison, D.I.; Lange, C.B.A.; et al. Glucosinolate profiles and phylogeny in Barbarea compared to other tribe Cardamineae (Brassicaceae) and Reseda (Resedaceae), based on a library of ion trap HPLC-MS/MS data of reference desulfoglucosinolates. Phytochemistry 2021, 185, 112658. [Google Scholar] [CrossRef] [PubMed]
- Dall’Acqua, S.; Ertani, A.; Pilon-Smits, E.A.H.; Fabrega-Prats, M.; Schiavon, M. Selenium biofortification differentially affects sulfur metabolism and accumulation of phytochemicals in two rocket species (Eruca sativa Mill. and Diplotaxis tenuifolia) grown in hydroponics. Plants 2019, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K.; Wu, Q. Effects of sulfur and selenium on glucosinolate biosynthesis in cabbage. Plant Mol. Biol. Rep. 2020, 38, 62–74. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-assisted versus conventional isolation of glucosinolate degradation products from Lunaria annua L. and their cytotoxic activity. Biomolecules 2020, 10, 215. [Google Scholar] [CrossRef]
- Wathelet, J.-P.; Iori, R.; Leoni, O.; Quinsac, A.; Palmieri, S.; Rollin, P. Guidelines for glucosinolate analysis in green tissues used for biofumigation. Agroindustria 2004, 3, 257–344. [Google Scholar]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Glucosinolate | Formula (desulfo) | Desulfo M | Desulfo [M + Na]+ | Reference | Na2SeO4 Solution | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 mM | 5 mM | ||||||||||||
| Root | Stem | Leaf | Root | Stem | Leaf | Root | Stem | Leaf | |||||
| Met derived | |||||||||||||
| 73 | 3-(Methylsulfinyl)propyl GSL (glucoiberin) a | C11H21NO7S2 | 343 | 366 | tr | tr | n.d. | tr | tr | n.d. | tr | tr | n.d. |
| 64 | 4-(Methylsulfinyl)butyl GSL (glucoraphanin) a | C12H23NO7S2 | 357 | 380 | 2.09 ± 0.12 A | 1.11 ± 0.06 D | tr | 1.40 ± 0.13 E | 0.58 ± 0.04 B | tr | 3.68 ± 0.87 C | 0.44 ± 0.12 E | tr |
| 95 | 3-(Methylsulfanyl)propyl GSL (glucoibervirin) b | C11H21NO6S2 | 327 | 350 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.03 ± 0.12 | n.d. | n.d. |
| 84 | 4-(Methylsulfanyl)butyl GSL (glucoerucin) a | C12H23NO6S2 | 341 | 364 | 4.88 ± 0.72 A | 0.13 ± 0.00 D | n.d. | 9.05 ± 0.12 B | 0.11 ± 0.00 E | n.d. | 9.89 ± 0.57 B | tr | n.d. |
| 94 | 5-(Methylsulfanyl)pentyl GSL (glucoberteroin) a | C15H21NO6S | 355 | 378 | tr | n.d. | tr | n.d. | n.d. | n.d. | tr | n.d. | n.d. |
| Phe derived | |||||||||||||
| 105 | 2-Phenylethyl GSL (gluconasturtiin) a | C11H21NO7S2 | 343 | 366 | tr | n.d. | tr | tr | n.d. | n.d. | 0.31 ± 0.00 | n.d. | n.d. |
| Trp derived | |||||||||||||
| 43 | Indol-3-ylmethyl GSL (glucobrassicin) a | C16H20N2O6S | 368 | 391 | 0.68 ± 0.14 A | 0.07 ± 0.00 D | 0.31 ± 0.04 G | 2.34 ± 0.13 B | 0.55 ± 0.12 E | 0.09 ± 0.00 H | 1.20 ± 0.18 C | 0.25 ± 0.00 F | 0.58 ± 0.12 I |
| 28 | 4-Hydroxyindol-3-ylmethyl GSL (4-hydroxyglucobrassicin) a | C16H20N2O7S | 384 | 407 | 0.62 ± 0.10 A | tr | 0.60 ± 0.20 G | 0.78 ± 0.19 A | 0.22 ± 0.09 | 0.51 ± 0.15 G | tr | tr | 0.57 ± 0.00 G |
| 48 | 4-Methoxyindol-3-ylmethyl GSL (4-methoxyglucobrassicin) a | C17H22N2O7S | 398 | 421 | 3.23 ± 0.43 A | 0.15 ± 0.00 D | 0.09 ± 0.00 | 4.23 ± 0.12 B | 0.15 ± 0.00 D | 0.10 ± 0.00 | 3.80 ± 0.43 A,B | 0.53 ± 0.12 E | 0.45 ± 0.00 |
| [X] | Hydroxymethoxy- glucobrassicin | C17H22N2O8S | 414 | 437 | tr | tr | 0.24 ± 0.08 G | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.02 ± 0.00 H | n.d. | n.d. | 0.03 ± 0.00 H |
| 47 | N-Methoxyindol-3-ylmethyl GSL (neoglucobrassicin) a | C17H22N2O7S | 398 | 421 | 1.97 ± 0.31 A | 0.20 ± 0.05 D | 0.04 ± 0.00 G | 2.73 ± 0.33 B | 0.56 ± 0.12 E | tr | 2.43 ± 0.12 A,B | 0.20 ± 0.00 D | 0.18 ± 0.00 G |
| 138 | 1,4-Dimethoxyindol-3-ylmethyl GSL (1,4-dimethoxyglucobrassicin) b | C18H24N2O8S | 428 | 451 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | tr | n.d. | n.d. |
| Total (µmol/g DW) | 13.48 ± 1.82 J | 1.66 ± 0.11 M,N | 1.28 ± 0.32 P,R | 20.57 ± 1.02 K | 2.21 ± 0.37 M,N | 0.72 ± 0.15 P,R | 22.34 ± 2.29 K | 1.42 ± 0.24 M | 1.81 ± 0.12 P | ||||
| No. | Glucosinolate (GSL) | Formula (desulfo) | Desulfo M | Desulfo [M + Na]+ | Reference | Na2SeO4 solution | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 mM | 5 mM | |||||||||
| Root | Aerial Part | Root | Aerial Part | Root | Aerial Part | |||||
| Met derived | ||||||||||
| 64 | 4-(Methylsulfinyl)butyl GSL (glucoraphanin) a | C12H23NO7S2 | 357 | 380 | 3.23 ± 0.63 A | 0.87 ± 0.22 D | 1.69 ± 0.34 B | 0.95 ± 0.11 D | 0.75 ± 0.15 C | 0.74 ± 0.12 D |
| 135 | 4-(β-d-Glucopyranosyldisulfanyl)butyl GSL (diglucothiobeinin) b | C17H31NO11S3 | 521 | 544 | tr | 1.14 ± 0.24 | n.d. | n.d. | n.d. | n.d. |
| 95 | 3-(Methylsulfanyl)propyl GSL (glucoibervirin) b | C11H21NO6S2 | 327 | 350 | 1.88 ± 0.33 | 1.88 ± 0.17 | n.d. | n.d. | n.d. | n.d. |
| 84 | 4-(Methylsulfanyl)butyl GSL (glucoerucin) a | C12H23NO6S2 | 341 | 364 | 20.59 ± 1.32 A | 2.78 ± 0.33 D | 17.27 ± 1.12 A | 1.30 ± 0.08 E | 4.50 ± 0.55 B | 0.21 ± 0.05 F |
| 134 | Dimeric 4-mercaptobutyl GSL b | C22H40N2O12S4 | 652 | 675 | 4.33 ± 0.62 A | 0.20 ± 0.00 D | 17.14 ± 0.93 B | 1.49 ± 0.22 E | 6.78 ± 0.77 C | 0.55 ± 0.12 F |
| 65 | 10-(Methylsulfinyl)decyl GSL (glucocamelinin) a | C18H35NO7S2 | 441 | 464 | tr | 0.21 ± 0.12 | tr | tr | n.d. | tr |
| Se-Met derived | ||||||||||
| [145] | 4-(Methylseleno)butyl GSL (glucoselenoerucin) | C12H23NO6SSe | 389 | 412 | n.d. | n.d. | 4.48 ± 0.55 A | 1.24 ± 0.33 D | 0.78 ± 0.29 B | 0.36 ± 0.10 E |
| Trp derived | ||||||||||
| 28 | 4-Hydroxyindol-3-ylmethyl GSL (4-Hydroxyglucobrassicin) a | C16H20N2O7S | 384 | 407 | 0.23 ± 0.00 | n.d. | tr | n.d. | n.d. | n.d. |
| 48 | 4-Methoxyindol-3-ylmethyl GSL (4-Methoxyglucobrassicin) a | C17H22N2O7S | 398 | 421 | 1.16 ± 0.22 A | 5.96 ± 1.03 D | 0.80 ± 0.11 B | 1.64 ± 0.36 E | 0.07 ± 0.00 C | 0.57 ± 0.12 F |
| 138 | 1,4-Dimethoxyindol-3-ylmethyl GSL (1,4-dimethoxyglucobrassicin) b | C18H24N2O8S | 428 | 451 | 0.92 ± 0.12 A | 0.20 ± 0.00 | 2.06 ± 0.18 B | 0.15 ± 0.00 | 1.33 ± 0.31 A | 0.04 ±0.00 |
| Total (µmol/g DW) | 32.34 ± 3.24 G | 13.24 ± 2.11 J | 43.44 ± 3.23 H | 6.77 ± 1.10 K | 14.21 ± 2.07 I | 2.47 ± 0.51 L | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đulović, A.; Usanović, K.; Kukoč Modun, L.; Blažević, I. Selenium Biofortification Effect on Glucosinolate Content of Brassica oleracea var. italic and Eruca vesicaria. Molecules 2023, 28, 7203. https://doi.org/10.3390/molecules28207203
Đulović A, Usanović K, Kukoč Modun L, Blažević I. Selenium Biofortification Effect on Glucosinolate Content of Brassica oleracea var. italic and Eruca vesicaria. Molecules. 2023; 28(20):7203. https://doi.org/10.3390/molecules28207203
Chicago/Turabian StyleĐulović, Azra, Katarina Usanović, Lea Kukoč Modun, and Ivica Blažević. 2023. "Selenium Biofortification Effect on Glucosinolate Content of Brassica oleracea var. italic and Eruca vesicaria" Molecules 28, no. 20: 7203. https://doi.org/10.3390/molecules28207203
APA StyleĐulović, A., Usanović, K., Kukoč Modun, L., & Blažević, I. (2023). Selenium Biofortification Effect on Glucosinolate Content of Brassica oleracea var. italic and Eruca vesicaria. Molecules, 28(20), 7203. https://doi.org/10.3390/molecules28207203