Abstract
Pre-harvest sprouting is a frequent problem for wheat culture that can be simulated by laboratory-based germination. Despite reducing baking properties, wheat sprouting has been shown to increase the bioavailability of some nutrients. It was investigated whether wheat cultivars bearing distinct grain texture characteristics (BRS Guaraim, soft vs. BRS Marcante, hard texture) would have different behavior in terms of the changes in phytochemical compounds during germination. Using LC-Q-TOF-MS, higher contents of benzoxazinoids and flavonoids were found in the hard cultivar than in the soft one. Free phytochemicals, mainly benzoxazinoids, increased during germination in both cultivars. Before germination, soft and hard cultivars had a similar profile of matrix-bound phytochemicals, but during germination, these compounds have been shown to decrease only in the hard-texture cultivar, due to decreased levels of phenolic acids (trans-ferulic acid) and flavonoids (apigenin) that were bound to the cell wall through ester-type bonds. These findings confirm the hypothesis that hard and soft wheat cultivars have distinct behavior during germination concerning the changes in phytochemical compounds, namely the matrix-bound compounds. In addition, germination has been shown to remarkably increase the content of benzoxazinoids and the antioxidant capacity, which could bring a health-beneficial appeal for pre-harvested sprouted grains.
1. Introduction
Wheat (Triticum aestivum L.) is among the most-produced cereals in the world. Wheat production is affected by climate changes [1]. Under conditions of high humidity and temperature, cereal crops suffer a process of natural germination before harvest. This process, which is known as pre-harvest sprouting, reduces grain quality and generates economic losses worldwide [1]. Sprouted wheat is usually discarded due to the loss of its baking properties [2,3,4] or it can be destined for uses other than baking [5,6], such as for feed or industrial uses (production of starch, vital gluten, furfural, ethanol, etc.).
In contrast, grain germination has been also used as a natural method of biological processing to improve the nutritional, functional and sensory properties of grains along with increasing micronutrient content [7]. Grain germination can increase the content of health-beneficial phytochemicals, such as phenolic compounds [8,9] and benzoxazinoids [10]. Phenolic compounds have antioxidant, antidiabetic and antitumor properties, and have been shown to prevent cardiovascular diseases [11]. Benzoxazinoids exhibit antimicrobial properties, act as central nervous system stimulators, immunoregulators, appetite inhibitors and body weight reducers [10].
Phenolic compounds are found in the form of glycosides linked to different sugar fractions, or in other forms linked to organic acids, amines, lipids, carbohydrates and other phenols, mainly phenolic acids [12]. Cereal phenolic acids occur in free (~25%) or bound (~75%) forms [13], being linked through ester or ether bonds to cell wall polysaccharides [14].
Benzoxazinoids are a class of natural products that are widely distributed in cereals, being concentrated in the cover layer of grains (pericarp) [15]. They can be divided into three groups, namely hydroxamic acids, lactams and benzoxazolinones [16], and have been shown to be increased during the germination of some cereal grains [17], but their behavior during wheat germination remains unknown.
The phytochemical compounds found in the free state are easily extracted with conventional organic solvents [18], whereas those linked to sugars and proteins, or cell wall structures, require very hard conditions for their extraction, such as acid or alkaline hydrolysis [19,20]. Grain germination is a physiological process that can trigger both the hydrolysis of matrix-bound phytochemical compounds and the formation of new compounds [3,21,22].
Therefore, wheat that is naturally germinated before harvest could have a higher content of phytochemical compounds, which would improve its functional quality and, thus, add value to this product. In fact, germination can increase the bioavailability of nutrients, such as vitamins, bioelements and other biologically active substances, due to the partial hydrolysis of starch, proteins, hemicelluloses and celluloses. In this process, hydrolytic enzymes are activated (endohydrolase such as α and β-amylases, proteases, diphenoloxidase and catalase) that break down starch, fibers and proteins, and lead to an increase in the number of digestible compounds along with an improvement in functional properties [23]. Laboratory-based germination that simulates field humidity and temperature conditions can be used as a model to study the influence of pre-harvest sprouting on wheat grain properties [2,5,6].
Another important property beneficial to health is the antioxidant capacity of cereals. Antioxidant compounds protect cells against oxidative stress caused by reactive species [23]. The main compounds responsible for such effects in cereals are vitamins, sterols, and phenolic compounds. They all contribute to some extent to the antioxidant properties and are affected in different ways by germination. Phenolic acids have potential antioxidant properties due to the presence of an aromatic phenolic ring. Its antioxidant properties are explained by the donation of electrons and the transfer of the hydrogen atom to free radicals. They act as free radical scavengers, reducing agents and inhibitors of singlet oxygen formation. Benzoxazinoids also exhibit antioxidant properties that are related to the presence of a hydroxyl group attached to the heterocyclic nitrogen atom, mainly in hydroxamic acids [24].
Wheat cultivars exhibit physiological differences and can be classified according to the grain hardness (method 55-31.01, AACCI, [25]). Wheat grains cultivated in Brazil are similar to the hard red spring wheat from the United States, but wheat production occurs only in the winter. BRS Marcante is a Brazilian wheat cultivar that has hard texture and exhibits high values of grain hardness index (GHI > 67), gluten strength (W > 275 × 10−4 J) [26], dough development time (DDT > 21.5 min) and stability (>30.2 min) [2], being indicated mainly to produce different kinds of bread and pasta. In contrast, BRS Guaraim has much lower values for these markers, exhibiting a soft grain texture (GHI < 46), W (<206 × 10−4 J) and DDT (<4.1 min) [27] and is more suitable for cake and cookie production.
The hypothesis of this study is that wheat cultivars bearing distinct wheat grain texture characteristics would have different behavior in terms of the changes in phytochemical compounds during germination. In this sense, the objective of this study was to investigate the effect of germination on the profile of phytochemical compounds and antioxidant capacity of Brazilian wheat cultivars that have soft or hard grain texture.
2. Results and Discussion
2.1. Grain Characteristics, Alveography and Germination Follow-Up in Wheat Cultivars
The wheat cultivars selected for the study have different technological characteristics. BRS Marcante cultivar presents higher values for grain texture (GHI), GFN, hectoliter weight and thousand kernel weight compared to BRS Guaraim (111%, 62%, 8% and 12% higher, respectively; Supplementary materials, Table S1). Additionally, the alveograph data were also higher for BRS Marcante cultivar, as observed for gluten strength, tenacity and tenacity/extensibility ratio parameters (164%, 101% and 49% higher, respectively; Supplementary materials, Table S1), in relation to BRS Guaraim cultivar.
Grain hardness is the single most important trait in determining technological properties and end-use quality of wheat products. Hard-textured wheats have more glutenins, whereas soft-textured wheats have more gliadins [28]. The greater content of glutenins leads to increased gluten strength, elasticity and extensibility of the dough.
The germination process conditions used in this study resulted in small sprouts at 24 h (Supplementary materials, Table S2). From 48 h onwards, the grains began to emit small rootlets that became larger at 72 h, but the emergence of the seedling did not occur (Supplementary materials, Table S2). Germination is characterized by the activation of enzymes, including α-amylase, which is responsible for the degradation of starch into sugars that provide energy for the embryo’s growth [29]. GFN was assessed to characterize changes in amylase activity during wheat germination (Table 1). Before germination, GFN was higher for BRS Marcante than BRS Guaraim (p < 0.05; Table 1), indicating lower amylase activity for BRS Marcante.

Table 1.
Germination follow-up through grain falling number values (GFN, expressed in seconds) of wheat cultivars from soft (BRS Guaraim) and hard (BRS Marcante) texture.
As expected, the germination process decreased GFN, indicating increased amylolytic activity, which reduces starch content and gelatinization capacity [30]. The increase in amylolytic activity was faster for BRS Marcante than BRS Guaraim, as indicated by the lower GFN values at 24 and 48 h (p < 0.05; Table 1). The increase in grain moisture during germination causes a gradual decrease in the falling number and in the grain hardness index [31]. This causes the texture of the durum wheat grain to become softer after germination, possibly due to a decrease in the starch crystallinity [32]. This alteration facilitates the enzymatic attack and decrease of GFN with the advance of germination.
2.2. Tentative Identification of Phytochemical Compounds
Thirty-two phytochemical compounds, comprising amino acids, phenolic acids, flavonoids and benzoxazinoids, were identified in BRS Marcante and BRS Guaraim wheat cultivars based on the data provided by LC-Orbitrap-MS and LC-Q-TOF-MS analysis (Supplementary materials, Figure S1 and Table S3). Eighteen compounds were in the free state, whereas fourteen compounds were extracted only after alkaline hydrolysis (eight compounds), indicating that they were linked to the cell wall or simple sugars through ester-type bonds, or after acid hydrolysis (six compounds), indicating that they were linked to polysaccharides through ether-type glycosidic bonds [14]. Previous studies have already reported the presence of phenolic compounds and flavonoids [33,34,35,36], amino acids [37] and benzoxazinoids [36,38,39] in wheat.
The greatest diversity of compounds was identified in the free phytochemical fraction. Amino acids were identified in LC-Q-TOF-MS analysis in the positive ionization mode. Phenylalanine (RT 6.3 min) showed a protonated MS spectrum [M + H]+ at m/z 166 and MS2 fragments at m/z 120 e 103. The main fragmentation pathway of phenylalanine starts from the loss of H2O + CO to form a fragment ion at m/z 120.0851, and a fragment ion at m/z 103.0590 was formed by the additional loss of NH3. Tryptophan showed a protonated MS spectrum [M + H]+ at m/z 205 and MS2 fragments at m/z 118, 143, 146, 144 and 115. The fragmentation product m/z 146.0636 was obtained after the loss of NH3 and CH2CO. This fragment further dissociated to form the fragmented ion at m/z 118.0717 after the loss of CO. The fragment at m/z 144.0827 was formed after the loss of NH3 and CO2, and the dissociation of •H forms the fragment m/z 143.0776. The fragmented ion at m/z 115.0588 was formed after the sequential losses of NH3, H2O, CO and HCN. In addition, amino acids showed MS spectrum and MS2 fragmentation patterns similar to previously reported data [37].
Among benzoxazinoids, two compounds that belong to the hydroxamic acids subclass were identified, namely DIBOA-hex-hex and DIMBOA-hex-hex. At 17.2 min, the compound was tentatively identified as DIBOA-hex-hex (MW = 505). In the negative ionization mode, the MS spectrum showed the deprotonated molecule [M−H]− at m/z 504 and MS2 fragment at m/z 162 [M−H−342]−, corresponding to the loss of two hexose molecules (C12H22O11), and at m/z 134 [M −H−370]−, corresponding to the sequential loss of CO. At 24.3 min, the compound was tentatively identified as DIMBOA-hex-hex (MW = 535). In the negative ionization mode, the MS spectrum showed the deprotonated molecule [M−H]− at m/z 534 and MS2 fragment at m/z 192 [M−H−342]−, corresponding to the loss of two hexose molecules, at m/z 164 [M−H−370]−, corresponding to the sequential loss of CO, and at m/z 149 [M−H−385]−, corresponding to the sequential loss of •CH3. In addition, benzoxazinoids exhibited MS spectrum and MS2 fragmentation patterns similar to previously reported data [36,39].
Phenolic acids and some flavonoids from the free fraction were identified by comparison with standards and confirmed using LC-Orbitrap-MS. Four apigenin derivatives were identified using LC-Q-TOF-MS in the negative ionization mode. Compounds eluted at 38.5 and 42.8 min were identified as apigenin-hex-pent I e II (MW = 564). The MS spectrum showed the deprotonated molecule [M−H]− at m/z 563 and fragment in MS2 at m/z 443 [M−H−120]−, corresponding to the loss of C4H8O4, at m/z 353 [M−H−210]−, corresponding to the loss of C7H14O7, and at m/z 383 [M−H−90]−, corresponding to the loss of C8H14O6. Moreover, the MS spectrum and MS2 fragmentation patterns were similar to previously reported data [40]. The compounds eluted at 52.4 and 53.1 min were identified as apigenin-hex-hex-hex I e II (MW = 770). In the negative ionization mode, the MS spectrum showed the deprotonated molecule [M−H]− at m/z 769 and fragment in MS2 at m/z 425 [M−H−344]−, corresponding to the loss of a molecule of C6H10O7 (194) and a molecule of C5H10O5 (150), and the dissociation of •H forms the fragment m/z 426.0868. At m/z 545 [M−H−224]−, corresponding to the loss of C9H12O3 (168) and two molecules of CO (56), the dissociation of •H forms the fragment m/z 546.1238. Moreover, MS spectrum and MS2 fragmentation patterns were similar to previously reported data for apigenin-hex-hex-hex [41,42].
The fraction of phytochemicals extracted through alkaline and acid hydrolysis, which comprises matrix-bound compounds, showed less diversity compared to the free fraction, being composed mainly of phenolic acids and a flavonoid. All compounds were identified by comparison to an authentic standard using LC-Orbitrap-MS analysis. The presence of p-coumaric acid, trans and cis-ferulic acid were also confirmed using LC-Q-TOF-MS analysis.
The compound eluted at 27.8 min was tentatively identified as p-coumaric acid (MW = 164). In the negative ionization mode, the MS spectrum showed the deprotonated molecule [M−H]− at m/z 163 and fragment in MS2 at m/z 119 [M−H−44]−, corresponding to the loss of CO2. Moreover, the MS spectrum and MS2 fragmentation patterns were similar to previously reported data for p-coumaric acid [36].
The compounds eluted at 35.7 and 36.7 min were tentatively identified as trans and cis-ferulic acid (MW = 194). In the negative ionization mode, the MS spectrum showed the deprotonated molecule [M−H]− at m/z 193. Ferulic acid produces a typical negative fragment in MS2 at m/z 133 [M−H−60]− from demethylation and decarboxylation, corresponding to the loss of C2H4O2, and the dissociation of •H forms the fragment m/z 134.0368 [36].
Some peaks could not be identified, but one of them, which was the most abundant in the extract obtained after acid hydrolysis (Figure S1c), had MS characteristics that are not compatible with phenolic or benzoxazinoid compounds.
2.3. Changes in the Phytochemical Profile of Wheat during Germination
The phytochemical profiles of BRS Guaraim and BRS Marcante across 72 h of germination are shown in Table 2 and Table 3. Before germination, most phytochemicals of BRS Guaraim and BRS Marcante cultivars were bound to the grain matrix (64% released by alkaline hydrolysis and 8% released by acid hydrolysis), being composed mainly of phenolic acids, followed by flavonoids (Table 3). Trans-Ferulic acid was the major single compound found in the wheat grains (46–48% of the total phytochemicals) before germination followed by p-coumaric acid (9–12% of total phytochemicals), as previously reported [22,43,44]. The fraction of free phytochemicals amounted to 28% of total phytochemicals from wheat, and was mainly composed of flavonoids, followed by benzoxazinoids and a minor content of phenolic acids (Table 2). Flavonoids mainly occur in C-glycoside forms [42], and glycosylated apigenins were the main flavonoid structures found (Table 2 and Table 3). Benzoxazinoids amounted to almost 10% of free phytochemicals and DIBOA-hex-hex was the predominant form of benzoxazinoids in the wheat (Table 2), as previously reported [45,46].
Table 2.
Changes in free phytochemical compounds (mg·100 g−1 whole wheat flour d.b.) of wheat grains according to the germination time (G) and cultivar (C).
Table 3.
Changes in bound phytochemical compounds (mg·100 g−1 whole wheat flour d.b.) of wheat grains according to the germination time (G) and cultivar (C).
Free phenolic compounds are generally synthesized in the endoplasmic reticulum and stored in the vacuole of plant cells [19]. Looking at the sum of free phenolic acids, BRS Marcante had the highest levels (p < 0.05), but no changes were detected during germination regardless of the cultivar (Table 2). Sinapic acid was the major free phenolic acid being found at higher levels in BRS Marcante than BRS Guaraim (82% higher, p < 0.05), and its content was not altered during germination (Table 2). Before germination, vanillic acid was found only in BRS Guaraim, but decreased with the advance of germination, while in BRS Marcante, it showed a trend of increasing (Table 2).
DIBOA-hex-hex was the major benzoxazinoid in both cultivars and was 1.8-fold higher in BRS Marcante than in BRS Guaraim, whereas DIMBOA-hex-hex did not differ between cultivars (Table 2). Previous studies have already reported DIBOA dihexoside as the main benzoxazinoid found in wheat [41,45,47], but benzoxazinoid behavior during wheat germination was still unknown. Benzoxazinoids increased with the advance of germination (Table 2), reaching the highest levels at 72 h (480% higher value than at 0 h; Table 2). This finding is in line with previous reports showing the highest levels of benzoxazinoids in the sprouts of other cereals such as rye [17]. Benzoxazinoids are concentrated in the bran and germ [10], fractions which are mainly involved in germination.
Flavonoids were the major class of compounds in the free phytochemical fraction (~73% of free phytochemicals). Apigenins hex-pent II, hex-hex-hex I and hex-hex-hex II were higher in BRS Marcante than in BRS Guaraim, and this made the sum of flavonoids also higher in BRS Marcante (23% higher). Although most free flavonoids did not change during germination, apigenin (aglycone form) showed a U-shaped behavior that was not affected by cultivar, being decreased up to 48 h, followed by an increase at 72 h of germination (Table 2). BRS Guaraim showed a similar U-shaped behavior for apigenin hex-pent I that was not found in BRS Marcante (p < 0.05; Table 2). The cause of these changes may be related to the binding of apigenin aglycone with other structures, even forming glycosylated apigenins, or the release of cell wall bound apigenins or sugars and proteins [48]. De novo synthesis may also contribute to some of these changes [47]. The sum of free phytochemical compounds, which was 1.3-fold higher in BRS Marcante than BRS Guaraim, was remarkably increased during germination (48 h onwards, p < 0.05; Table 2).
Concerning the bound phytochemical compounds that are released by alkaline hydrolysis, there was no difference between cultivars in the sum of bound phenolic acids, but the content of sinapic acid released by alkaline hydrolysis was higher in BRS Marcante than BRS Guaraim (16% higher content, Table 3). Independent of the cultivar, there was a bell-shaped behavior for sinapic acid content during germination, with peak values at 48h of germination. Bound 4-hydroxybenzoic acid released by alkaline hydrolysis was linearly increased with the advance of germination in BRS Guaraim, while BRS Marcante showed a bell-shaped behavior with peak values at 48 h of germination (p < 0.05; Table 3). Bound vanillic acid released by alkaline hydrolysis, which was not detected in any wheat cultivar before germination, was found only in germinated grains of BRS Guaraim and exhibited a bell-shaped behavior with peak values at 48 h (Table 3). In the free fraction of this cultivar, it appeared only before germination. Free vanillic acid of BRS Guaraim may have been linked to complex cell wall structures through ester-type bonds during the beginning of germination, explaining the disappearance of the free form and its appearance after 48 h in the fraction released after alkaline hydrolysis [14]. Bound p-coumaric acid that was released by alkaline hydrolysis did not differ between cultivars or germination times (Table 3). trans-Ferulic acid was the major component (70%) among the bound phytochemicals released after alkaline hydrolysis (Table 3). trans-Ferulic acid and the sum of phenolic acids released by alkaline hydrolysis had a U-shaped behavior throughout germination in BRS Marcante, but not in BRS Guaraim (Table 3). The lowest content of trans-ferulic acid and of the sum of phenolic acids was found for BRS Marcante at 48 h, resulting in 1.8-fold lower content than BRS Guaraim.
Apigenin was the only flavonoid quantified in the bound fraction released after alkaline hydrolysis. In BRS Marcante, apigenin underwent a significant decrease at 72 h of germination (Table 3), which was not observed in BRS Guaraim.
Although there was no difference in the sum of bound phytochemicals released by alkaline hydrolysis between cultivars, the level of this group of compounds decreased in BRS Marcante from 48 h onwards, resulting in lower levels than BRS Guaraim (41% lower levels, Table 3). One explanation for this behavior is the release of compounds bound to the cell wall during germination, which increases the content of free compounds [48]. Accordingly, the sum of free phenolic compounds increased during germination.
In the fraction of bound compounds that were released by acid hydrolysis, only p-coumaric acid was identified (Table 3). There was a significant cultivar vs. germination time interaction in the levels of p-coumaric acid released by acid hydrolysis, resulting in higher levels for BRS Marcante than BRS Guaraim at 24 h of germination (25% higher levels, p < 0.05). However, no marked changes were observed in the content of this phenolic acid throughout germination. Kim, Kwak and Kim [44] found gallic, 4-hydroxybenzoic, vanillic, caffeic, syringic, ferulic and p-coumaric acids in the free and bound fractions of germinated and not-germinated wheat. Initially, they observed low levels followed by an increase with the advance of germination, and the highest levels were found at 72 h and 96 h of germination. Ferulic and vanillic acid had the highest increase with values up to 1.5-fold higher in germinated than in not-germinated grains. Most phenolic acids in wheat occur in the bound form and have been shown to increase with advancing germination, i.e., p-coumaric, ferulic and sinapic acids, likely through the decomposition of lignin and other chemical reactions [43,44].
2.4. Multivariate Analysis of Phytochemical Changes in during Wheat Germination
MANOVA using the likelihood ratio test (Wilks), Pillai, Hotelling–Lawley and Roy tests revealed that, when all dependent variables (phytochemical compounds) were combined in the analysis, there was a significant interaction effect of wheat cultivar vs. germination time (Supplementary materials, Table S4). This finding confirms the hypothesis that the soft and hard wheat cultivars evaluated showed distinct changes in free and bound phytochemical compounds during germination, and qualifies the data for the Principal Component Analysis (PCA).
Cluster analysis further confirmed the distinct behavior of BRS Guaraim and BRS Marcante concerning the changes in phytochemicals during germination. Cluster analysis revealed that wheat cultivars were grouped into three groups according to the free phytochemical variables during germination (Figure 1(a1)), explaining 63.3% of the data variation. BRS Marcante was grouped into a single group regardless of the germination time (M-0 h, -24 h, -48 h and -72 h), whereas BRS Guaraim was divided into two groups: one for 0 and 24 h (G-0 h and -24 h) and the other for 48 h and 72 h (G-48 h and -72 h). Cluster analysis of the free phytochemical variables (Figure 1(a2)) also revealed three groups, which were able to explain 73% of the data variation. One of these groups included vanillic acid and DIBOA-hex-hex, another group was composed of DIMBOA-hex-hex, apigenin-hex-pent I and II and sinapic acid, and the last one was composed of apigenins hex-hex-hex I, II and aglycone.
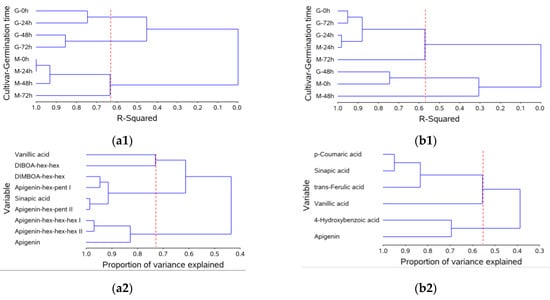
Figure 1.
Dendrogram of wheat cultivars (G = Guaraim, M = Marcante) under different germination times (0, 24, 48 and 72 h; ordinate axis) in relation to the coefficient of determination (r2, abscissa axis) using Euclidean distance as a measure of dissimilarity and Ward’s agglomerative hierarchical algorithm as a clustering method for the free phytochemical fraction (a1) and for the bound phytochemical fraction that was released after alkaline hydrolysis (b1); and dendrogram of phytochemical content (mg/100 g, ordinate axis) in relation to the coefficient of determination (r2, abscissa axis) using the correlation matrix as a measure of similarity and the principal component as a clustering method for the free phytochemical fraction (a2) and for the bound phytochemical fraction that was released after alkaline hydrolysis (b2). a1 = 63.3% and a2 = 73.0%; b1 = 57.1% and b2 = 54.2%.
The cluster analysis of wheat cultivars during germination according to the bound phytochemicals that were released after alkaline hydrolysis divided samples into three groups (57.1%) (Figure 1(b1)). The first group included BRS Guaraim 0, 24 and 72 h (G-0 h, -24 h and -72 h) and BRS Marcante 24 and 72 h (M-24 h and -72 h), the second group included BRS Guaraim 48 h and BRS Marcante 0 h (G -48 h and M-0 h) and the third group was represented only by BRS Marcante 48 h (M-48 h). The dependent variables (bound phytochemical compounds) of this analysis, in turn, were only divided into two groups (54.2%), one comprising p-coumaric, sinapic, trans-ferulic and vanillic acid, and the other comprising 4-hydroxybenzoic acid and apigenin (Figure 1(b2)).
PCA was used as an exploratory analysis to verify if changes in the phytochemical compounds of free or bound fractions would allow the discrimination of wheat cultivars during germination (Figure 2). A biplot (cultivar and germination time vs. content of free phytochemicals, Figure 2a) confirmed the findings of cluster analysis and increased the proportion of explained variance to 86.6% using the first three principal components. Another biplot (cultivar and germination time vs. content of bound phytochemicals released after alkaline hydrolysis, Figure 2b) confirmed the findings of cluster analysis and increased the proportion of explained variance to 84.5% using the first three principal components.
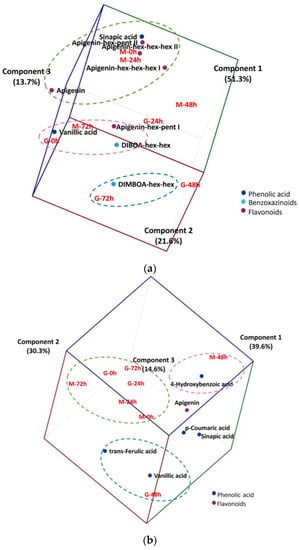
Figure 2.
Three-dimensional biplot of wheat cultivars (G = Guaraim, M = Marcante) at different germination times (0, 24, 48 and 72 h) (scores) versus phytochemicals (loadings) in relation to the main components of principal component analysis for the free phytochemical fraction (a) and for the bound phytochemical fraction that was released after alkaline hydrolysis (b); a = 86.6% and b = 84.5%. Colored dashed ellipses indicate the proximity among samples and variables.
BRS Marcante before germination and up to 24 h of germination was associated with high levels of some free phytochemicals, namely sinapic acid, apigenin and apigenin hex-pent II, and apigenin hex-hex-hex I and II (Figure 2a, green circle). With the progression of germination, BRS Marcante changed the profile of free phytochemical compounds, becoming associated with high levels of vanillic acid, apigenin hex-pent I and DIBOA hex-hex after 72 h of germination (Figure 2a, pink circle). On the other hand, BRS Guaraim was already associated with high levels of vanillic acid, apigenin hex-pent I and DIBOA hex-hex before germination (Figure 2a, pink circle), which were changed to a free phenolic profile associated with DIMBOA hex-hex after 72 h of germination (Figure 2a, blue circle).
Bound phenolic compounds are located mainly in the cell wall and are formed through the conjugation of free phenolic compounds with macromolecules such as cellulose and proteins [22]. Concerning the profile of bound phytochemicals that were released after alkaline hydrolysis, there was an association between BRS Guaraim and BRS Marcante samples at the start (0 and 24 h) and in the end of germination (72 h) (Figure 2b, green circle). However, at 48 h of germination there was a distinct profile of bound phytochemicals between cultivars. While BRS Marcante was associated with high levels of 4-hydroxybenzoic acid (pink circle), BRS Guaraim was associated with high levels of vanillic acid and trans-ferulic acid (blue circle; Figure 2b).
2.5. Antioxidant Capacity of Wheat Cultivars during Germination
During germination, in addition to the hydrolysis of macronutrients such as protein [49] and starch (Table 1), phytochemical compounds may be synthesized, released or conjugated, which may result in changes in the antioxidant capacity (Figure 3). The antioxidant capacity is associated with the ability to scavenge free radicals, break radical chain reactions and chelate metals [50]. The oxygen radical absorbance assay was used to evaluate the antioxidant capacity of wheat grains. The antioxidant capacity was assessed only in the extract containing the free phytochemicals, which is expected to be solubilized during digestion and available for intestinal absorption. Although bound phytochemicals, namely bound phenolic compounds, have already been demonstrated to exert antioxidant effects in vivo [51,52], such effects depend on their interaction and biotransformation by gut microbiota, which was not investigated in the present study.
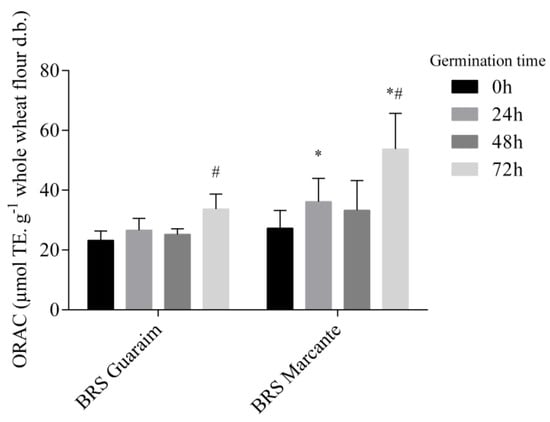
Figure 3.
Changes in the oxygen radical absorbance capacity (ORAC) of the free phytochemical fraction of germinated wheat. # Significantly different (p < 0.05) from the same cultivar at 0 h. * Significantly different (p < 0.05) from BRS Guaraim at the same germination time. n = 6.
Although BRS Marcante had a higher content of free phytochemical compounds than BRS Guaraim (Table 2), the peroxyl radical scavenging capacity of both cultivars was similar before germination (Figure 3). The compounds that most contribute to this antioxidant activity are phenolic acids and flavonoids [53]. In both cultivars, there was an increase in the antioxidant capacity after 72 h of germination (p < 0.05), and this increase was higher for BRS Marcante, which had a higher antioxidant capacity than BRS Guaraim at 24 h (58% higher) and 72 h (41% higher; p < 0.05).
Many studies demonstrate increased antioxidant capacity in sprouted wheat [54,55,56,57,58] and generally attribute this to the release of bound phytochemicals. In contrast to the free phytochemical compounds that are soluble in conventional organic solvent, most phytochemical compounds of wheat are bound to cell wall components or sugars. These matrix-bound phytochemical compounds (BPC) are insoluble and, therefore, will not contribute to the antioxidant capacity assessed in the extracts of wheat grain. However, during food digestion, BPC will reach the colon, where they can be fermented by gut microbiota and exert nutraceutical properties [59]. In addition, BPC could be a substrate for metabolic transformation during grain germination. The diverse and complex structure of BPC makes it difficult to analyze these compounds. For this reason, alkaline and acid hydrolysis were used to release the small units of phytochemical compounds and obtain some insight into the type of chemical bonds responsible for their linkage to the food matrix. The total concentration of phytochemicals bound to the cell wall (ester-type bonds, alkaline hydrolysis) was 2.1- and 1.5-fold higher for BRS Guaraim and BRS Marcante, respectively, than the total concentration of free phytochemicals. Phytochemicals linked to simple sugars (ether-type glycosidic bonds, acid hydrolysis) were 3.6- and 4.5-fold lower, respectively. Of the total phytochemical compounds found in wheat, approximately 70% and 64% correspond to the total bound fraction, respectively, for BRS Guaraim and BRS Marcante. Our result agrees with [13], which says that 60% to 90% of the phytochemical compounds in cereals occur in a bound form.
3. Materials and Methods
3.1. Wheat Samples
Wheat (T. aestivum L.) grains of BRS Marcante and BRS Guaraim cultivars were obtained in the experimental fields of the Brazilian Agricultural Research Corporation (Embrapa Trigo) in Passo Fundo, RS state, Brazil (geographical coordinates: 28°15′46″ S; 52°24′24″ W; 687 m), in two crop seasons.
3.2. Samples Characterization
The GHI (method 55-31.01, AACCI [25], Perten Instruments, Springfield, IL, USA), grain falling number, (GFN, method 56-81.03, AACCI [25], Perten Instruments, Springfield, IL, USA), hectoliter weight (method 55-10.01, AACCI [25] with results expressed in kg·hL−1) and thousand kernel weight [60] were assessed to characterize the grains. The technological quality of wheat cultivars was evaluated according to AACCI [25] and included alveography (method 54-30.02, AACCI [25], Chopin Alveograph, Villeneuve-la-Garenne, France) of whole wheat flours, considering the parameters gluten strength (W), tenacity (P) and tenacity/extensibility ratio (P/L).
3.3. Germination
Wheat grain germination was performed as described by Baranzelli et al. [2] with some modification. Wheat grains were germinated in triplicate under controlled lighting conditions (12 h day/12 h night), relative humidity (80%), grain moisture (30%) and temperature (25 °C day/15 °C night) for0, 24, 48, and 72 h. Germination temperatures were selected according to the average of the maximum (day) and minimum (night) temperatures of the years 2014, 2015 and 2016 in Passo Fundo during the wheat crop season (September to November). This region is among the major Brazilian wheat producers.
3.4. Grain Milling
The whole wheat grains were first ground in a knife mill (Marconi, Piracicaba, SP, Brazil), followed by grinding in a hammer mill (Perten Instruments, Springfield, IL, USA) with a 0.8 mm sieve.
3.5. Germination Marker
The GFN was evaluated according to method 56–81.03 (AACCI [25], Perten Instruments, Springfield, IL, USA) with altitude correction (Passo Fundo, RS, Brazil = 687 m).
3.6. Extraction of Free and Matrix-Bound Phytochemical Compounds
Free phytochemical compounds (FPC) were extracted from 0.25 g of whole wheat flour diluted in 2.5 mL of acetone/water (80:20, v/v) solution acidified with 0.1% formic acid. Samples were vortex-mixed for 2 min, centrifuged at 1500× g for 3 min and the supernatant was collected. Extraction was repeated three times, the supernatants were pooled and 6 mL of extract was concentrated in a rotary evaporator at 38 °C.
Matrix-bound phytochemical compounds (BPC) were extracted through alkaline and acid hydrolysis according to Zhang et al. [61] with adaptations. For alkaline hydrolysis, 2.0 mL of distilled water and 1.5 mL of 3 mol·L−1 NaOH were added to the residue remaining after FPC extraction. Samples were shaken for 16 h at 20 °C and then had the pH adjusted to ~2.0. Phytochemical compounds released were extracted three times with 15 mL of diethyl ether/ethyl acetate solution (1:1, v/v). The supernatants were mixed, and 40 mL was dried in a rotary evaporator at 38 °C. The acid hydrolysis was carried out by adding 1.5 mL of 6 mol·L−1 HCl to the residue remaining after alkaline hydrolysis. Samples were incubated at 85 °C for 30 min and, thereafter, had the pH adjusted to ~2.0. The extraction followed the same steps as described for alkaline hydrolysis.
Extracts of FPC and BPC obtained through alkaline and acidic hydrolysis were resuspended in 1.2 mL of acidified water/methanol solution (1:0.2 v/v, 0.1% formic acid) and stored at −20 °C until further analysis.
3.7. Phytochemical Compounds Identification
The phytochemical compounds of wheat extracts were identified using two different sets of liquid chromatograph (LC) equipment. The first exploratory analysis was conducted using an LC system connected to a mass spectrometer (MS) that had a hybrid Orbitrap analyzer and an electrospray ionization source (ESI, Q-ExactiveTM, Thermo Fisher Scientific, Bremen, Germany), using the chromatographic conditions previously described by Astudillo-Pascual et al. [62].
Thereafter, compound identification was accomplished using an LC-MS equipped with a quadrupole-time-of flight (Q-TOF) analyzer and an electrospray ionization source (ESI) (Bruker Daltonics, micrOTOF-Q III model, Bremen, Germany). Chromatographic parameters were set according to Quatrin et al. [63] and Q-TOF-MS parameters were set according to Mallmann et al. [64].
The identification of phytochemical compounds was based on the retention time and order of elution in the reverse phase column, maximum absorption wavelength (UV-vis) and characteristics of the MS spectrum compared to authentic standards analyzed under the same conditions or with literature data. For the reliable identification of compounds, comparison was made with analytical standards and databases, based on neutral mass isotope distribution, retention time and MS/MS fragments using the customized database of polyphenols from PubChem (https://pubchem.ncbi.nlm.nih.gov/ (accessed on 20 July 2020)) and other metabolites from KEGG (http://www.genome.jp/kegg/ (accessed on 23 July 2020)), MoNA: MassBank of North America (https://mona.fiehnlab.ucdavis.edu/ (accessed on 3 August 2020)), FooDB (https://foodb.ca (accessed on 12 August 2020)) and ReSpect for Phytochemicals (http://spectra.psc.riken.jp/menta.cgi/respect/index (accessed on 13 August 2020)). For the non-targeted identification, the parameters generated by the software were applied in descending order of importance: precursor exact mass error and fragment mass error (<10 ppm); isotopic similarity (>80%) and highest fragmentation score (score > 30).
3.8. Quantification of Phytochemical Compounds
The quantification of free and matrix-bound phytochemical compounds of germinated wheat samples was performed according to Quatrin et al. [63].
Absorption spectra were recorded from 200 to 800 nm, and chromatograms for the quantification of phytochemical compounds were obtained at 280, 320 and 360 nm. Results were expressed as mg·100 g−1 of whole wheat flour on dry basis. The quantification conditions were validated in our laboratory (Nidal-UFSM) (Supplementary materials, Box S1 and Table S5) following the International Conference on Harmonization Guidelines.
3.9. Oxygen Radical Absorbance Capacity (ORAC)
The oxygen radical absorbance capacity (ORAC) of FPC was determined according to the method described by Ou, Hampsch-woodill and Prior [65]. Values were expressed as µmol of Trolox equivalents.g−1 sample on dry basis.
3.10. Statistical Analysis
The results of germination markers and ORAC were analyzed using 2-way analysis of variance (ANOVA, 2 cultivars × 4 germination times) with six replicates (n = 6, where three replicates each were provided from two different crop seasons), followed by Tukey’s test for mean comparison at a level of 5% significance. Statistica software (version 9.0, StatSoft Inc., Tulsa, OK, USA) was used to perform this statistical analysis.
For the results of the phytochemical quantification, the experimental design was randomized into blocks in a 2 × 4 factorial scheme (2 wheat cultivars × 4 germination times), totaling eight (8) treatments with six replicates each (n = 6). Data were submitted to ANOVA using the GLM procedure, their means were adjusted using the ordinary least squares method with the LSMEANS command and compared using the Student–Newman–Keuls (SNK) test.
Then, multivariate analysis of variance (MANOVA) was performed, in which the matrices of sums of squares and products were obtained. To test the hypothesis that the treatment means vectors were null, the Wilks (λ), Pillai (V), Hotelling–Lawley (U) and Roy (F0) tests were performed. Additionally, cluster analysis was performed for treatments using the DISTANCE, CLUSTER and TREE procedures, using the average Euclidean distance as a dissimilarity measure and Ward as the clustering method. A cluster analysis of the dependent variables was also performed using the VARCLUS and TREE procedures, using the correlation matrix as input. Principal component analysis was performed using the PRINQUAL, PRINCOMP and FACTOR procedures (Ravindra Khattree and Dayanand N. Naik 2000). Statistical analyses were performed on the SAS® System for Windows™ version 9.4 (SAS Institute Inc., Cary, NC, USA), at a 5% significance level.
Chromatogram figures were obtained from the LabSolutions software, dendogram figures were obtained from the SAS® software, PCA figures were produced using Microsoft Excel and the antioxidant capacity figure was produced using GraphPad Prism version 6 for Windows (GraphPad Software, San Diego, CA, USA).
4. Conclusions
Germination proved to be a good tool to increase and diversify the content of bioactive compounds in wheat, as indicated by the increase in the content of benzoxazinoids and antioxidant capacity. Changes in the profile of phytochemical compounds during germination were different between the soft- and hard-texture cultivars, and the hard-texture cultivar had the greatest increase in antioxidant capacity after germination. Before germination, the hard-texture cultivar, BRS Marcante, had a higher content of total benzoxazinoids and flavonoids than the soft-texture cultivar, BRS Guaraim, including DIBOA-hex-hex, free and bound sinapic acid, and apigenins hex-pent II, and hex-hex-hex I and II. On the other hand, the soft-texture cultivar, BRS Guaraim, had a greater content of DIMBOA-hex-hex. Free phytochemical compounds, mainly benzoxazinoids, increased during germination in both cultivars. Before germination, few differences were observed between the soft and hard cultivar concerning the profile of matrix-bound phytochemicals, which comprise the major group of phenolic compounds in wheat. During germination, the levels of this group of compounds have been shown to decrease only in the hard-texture cultivar, due to decreased levels of phenolic acids (trans-ferulic acid) and flavonoids (apigenin) that were bound to the cell wall through ester-type bonds. These findings confirm the hypothesis that hard and soft wheat cultivars have distinct behavior during germination concerning the changes in phytochemical compounds, namely the matrix-bound compounds. In addition, germination has been shown to remarkably increase the content of benzoxazinoids, and the antioxidant capacity of the free phytochemical fraction of wheat, which could bring a health-beneficial appeal for pre-harvested sprouted grains that have lost their baking properties.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules28020721/s1. Table S1. Technological characteristics of two Brazilian wheat cultivars bearing soft (BRS Guaraim) and hard (BRS Marcante) texture grains. Table S2. Grain images during germination. Figure S1. Representative LC-PDA chromatograms of phytochemicals extracted from wheat grains. Free compounds (a) and bound compounds obtained by alkaline (b) and acid (c) hydrolysis. Table S3. Tentative identification of wheat phytochemicals by LC-Q-TOF-MS and LC-Orbitrap-MS. Table S4. Results of multivariate analysis of variance (MANOVA) for phytochemical compounds from wheat cultivars (C) under different germination times (T), using likelihood ratio test (Wilks) and Pillai, Hotelling-Lawley and Roy’s tests. Box S1. Validation data for the analysis of benzoxazinone and phenolic compounds. Table S5. Accuracy test for the analysis of phenolic and benzoxazinoid compounds in wheat grains.
Author Contributions
Conceptualization, J.B. and T.E.; data curation, R.d.O.M., E.R. and O.D.P.; formal analysis, J.B., C.S.M., R.d.O.M., E.R., O.D.P., R.L.-R., A.G.F., R.R.-G. and M.Z.d.M.; funding acquisition, T.E.; investigation, S.S., M.Z.d.M. and T.E.; methodology, J.B., S.S., C.S.M., R.d.O.M., E.R., O.D.P. and M.Z.d.M.; project administration, M.Z.d.M. and T.E.; resources, T.E.; supervision, S.S., M.Z.d.M. and T.E.; validation, J.B. and S.S.; writing—original draft, J.B.; writing—review and editing, J.B., S.S., M.Z.d.M. and T.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CAPES (Coordenação de Aperfeiçoamento Pessoal de Nível Superior), grant number 001 and CNPq (National Council of Scientific and Technological Development) grant numbers 435932/2018-7 and 309604/2021-4.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Enio Marchesan and his team from the Phytotechnics Department of the Federal University of Santa Maria for loaning the germination chambers.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, C.; Nonogaki, H.; Barrero, J. Seed Dormancy, Germination and Pre-Harvest Sprouting; Balestrazzi, A., Ed.; University of Pavia: Pavia, Italy, 2019; Volume 9, ISBN 9782889457625. [Google Scholar]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; de Miranda, M.Z.; Zavareze, E.d.R.; Dias, A.R.G. Changes in Enzymatic Activity, Technological Quality and Gamma-Aminobutyric Acid (GABA) Content of Wheat Flour as Affected by Germination. LWT-Food Sci. Technol. 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Olaerts, H.; Courtin, C.M. Impact of Preharvest Sprouting on Endogenous Hydrolases and Technological Quality of Wheat and Bread: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Olaerts, H.; Roye, C.; Derde, L.J.A.; Sinnaeve, G.; Meza, W.R.; Bodson, B.; Courtin, C.M. Impact of Preharvest Sprouting of Wheat (Triticum Aestivum) in the Field on Starch, Protein and Arabinoxylan Properties. J. Agric. Food Chem. 2016, 64, 8324–8332. [Google Scholar] [CrossRef] [PubMed]
- Baranzelli, J.; Kringel, D.H.; Mallmann, J.F.; Bock, E.; el Halal, S.L.M.; Prietto, L.; Zavareze, E.d.R.; de Miranda, M.Z.; Dias, A.R.G. Impact of Wheat (Triticum aestivum L.) Germination Process on Starch Properties for Application in Films. Starch/Staerke 2019, 71, 1800262. [Google Scholar] [CrossRef]
- Kringel, D.H.; Baranzelli, J.; Schöffer, J.D.N.; el Halal, S.L.M.; de Miranda, M.Z.; Dias, A.R.G.; Zavareze, E.D.R. Germinated Wheat Starch as a Substrate to Produce Cyclodextrins: Application in Inclusion Complex to Improve the Thermal Stability of Orange Essential Oil. Starch/Staerke 2020, 72, 1–7. [Google Scholar] [CrossRef]
- Hefni, M.; Witthöft, C.M. Increasing the Folate Content in Egyptian Baladi Bread Using Germinated Wheat Flour. LWT-Food Sci. Technol. 2011, 44, 706–712. [Google Scholar] [CrossRef]
- Sibian, M.S.; Saxena, D.C.; Riar, C.S. Effect of Germination on Chemical, Functional and Nutritional Characteristics of Wheat, Brown Rice and Triticale: A Comparative Study. J. Sci. Food Agric. 2017, 97, 4643–4651. [Google Scholar] [CrossRef]
- Nelson, K.; Mathai, M.L.; Ashton, J.F.; Donkor, O.N.; Vasiljevic, T.; Mamilla, R.; Stojanovska, L. Effects of Malted and Non-Malted Whole-Grain Wheat on Metabolic and Inflammatory Biomarkers in Overweight/Obese Adults: A Randomised Crossover Pilot Study. Food Chem. 2016, 194, 495–502. [Google Scholar] [CrossRef]
- Adhikari, K.B.; Tanwir, F.; Gregersen, P.L.; Steffensen, S.K.; Jensen, B.M.; Poulsen, L.K.; Nielsen, C.H.; Høyer, S.; Borre, M.; Fomsgaard, I.S. Benzoxazinoids: Cereal Phytochemicals with Putative Therapeutic and Health-Protecting Properties. Mol. Nutr. Food Res. 2015, 59, 1324–1338. [Google Scholar] [CrossRef]
- Kadiri, O. A Review on the Status of the Phenolic Compounds and Antioxidant Capacity of the Flour: Effects of Cereal Processing. Int. J. Food Prop. 2017, 20, S798–S809. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, E.; Moroni, A.v.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.-A.; van den Broeck, H.C.; Brouns, F.J.P.H.; de Brier, N.; et al. Impact of Cereal Seed Sprouting on Its Nutritional and Technological Properties: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 305–328. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound Phenolics in Foods, a Review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Kowalska, I.; Jędrejek, D. Benzoxazinoid and Alkylresorcinol Content, and Their Antioxidant Potential, in a Grain of Spring and Winter Wheat Cultivated under Different Production Systems. J. Cereal Sci. 2020, 95, 103063. [Google Scholar] [CrossRef]
- de Bruijn, W.J.C.; Gruppen, H.; Vincken, J.P. Structure and Biosynthesis of Benzoxazinoids: Plant Defence Metabolites with Potential as Antimicrobial Scaffolds. Phytochemistry 2018, 155, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Tanwir, F.; Dionisio, G.; Adhikari, K.B.; Fomsgaard, I.S.; Gregersen, P.L. Biosynthesis and Chemical Transformation of Benzoxazinoids in Rye during Seed Germination and the Identification of a Rye Bx6-like Gene. Phytochemistry 2017, 140, 95–107. [Google Scholar] [CrossRef]
- Lattanzio, V. Phenolic Compounds: Introduction. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Merillon, J.-M., Ramawat, K.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 1, pp. 1543–1580. ISBN 9783642221446. [Google Scholar]
- Saura-Calixto, F. Concept and Health-Related Properties of Nonextractable Polyphenols: The Missing Dietary Polyphenols. J. Agric. Food Chem. 2012, 60, 11195–11200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, S.; Ge, S.; Lin, S. Review of Distribution, Extraction Methods, and Health Benefits of Bound Phenolics in Food Plants. J. Agric. Food Chem. 2020, 68, 3330–3343. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted Grains: A Comprehensive Review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef]
- Gan, R.Y.; Lui, W.Y.; Wu, K.; Chan, C.L.; Dai, S.H.; Sui, Z.Q.; Corke, H. Bioactive Compounds and Bioactivities of Germinated Edible Seeds and Sprouts: An Updated Review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, S. Bioactive Components and Functional Properties of Biologically Activated Cereal Grains: A Bibliographic Review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3051–3071. [Google Scholar] [CrossRef]
- Makowska, B.; Bakera, B.; Rakoczy-Trojanowska, M. The Genetic Background of Benzoxazinoid Biosynthesis in Cereals. Acta Physiol. Plant 2015, 37, 176. [Google Scholar] [CrossRef]
- AACCI. American Association of Cereal Chemists, 11th ed.; Approved Methods of the AACC International: St. Paul, MI, USA, 2010. [Google Scholar]
- Caierão, E.; e Silva, M.S.; Scheeren, P.L.; de Castro, R.L.; Eichelberger, L.; do Nascimento Junior, A.; Guarienti, E.M.; de Miranda, M.Z.; Pires, J.L.F.; Maciel, J.L.N.; et al. Resultados Agronômicos e Qualitativos Da Nova Cultivar de Trigo ‘BRS Marcante’. Ciência Rural 2015, 45, 644–646. [Google Scholar] [CrossRef]
- Scheeren, P.L.; Caierão, E.; e Silva, M.S.; de Castro, R.L.; Caetano, V.d.R.; Bassoi, M.C.; Pires, J.L.F.; Eichelberger, L.; de Miranda, M.Z.; Guarienti, E.M.; et al. BRS-Guaraim-Cultivar de Trigo Brando e Farinha Branqueadora. XI Reun. Comissão Bras. Pesqui. Trigo Triticale 2017, 11, 1–5. [Google Scholar]
- Sharma, S.; Katyal, M.; Singh, N.; Singh, A.M.; Ahlawat, A.K. Comparison of Effect of Using Hard and Soft Wheat on the High Molecular Weight-Glutenin Subunits Profile and the Quality of Produced Cookie. J. Food Sci. Technol. 2022, 59, 2545–2561. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, M.; Nonogaki, H. Germination. In Encyclopedia of Applied Plant Sciences; Thomas, B., Ed.; Academic Press: Salem, OR, USA, 2017; Volume 1, pp. 509–512. ISBN 9780123948083. [Google Scholar]
- Olaerts, H.; Roye, C.; Derde, L.J.A.; Sinnaeve, G.; Meza, W.R.; Bodson, B.; Courtin, C.M. Evolution and Distribution of Hydrolytic Enzyme Activities during Preharvest Sprouting of Wheat (Triticum aestivum) in the Field. J. Agric. Food Chem. 2016, 64, 5644–5652. [Google Scholar] [CrossRef]
- Miã, Â.A.; Grundas, S. Wheat Grain Hardness Modified by the Laboratory Sprouting Test. Int. Agrophys. 2012, 16, 283–288. [Google Scholar]
- You, S.-Y.; Oh, S.-G.; Han, H.M.; Jun, W.; Hong, Y.-S.; Chung, H.-J. Impact of Germination on the Structures and in vitro Digestibility of Starch from Waxy Brown Rice. Int. J. Biol. Macromol. 2016, 82, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Zilic, S. Phenolic Compounds of Wheat. Their Content, Antioxidant Capacity and Bioaccessibility. MOJ Food Process. Technol. 2016, 2, 37. [Google Scholar] [CrossRef]
- Gammoh, S.; Alu’datt, M.H.; Alhamad, M.N.; Rababah, T.; Ereifej, K.; Almajwal, A.; Ammari, Z.A.; al Khateeb, W.; Hussein, N.M. Characterization of Phenolic Compounds Extracted from Wheat Protein Fractions Using High-Performance Liquid Chromatography/Liquid Chromatography Mass Spectrometry in Relation to Anti-Allergenic, Anti-Oxidant, Anti-Hypertension, and Anti-Diabetic Propertie. Int. J. Food Prop. 2017, 20, 2383–2395. [Google Scholar] [CrossRef]
- Sharma, M.; Rahim, M.S.; Kumar, P.; Mishra, A.; Sharma, H.; Roy, J. Large-Scale Identification and Characterization of Phenolic Compounds and Their Marker–Trait Association in Wheat. Euphytica 2020, 216, 127. [Google Scholar] [CrossRef]
- Koistinen, V.M.; Hanhineva, K. Mass Spectrometry-Based Analysis of Whole-Grain Phytochemicals. Crit. Rev. Food Sci. Nutr. 2017, 57, 1688–1709. [Google Scholar] [CrossRef]
- Zhang, P.; Chan, W.; Ang, I.L.; Wei, R.; Lam, M.M.T.; Lei, K.M.K.; Poon, T.C.W. Revisiting Fragmentation Reactions of Protonated α-Amino Acids by High-Resolution Electrospray Ionization Tandem Mass Spectrometry with Collision-Induced Dissociation. Sci. Rep. 2019, 9, 6453. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Keski-Rahkonen, P.; Lappi, J.; Katina, K.; Pekkinen, J.; Savolainen, O.; Timonen, O.; Paananen, J.; Mykkänen, H.; Poutanen, K. The Postprandial Plasma Rye Fingerprint Includes Benzoxazinoid-Derived Phenylacetamide Sulfates. J. Nutr. 2014, 144, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, W.J.C.; Vincken, J.P.; Duran, K.; Gruppen, H. Mass Spectrometric Characterization of Benzoxazinoid Glycosides from Rhizopus-Elicited Wheat (Triticum aestivum) Seedlings. J. Agric. Food Chem. 2016, 64, 6267–6276. [Google Scholar] [CrossRef]
- Santos, M.C.B.; Lima, L.R.d.S.; Nascimento, F.R.; do Nascimento, T.P.; Cameron, L.C.; Ferreira, M.S.L. Metabolomic Approach for Characterization of Phenolic Compounds in Different Wheat Genotypes during Grain Development. Food Res. Int. 2018, 124, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Pihlava, J.-M.; Hellstrom, J.; Kurtelius, T.; Mattila, P. Flavonoids, Anthocyanins, Phenolamides, Benzoxazinoids, Lignans and Alkylresorcinols in Rye (Secale cereale) and Some Rye Products. J. Cereal Sci. 2018, 79, 183–192. [Google Scholar] [CrossRef]
- Zhu, Y.; Sang, S. Phytochemicals in Whole Grain Wheat and Their Health-Promoting Effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef]
- Tian, C.; Wang, Y.; Yang, T.; Sun, Q.; Ma, M.; Li, M. Evolution of Physicochemical Properties, Phenolic Acid Accumulation, and Dough-Making Quality of Whole Wheat Flour During Germination Under UV-B Radiation. Front. Nutr. 2022, 1, 877324. [Google Scholar] [CrossRef]
- Kim, M.J.; Kwak, H.S.; Kim, S.S. Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat. Molecules 2018, 23, 2244. [Google Scholar] [CrossRef]
- Tanwir, F.; Fredholm, M.; Gregersen, P.L.; Fomsgaard, I.S. Comparison of the Levels of Bioactive Benzoxazinoids in Different Wheat and Rye Fractions and the Transformation of These Compounds in Homemade Foods. Food Chem. 2013, 141, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Dihm, K.; Lind, M.V.; Sundén, H.; Ross, A.; Savolainen, O. Quantification of Benzoxazinoids and Their Metabolites in Nordic Breads. Food Chem. 2017, 235, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Rogachev, I.; Aura, A.M.; Aharoni, A.; Poutanen, K.; Mykkänen, H. Qualitative Characterization of Benzoxazinoid Derivatives in Whole Grain Rye and Wheat by LC-MS Metabolite Profiling. J. Agric. Food Chem. 2011, 59, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rao, J.; Chen, B. Phenolic Compounds in Germinated Cereal and Pulse Seeds: Classification, Transformation, and Metabolic Process. Crit. Rev. Food Sci. Nutr. 2020, 60, 740–759. [Google Scholar] [CrossRef]
- Zilic, S.; Basic, Z.; Sukalovic, V.H.-T.; Maksimovic, V.; Jankovic, M.; Filipovic, M. Can the Sprouting Process Applied to Wheat Improve the Contents of Vitamins and Phenolic Compounds and Antioxidant Capacity of the Flour? Int. J. Food Sci. Technol. 2014, 49, 1040–1047. [Google Scholar] [CrossRef]
- Maurer, L.H.; Cazarin, C.B.B.; Quatrin, A.; Nichelle, S.M.; Minuzzi, N.M.; Teixeira, C.F.; da Cruz, I.B.M.; Júnior, M.R.M.; Emanuelli, T. Dietary Fiber and Fiber-Bound Polyphenols of Grape Peel Powder Promote GSH Recycling and Prevent Apoptosis in the Colon of Rats with TNBS-Induced Colitis. J. Funct. Foods 2020, 64, 103644. [Google Scholar] [CrossRef]
- Maurer, L.H.; Cazarin, C.B.B.; Quatrin, A.; Minuzzi, N.M.; Nichelle, S.M.; Lamas, C.d.A.; Cagnon, V.H.A.; Morari, J.; Velloso, L.A.; Júnior, M.R.M.; et al. Grape Peel Powder Attenuates the Inflammatory and Oxidative Response of Experimental Colitis in Rats by Modulating the NF-ΚB Pathway and Activity of Antioxidant Enzymes. Nutr. Res. 2020, 76, 52–70. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Junior, M.R.M. Phenolic Compounds: Structure, Classification, and Antioxidant Power. In Bioactive Compounds: Health Benefits and Potential Applications; Elsevier: Amsterdam, The Netherlands, 2019; Volume Chapter 2, pp. 33–50. ISBN 9780128147757. [Google Scholar]
- Naumenko, N.; Potoroko, I.; Kalinina, I. Stimulation of Antioxidant Activity and γ-Aminobutyric Acid Synthesis in Germinated Wheat Grain Triticum Aestivum L. by Ultrasound: Increasing the Nutritional Value of the Product. Ultrason. Sonochem. 2022, 86, 106000. [Google Scholar] [CrossRef]
- Miyahira, R.F.; Pena, F.d.L.; Fabiano, G.A.; Lopes, J.d.O.; Ponte, L.G.S.; da Cunha, D.T.; Bezerra, R.M.N.; Antunes, A.E.C. Changes in Phenolic Compound and Antioxidant Activity of Germinated Broccoli, Wheat, and Lentils during Simulated Gastrointestinal Digestion. Plant Foods Hum. Nutr. 2022, 77, 233–240. [Google Scholar] [CrossRef]
- Yang, B.; Yin, Y.; Liu, C.; Zhao, Z.; Guo, M. Effect of Germination Time on the Compositional, Functional and Antioxidant Properties of Whole Wheat Malt and Its End-Use Evaluation in Cookie-Making. Food Chem. 2021, 349, 129125. [Google Scholar] [CrossRef]
- Dhillon, B.; Choudhary, G.; Sodhi, N.S. A Study on Physicochemical, Antioxidant and Microbial Properties of Germinated Wheat Flour and Its Utilization in Breads. J. Food Sci. Technol. 2020, 57, 2800–2808. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, P.; Weng, Y.; Ma, Y.; Gu, Z.; Yang, R. Comparison of Phenolic Profiles, Antioxidant Capacity and Relevant Enzyme Activity of Different Chinese Wheat Varieties during Germination. Food Biosci. 2017, 20, 159–167. [Google Scholar] [CrossRef]
- Maurer, L.H.; Cazarin, C.B.B.; Quatrin, A.; Minuzzi, N.M.; Costa, E.L.; Morari, J.; Velloso, L.A.; Leal, R.F.; Rodrigues, E.; Bochi, V.C.; et al. Grape Peel Powder Promotes Intestinal Barrier Homeostasis in Acute TNBS-Colitis: A Major Role for Dietary Fiber and Fiber-Bound Polyphenols. Food Res. Int. 2019, 123, 425–439. [Google Scholar] [CrossRef]
- Brasil, M.A.P.A. Regras Para Análise de Sementes; Ministério da Agricultura Pecuária e Abastecimento: Brasília, Brazil, 2009; Volume 1, ISBN 1932-6203.
- Zhang, Y.; Wang, L.; Yao, Y.; Yan, J.; He, Z. Phenolic Acid Profiles of Chinese Wheat Cultivars. J. Cereal Sci. 2012, 56, 629–635. [Google Scholar] [CrossRef]
- Astudillo-Pascual, M.; Domínguez, I.; Aguilera, P.A.; Frenich, A.G. New Phenolic Compounds in Posidonia Oceanica Seagrass: A Comprehensive Array Using High Resolution Mass Spectrometry. Plants 2021, 10, 864. [Google Scholar] [CrossRef]
- Quatrin, A.; Pauletto, R.; Maurer, L.H.; Minuzzi, N.; Nichelle, S.M.; Carvalho, J.F.C.; Maróstica, M.R.; Rodrigues, E.; Bochi, V.C.; Emanuelli, T. Characterization and Quantification of Tannins, Flavonols, Anthocyanins and Matrix-Bound Polyphenols from Jaboticaba Fruit Peel: A Comparison between Myrciaria Trunciflora and M. Jaboticaba. J. Food Compos. Anal. 2019, 78, 59–74. [Google Scholar] [CrossRef]
- Mallmann, L.P.; Tischer, B.; Vizzotto, M.; Rodrigues, E.; Manfroi, V. Comprehensive Identification and Quantification of Unexploited Phenolic Compounds from Red and Yellow Araçá (Psidium cattleianum Sabine) by LC-DAD-ESI-MS/MS. Food Res. Int. 2020, 131, 108978. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).