
Spin-Label Electron Paramagnetic Resonance Spectroscopy Reveals Effects of Wastewater Filter Membrane Coated with Titanium Dioxide Nanoparticles on Bovine Serum Albumin


 ,
,  , and
, and 
Abstract
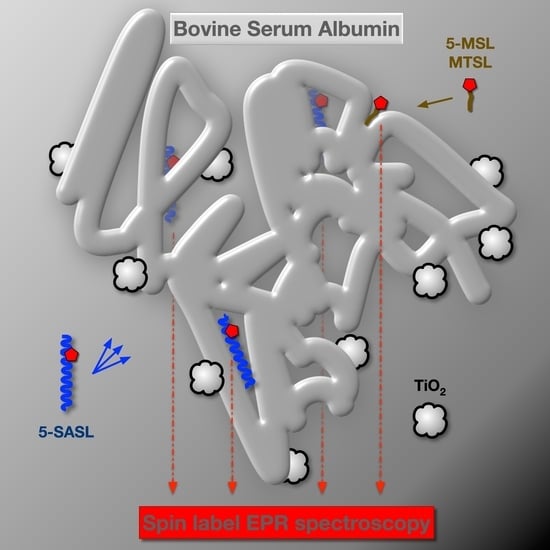
:
1. Introduction
2. Results and Discussion
2.1. Chemical Oxygen Demand (COD)
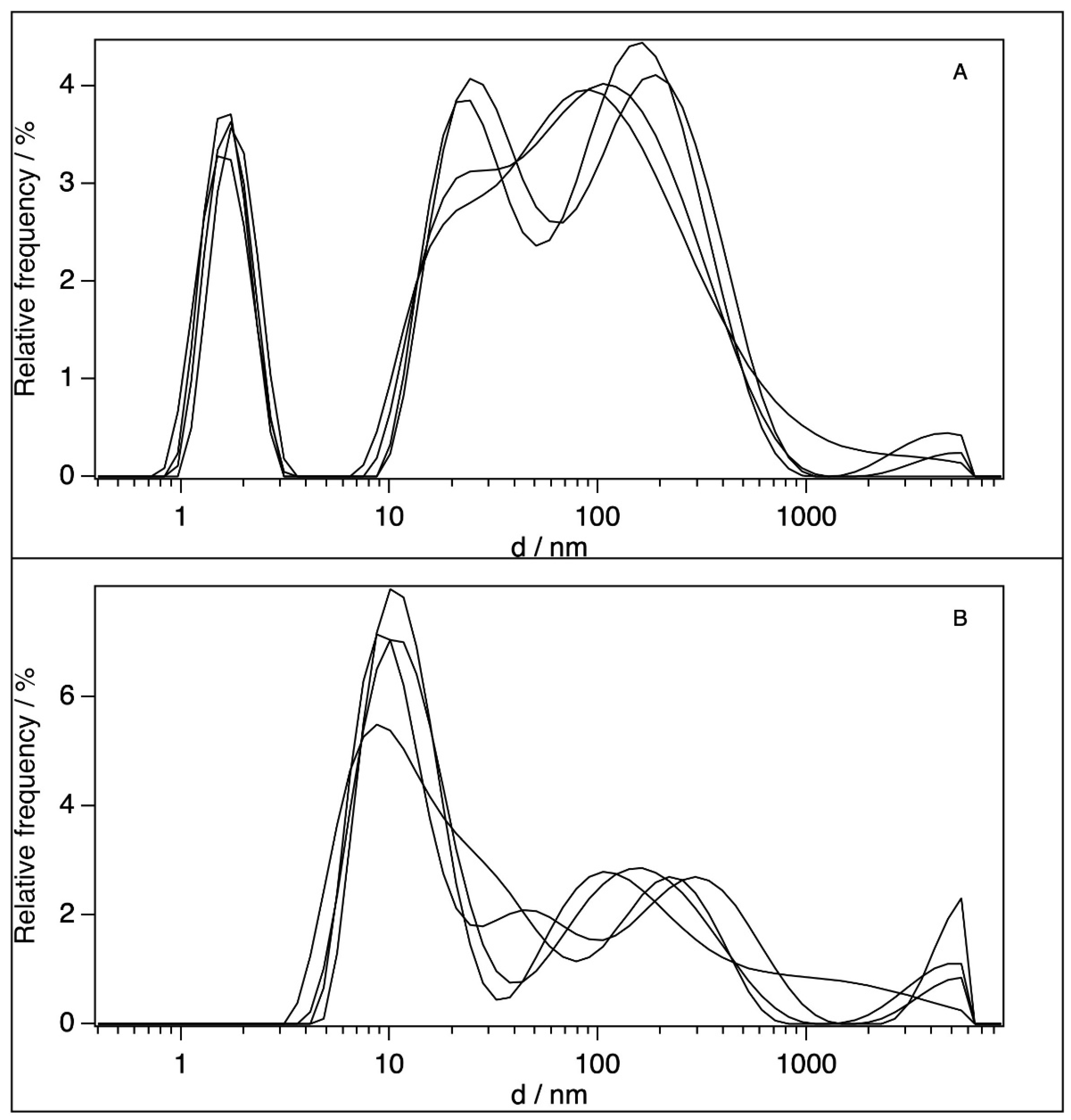
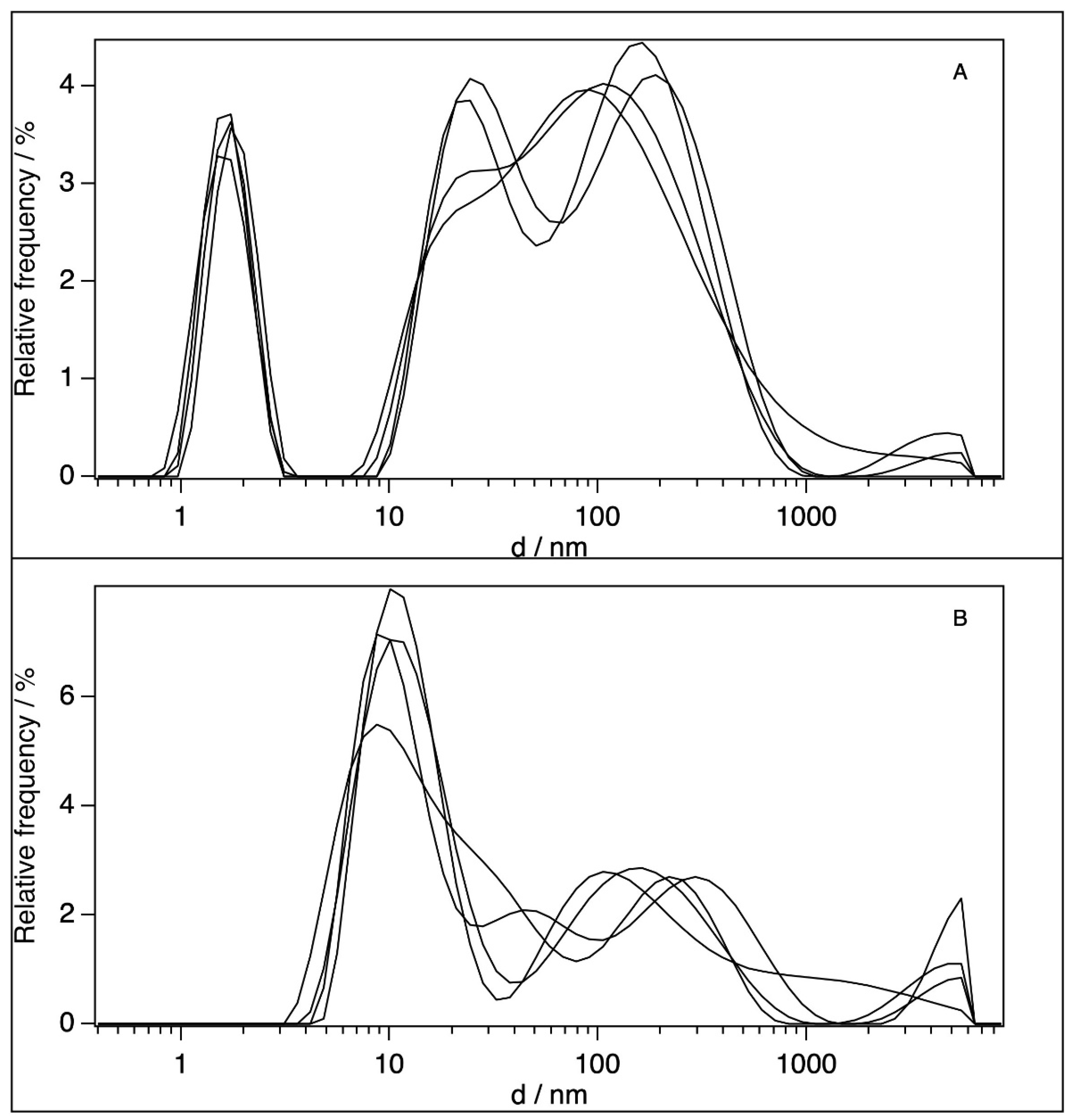
2.2. Dynamic Light Scattering (DLS)
2.3. Samples for Fluorescence and EPR Spectroscopy
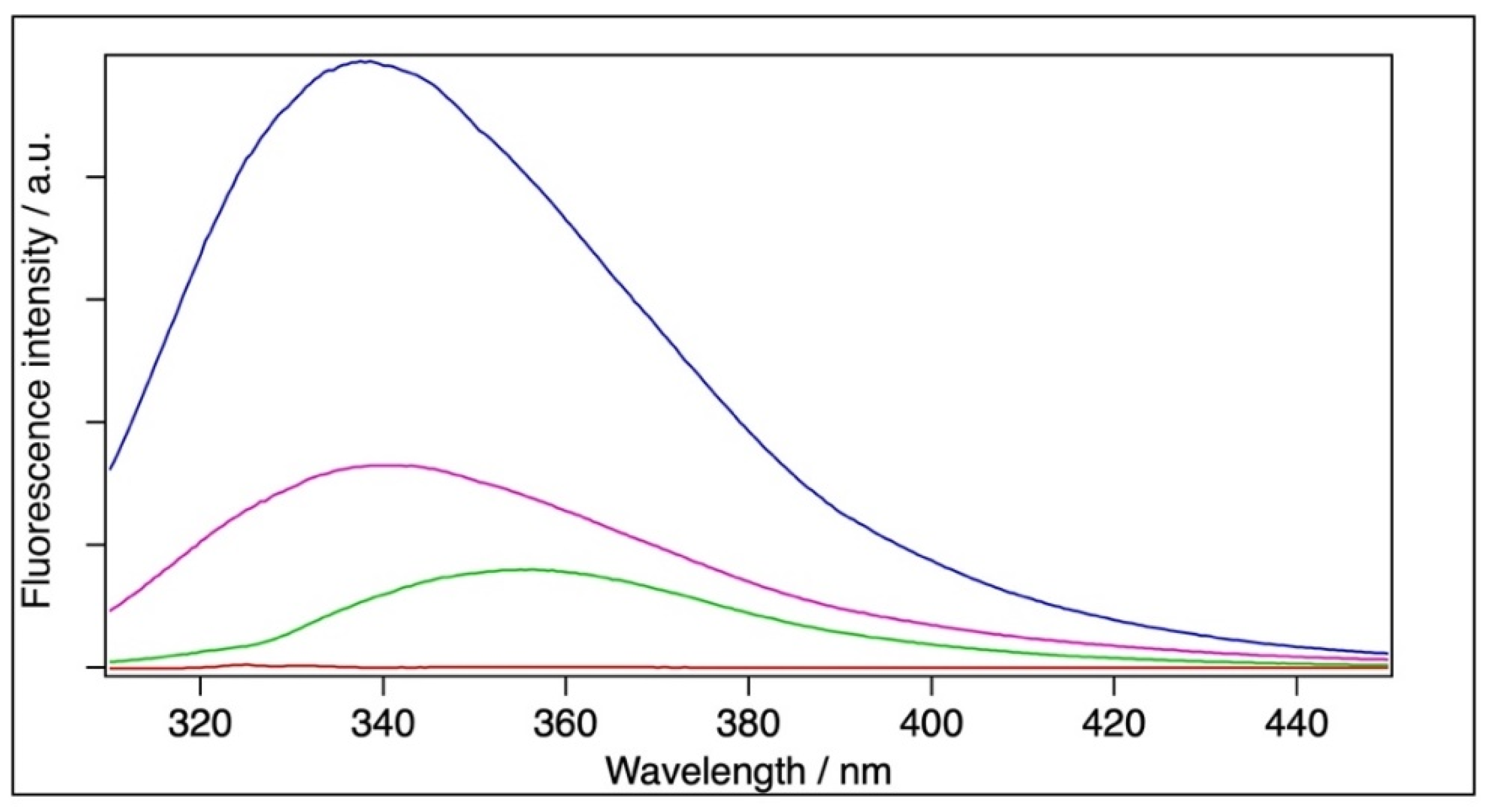
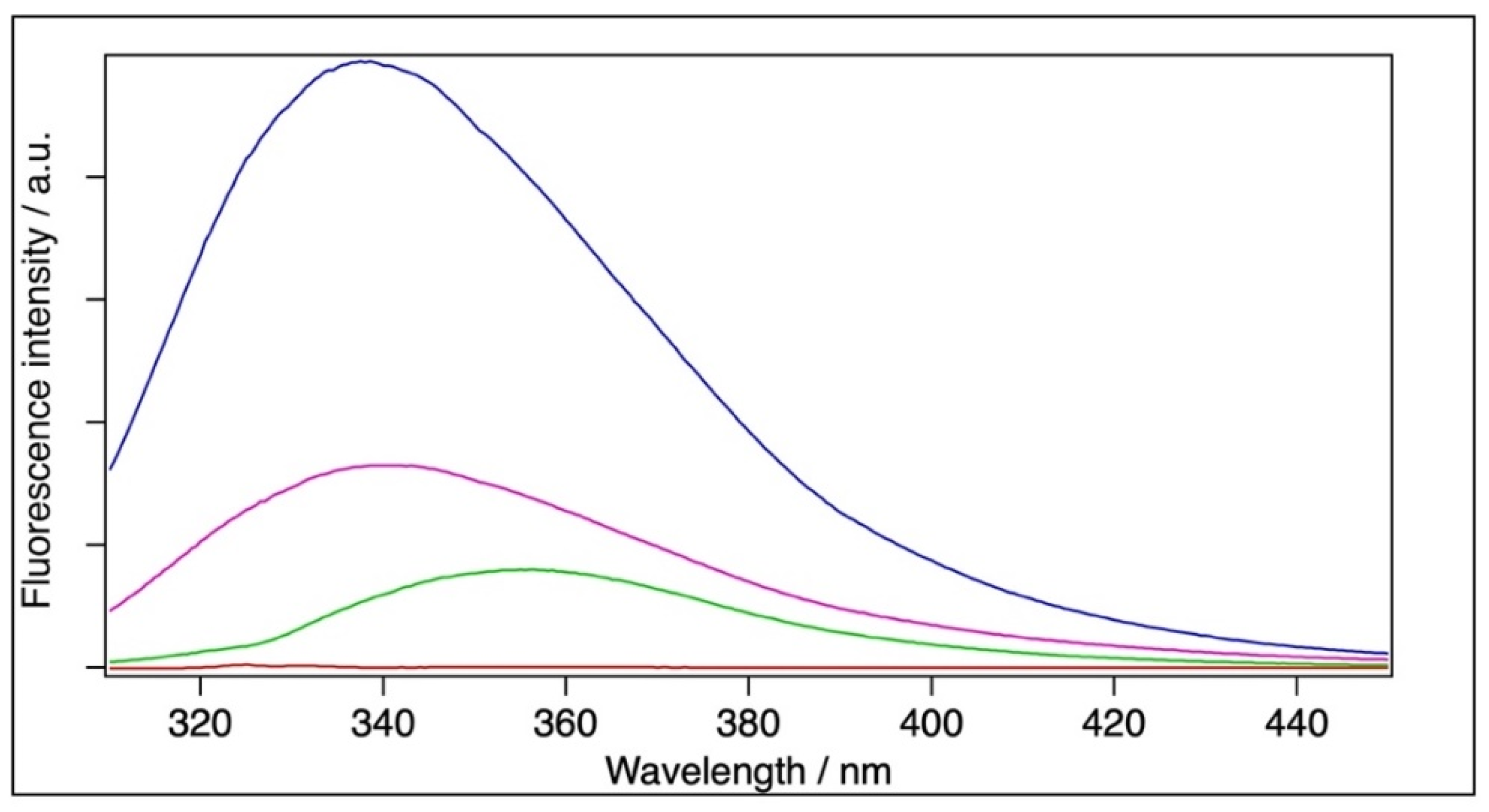
2.4. Fluorescence Spectroscopy
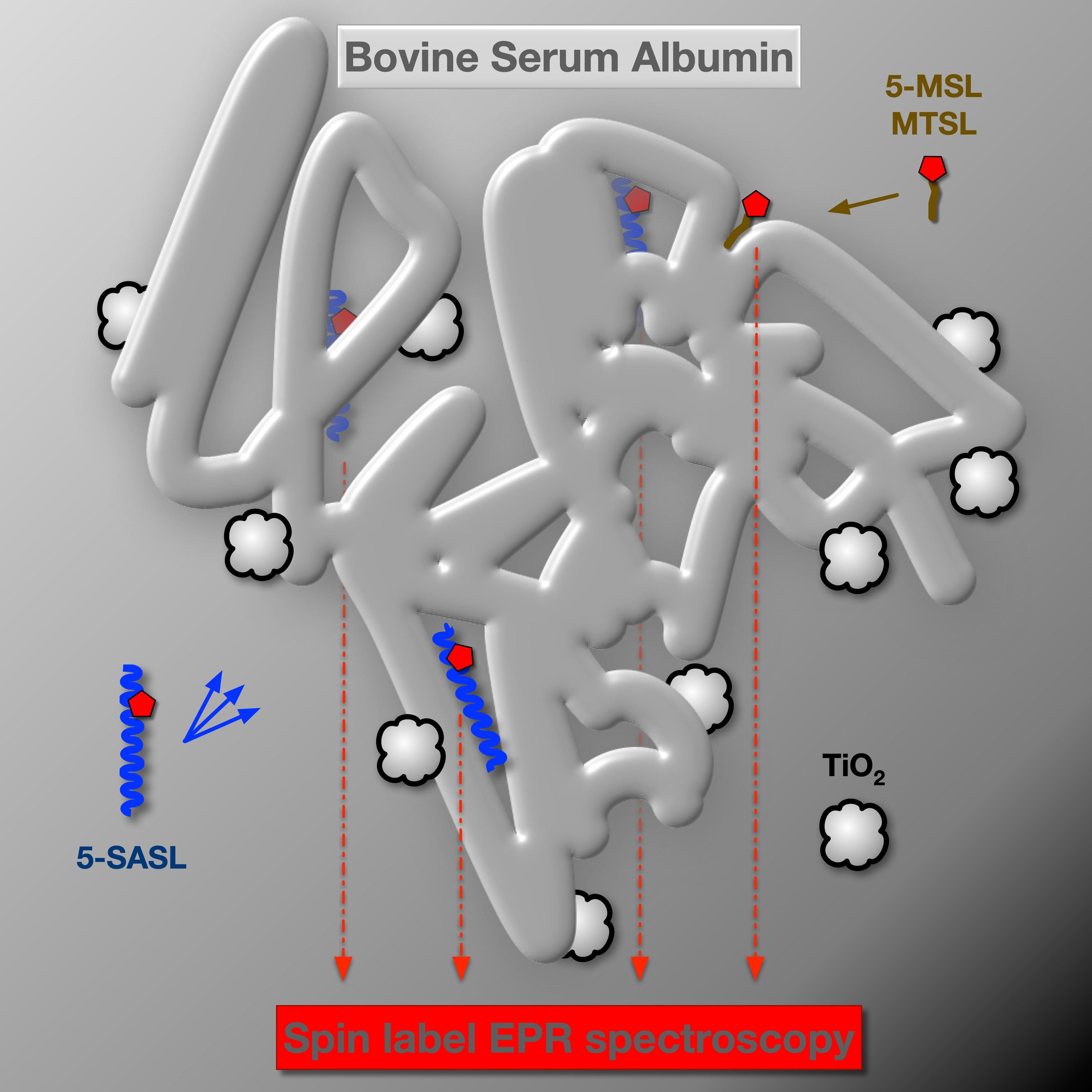
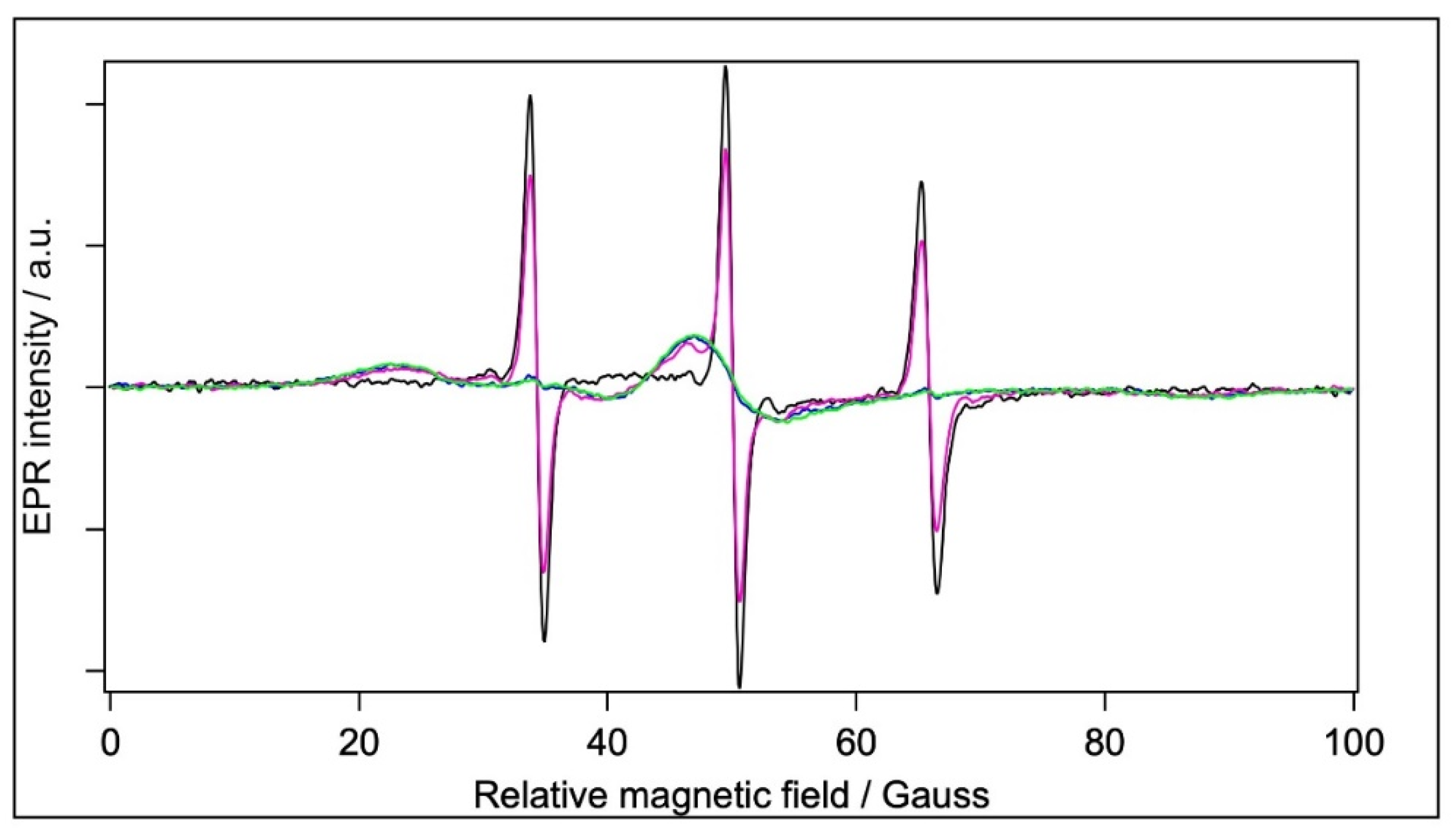
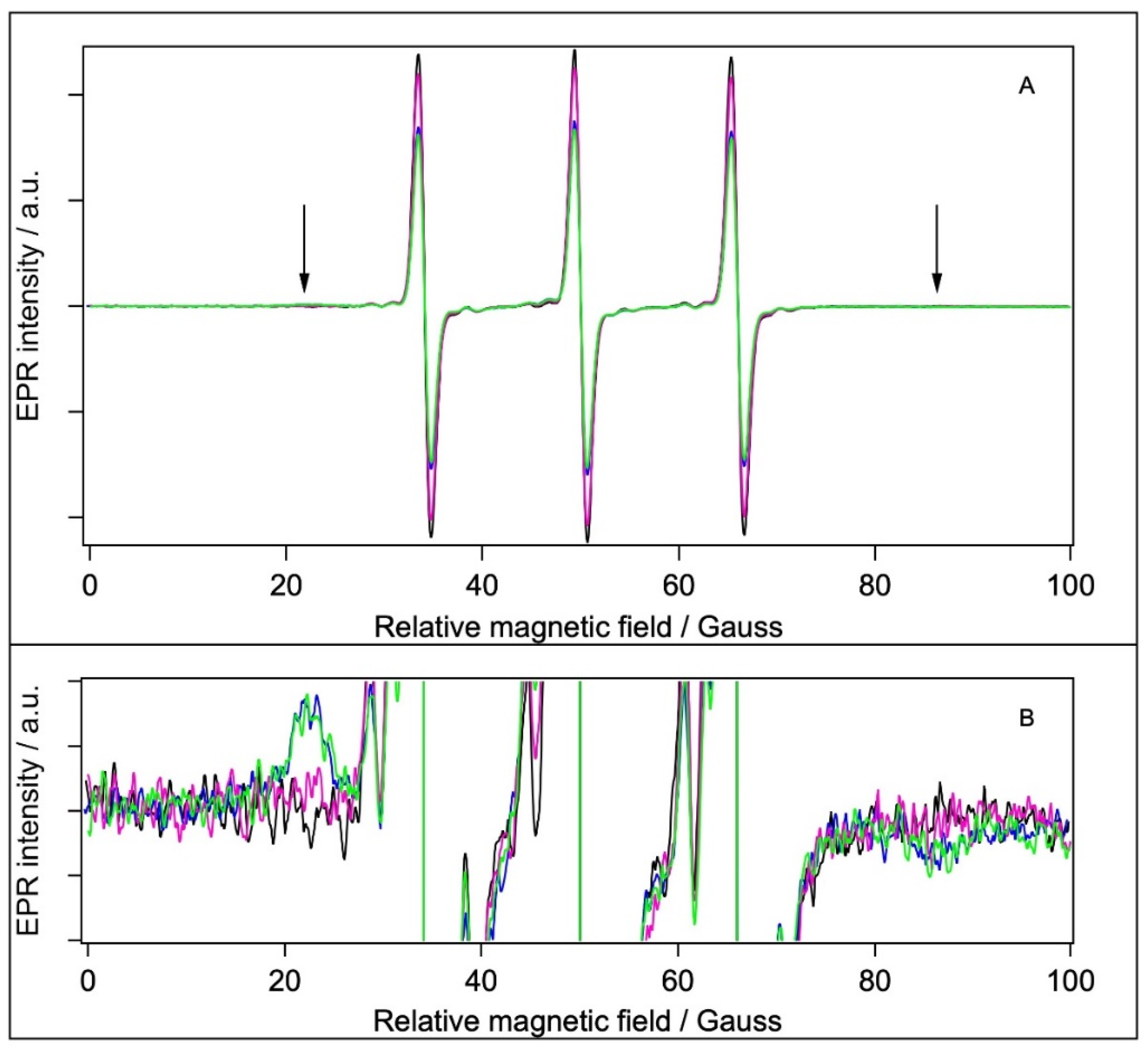
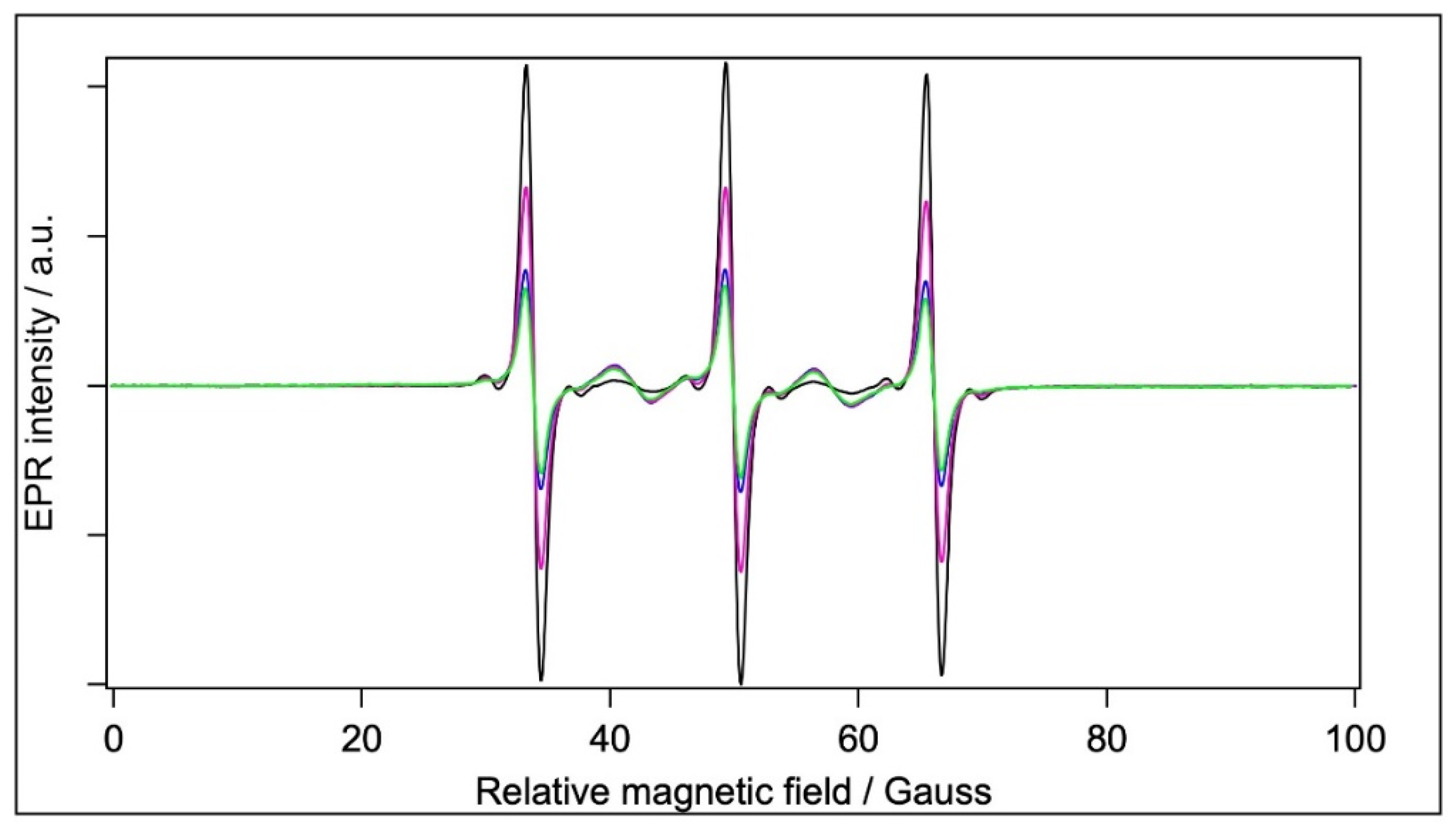
2.5. EPR Spectroscopy
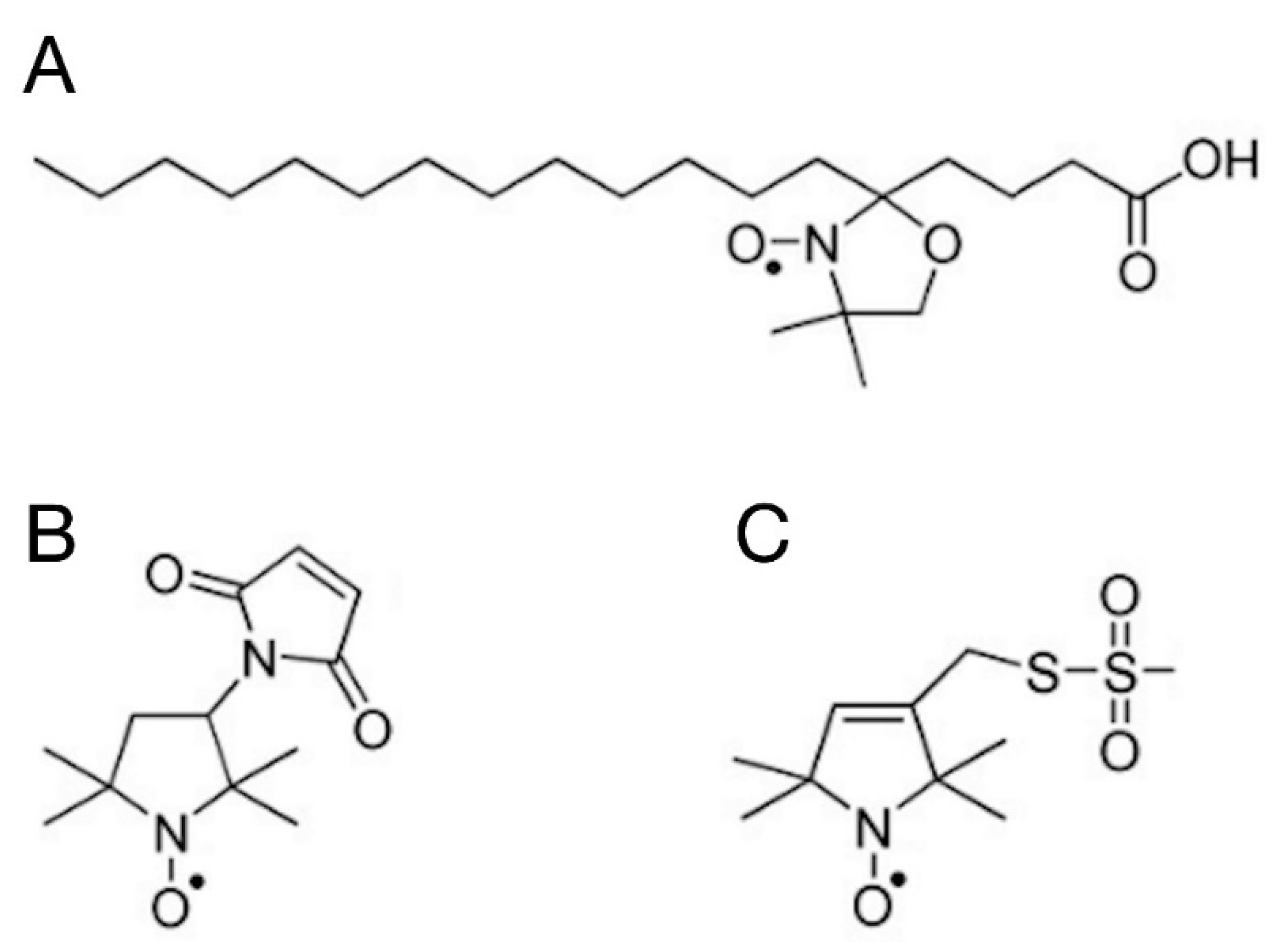
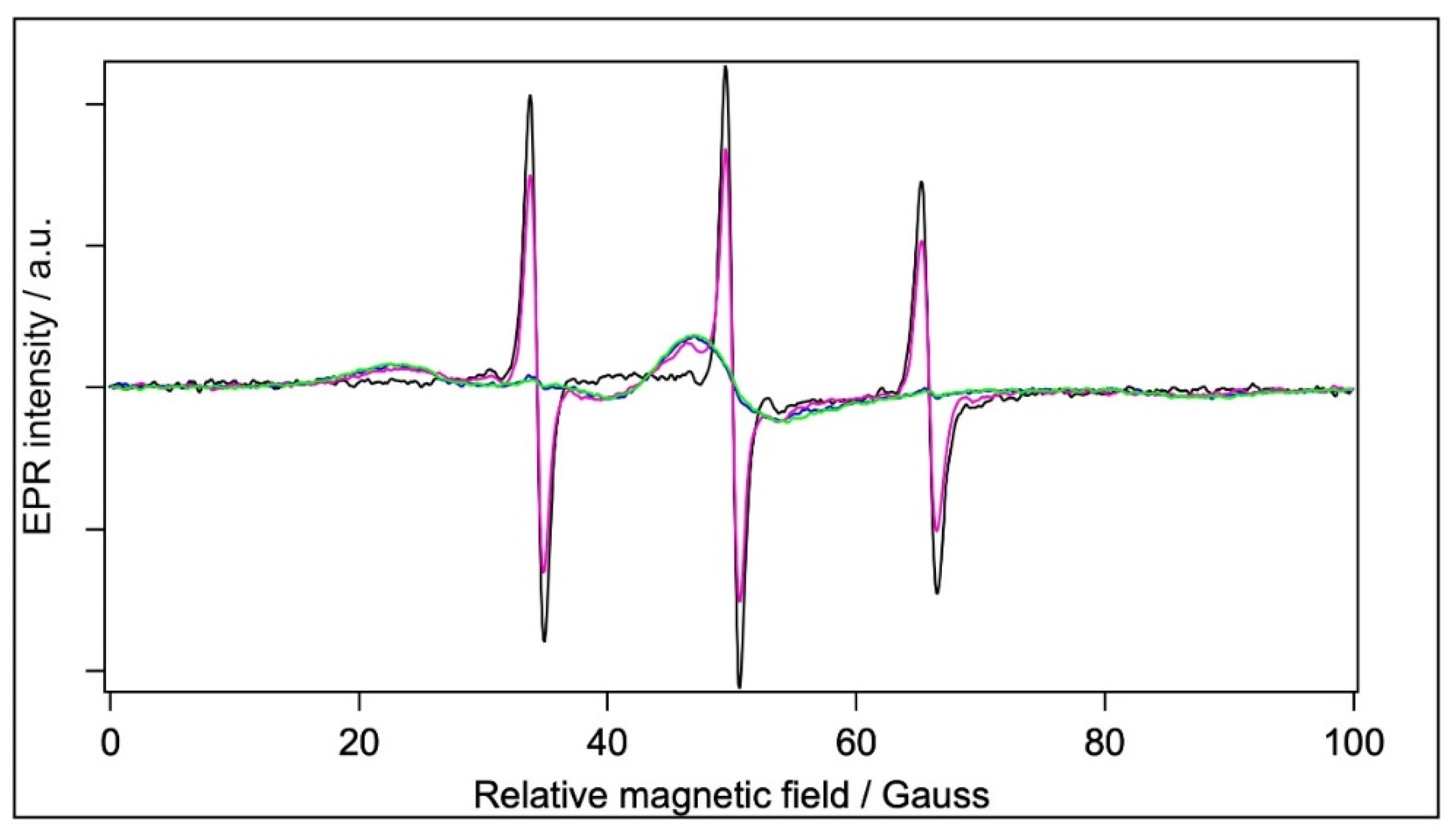
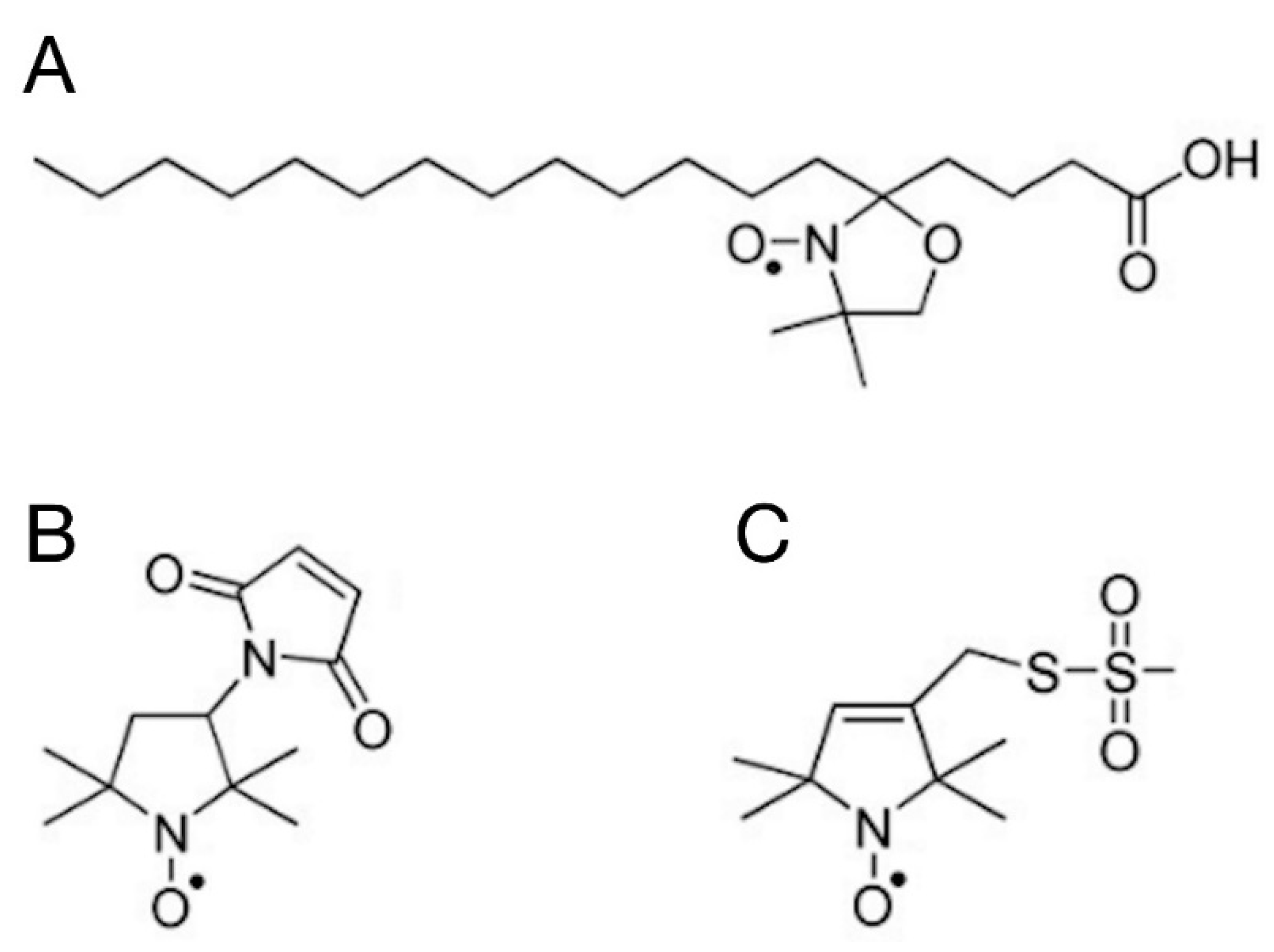
2.5.1. Spin Labelling with 5-SASL
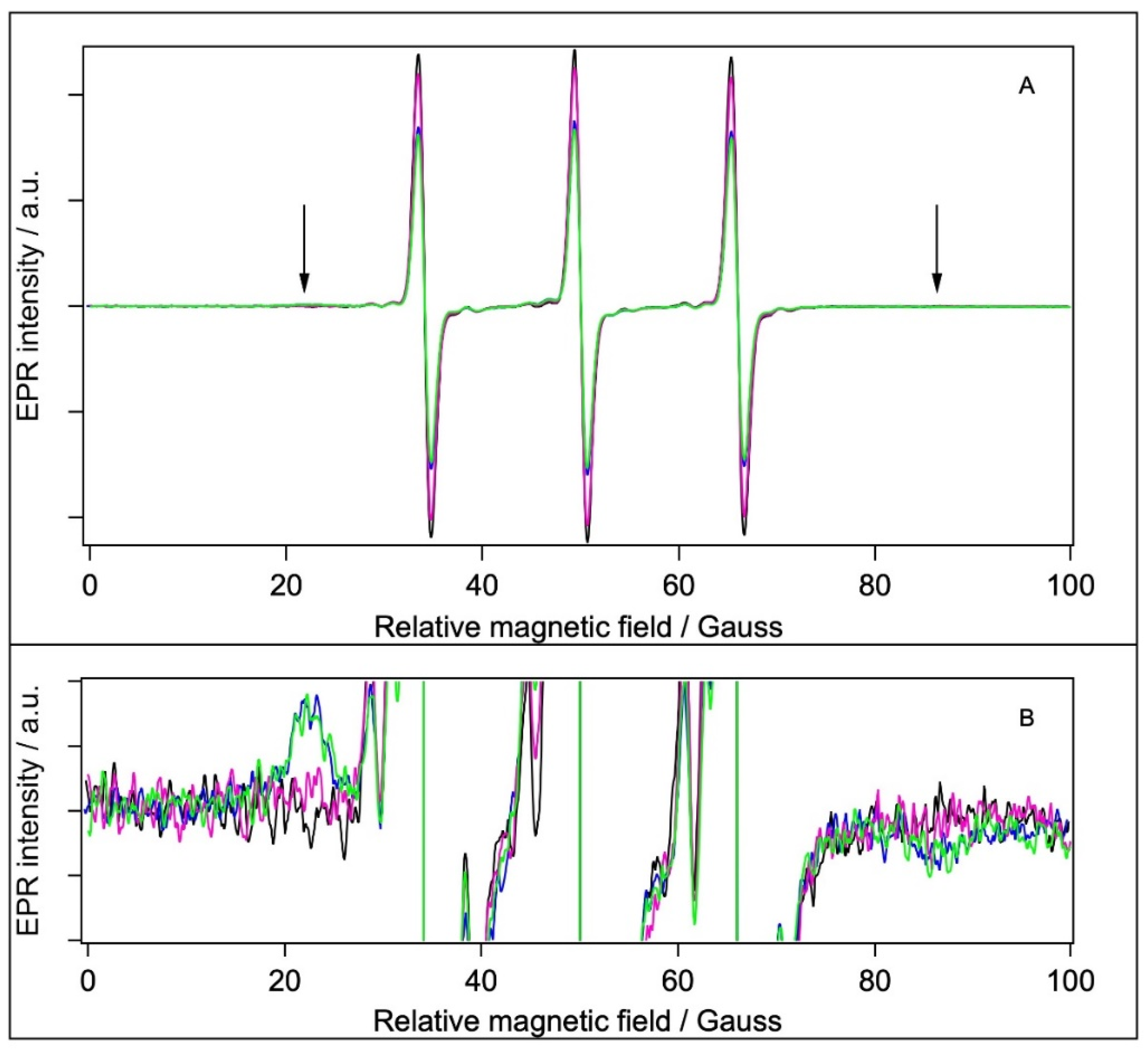
2.5.2. Spin Labelling with 5-MSL
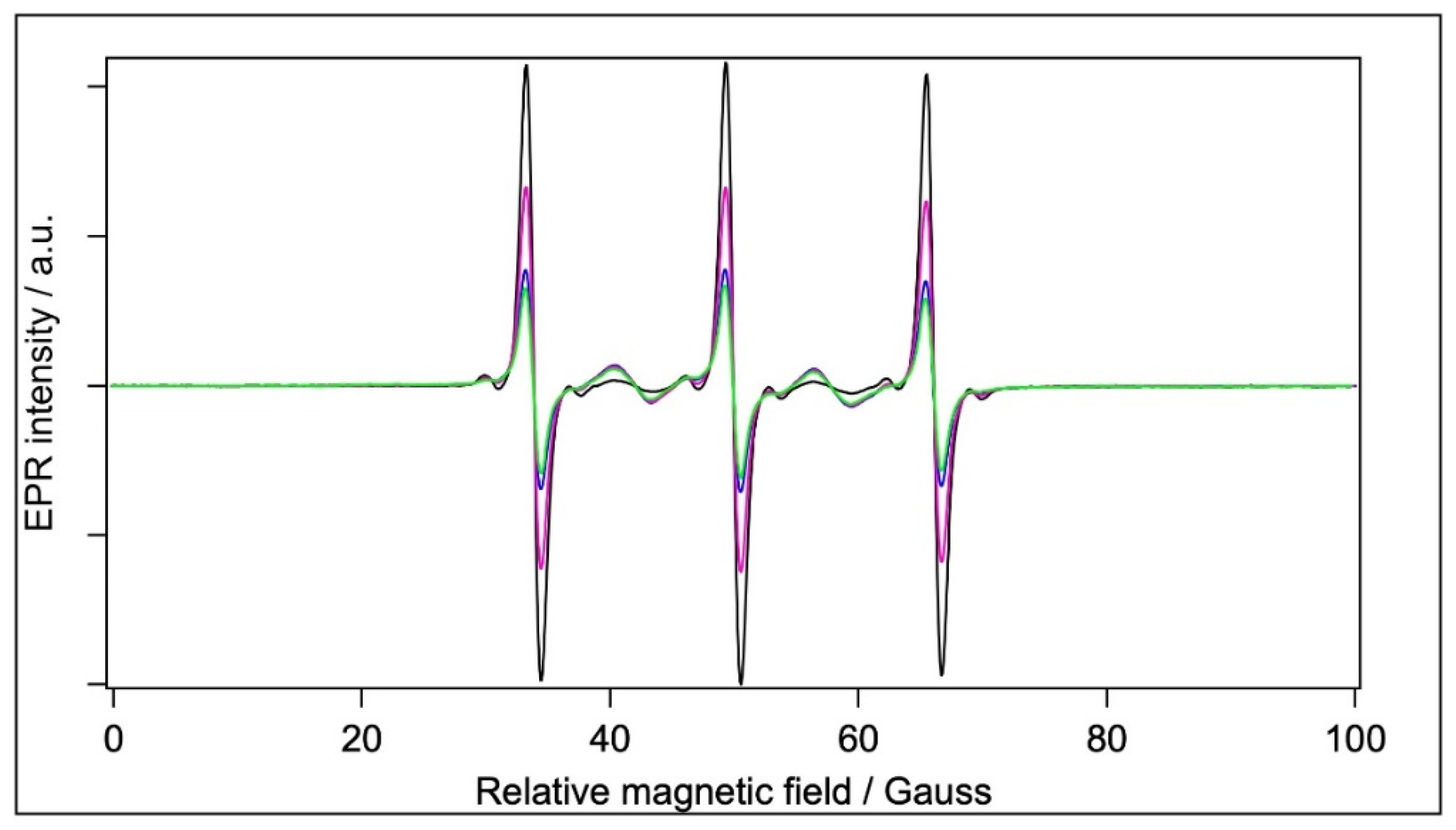
2.5.3. Spin Labelling with MTSL
3. Materials and Methods
3.1. Chemicals and Sample Preparation
3.2. Experimental Procedures and Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wangoo, N.; Suri, C.R.; Shekhawat, G. Interaction of Gold Nanoparticles with Protein: A Spectroscopic Study to Monitor Protein Conformational Changes. Appl. Phys. Lett. 2008, 92, 133104. [Google Scholar] [CrossRef]
- Gheshlaghi, Z.N.; Riazi, G.H.; Ahmadian, S.; Ghafari, M.; Mahinpour, R. Toxicity and Interaction of Titanium Dioxide Nanoparticles with Microtubule Protein. Acta Biochim. Biophys. Sin. 2008, 40, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, M.; Mandal, G.; Ganguly, T. Steady State, Time Resolved, and Circular Dichroism Spectroscopic Studies to Reveal the Nature of Interactions of Zinc Oxide Nanoparticles with Transport Protein Bovine Serum Albumin and to Monitor the Possible Protein Conformational Changes. J. Appl. Phys. 2009, 106, 034701. [Google Scholar] [CrossRef]
- Jigar, E.; Bagi, K.; Fazekas, Á.; Kertész, S.; Veréb, G.; László, Z. Filtration of BSA through TiO2 Photocatalyst Modified PVDF Membranes. Desalin. Water Treat. 2020, 192, 392–399. [Google Scholar] [CrossRef]
- Sisay, E.J.; Fazekas, Á.F.; Gyulavári, T.; Kopniczky, J.; Hopp, B.; Veréb, G.; László, Z. Investigation of Photocatalytic PVDF Membranes Containing Inorganic Nanoparticles for Model Dairy Wastewater Treatment. Membranes 2023, 13, 656. [Google Scholar] [CrossRef]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of Nanoparticles with Proteins: Relation to Bio-Reactivity of the Nanoparticle. J. Nanobiotechnol. 2013, 11, 26. [Google Scholar] [CrossRef]
- Cao, X.; Ma, J.; Shi, X.; Ren, Z. Effect of TiO2 Nanoparticle Size on the Performance of PVDF Membrane. Appl. Surf. Sci. 2006, 253, 2003–2010. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Y.; Hong, Q.; Rao, C.; Zhang, H.; Dong, Y.; Huang, L.; Lu, X.; Bao, N. Bovine Serum Albumin Adsorption in Mesoporous Titanium Dioxide: Pore Size and Pore Chemistry Effect. Langmuir 2016, 32, 3995–4003. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, M.; Meng, X.; Wang, L.; Huang, D. Effect of Protein on PVDF Ultrafiltration Membrane Fouling Behavior under Different pH Conditions: Interface Adhesion Force and XDLVO Theory Analysis. Front. Environ. Sci. Eng. 2016, 10, 12. [Google Scholar] [CrossRef]
- Marquez, A.; Berger, T.; Feinle, A.; Hüsing, N.; Himly, M.; Duschl, A.; Diwald, O. Bovine Serum Albumin Adsorption on TiO2 Colloids: The Effect of Particle Agglomeration and Surface Composition. Langmuir 2017, 33, 2551–2558. [Google Scholar] [CrossRef]
- Shen, L.; Feng, S.; Li, J.; Chen, J.; Li, F.; Lin, H.; Yu, G. Surface Modification of Polyvinylidene Fluoride (PVDF) Membrane via Radiation Grafting: Novel Mechanisms Underlying the Interesting Enhanced Membrane Performance. Sci. Rep. 2017, 7, 2721. [Google Scholar] [CrossRef] [PubMed]
- Nascimben Santos, E.; Fazekas, Á.; Hodúr, C.; László, Z.; Beszédes, S.; Scheres Firak, D.; Gyulavári, T.; Hernádi, K.; Arthanareeswaran, G.; Veréb, G. Statistical Analysis of Synthesis Parameters to Fabricate PVDF/PVP/TiO2 Membranes via Phase-Inversion with Enhanced Filtration Performance and Photocatalytic Properties. Polymers 2021, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Squire, P.G.; Moser, P.; O’Konski, C.T. The Hydrodynamic Properties of Bovine Serum Albumin Monomer and Dimer. Biochemistry 1968, 7, 4261–4272. [Google Scholar] [CrossRef] [PubMed]
- Brahma, A.; Mandal, C.; Bhattacharyya, D. Characterization of a Dimeric Unfolding Intermediate of Bovine Serum Albumin under Mildly Acidic Condition. Biochim. Biophys. Acta 2005, 1751, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, M.A. Study of Bovine Serum Albumin Solubility in Aqueous Solutions by Intrinsic Viscosity Measurements. Adv. Phys. Chem. 2013, 2013, 360239. [Google Scholar] [CrossRef]
- Devi, L.B.; Mandal, A.B. BSA can Form Micelle in Aqueous Solution. J. Surf. Sci. Technol. 2015, 31, 21–29. [Google Scholar]
- Li, R.; Wu, Z.; Wangb, Y.; Ding, L.; Wang, Y. Role of pH-Induced Structural Change in Protein Aggregation in Foam Fractionation of Bovine Serum Albumin. Biotechnol. Rep. (Amst.) 2016, 9, 46–52. [Google Scholar] [CrossRef]
- Varga, N.; Hornok, V.; Sebők, D.; Dékány, I. Comprehensive Study on the Structure of the BSA from Extended-to Aged Form in Wide (2-12) pH Range. Int. J. Biol. Macromol. 2016, 88, 51–58. [Google Scholar] [CrossRef]
- de Sousa Neto, D.; Salmon, C.E.; Alonso, A.; Tabak, M. Interaction of Bovine Serum Albumin (BSA) with Ionic Surfactants Evaluated by Electron Paramagnetic Resonance (EPR) Spectroscopy. Colloids Surf. B Biointerfaces 2009, 70, 147–156. [Google Scholar] [CrossRef]
- Neacsu, M.-V.; Matei, I.; Ionita, G. The Extent of Albumin Denaturation Induced by Aliphatic Alcohols: An Epr and Circular Dichroism Study. Rev. Roum. Chim. 2017, 62, 637–643. [Google Scholar]
- Pavićević, A.; Luo, J.; Popović-Bijelić, A.; Mojović, M. Maleimido-proxyl as an EPR Spin Label for the Evaluation of Conformational Changes of Albumin. Eur. Biophys. J. 2017, 46, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Reichenwallner, J.; Oehmichen, M.-T.; Schmelzer, C.; Hauenschild, T.; Kerth, A.; Hinderberger, D. Exploring the pH-Induced Functional Phase Space of Human Serum Albumin by EPR Spectroscopy. Magnetochemistry 2018, 4, 47. [Google Scholar] [CrossRef]
- Spector, A.A.; John, K.; Fletcher, J.E. Binding of Long-chain Fatty Acids to Bovine Serum Albumin. J. Lipid Res. 1969, 10, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Simard, J.R.; Zunszain, P.A.; Hamilton, J.A.; Curry, S. Location of High and Low Affinity Fatty Acid Binding Sites on Human Serum Albumin Revealed by NMR Drug-competition Analysis. J. Mol. Biol. 2006, 361, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Fanali, G.; di Masi, A.; Trezza, V.; Marino, M.; Fasano, M.; Ascenzi, P. Human Serum Albumin: From Bench to Bedside. Mol. Aspects Med. 2012, 33, 209–290. [Google Scholar] [CrossRef] [PubMed]
- Rizzuti, B.; Bartucci, R.; Sportelli, L.; Guzzi, R. Fatty Acid Binding into the Highest Affinity Site of Human Serum Albumin Observed in Molecular Dynamics Simulation. Arch. Biochem. Biophys. 2015, 579, 18–25. [Google Scholar] [CrossRef]
- Ge, M.T.; Rananavare, S.B.; Freed, J.H. ESR Studies of Stearic Acid Binding to Bovine Serum Albumin. Biochim. Biophys. Acta 1990, 1036, 228–236. [Google Scholar] [CrossRef]
- Livshits, V.A.; Marsh, D. Fatty Acid Binding Sites of Serum Albumin Probed by Non-linear Spin-label EPR. Biochim. Biophys. Acta 2000, 1466, 350–360. [Google Scholar] [CrossRef]
- Junk, M.J.; Spiess, H.W.; Hinderberger, D. DEER in Biological Multispin-systems: A Case Study on the Fatty Acid Binding to Human Serum Albumin. J. Magn. Reson. 2011, 210, 210–217. [Google Scholar] [CrossRef]
- Pavićević, A.A.; Popović-Bijelić, A.D.; Mojović, M.D.; Šušnjar, S.V.; Bačić, G.G. Binding of Doxyl Stearic Spin Labels to Human Serum Albumin: An EPR Study. J. Phys. Chem. B 2014, 118, 10898–10905. [Google Scholar] [CrossRef]
- Reichenwallner, J.; Hauenschild, T.; Schmelzer, C.E.H.; Hülsmann, M.; Godt, A.; Hinderberger, D. Fatty Acid Triangulation in Albumins Using a Landmark Spin Label. Isr. J. Chem. 2019, 59, 1059–1074. [Google Scholar] [CrossRef]
- Morrisett, J.D.; Pownall, H.J.; Gotto, A.M. Bovine serum albumin. Study of the Fatty Acid and Steroid Binding Sites using Spin-labeled lipids. J. Biol. Chem. 1975, 250, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Perkins, R.C., Jr.; Abumrad, N.; Balasubramanian, K.; Dalton, L.R.; Beth, A.H.; Park, J.H.; Park, C.R. Equilibrium Binding of Spin-labeled Fatty Acids to Bovine Serum Albumin: Suitability as Surrogate Ligands for Natural Fatty Acids. Biochemistry 1982, 21, 4059–4064. [Google Scholar] [CrossRef]
- Benga, G.; Strach, S.J. Interpretation of the Electron Spin Resonance Spectra of Nitroxide-maleimide-labelled Proteins and the Use of this Technique in the Study of Albumin and Biomembranes. Biochim. Biophys. Acta 1975, 400, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D.; Livshits, V.A.; Pali, T.; Gaffney, B.J. Recent Development in Biological Spin-label Spectroscopy. In Spectroscopy of Biological Molecules: New Directions; Greve, J., Puppels, G.J., Otto, C., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1999; pp. 647–650. [Google Scholar] [CrossRef]
- Páli, T.; Marsh, D. Structural Studies on Membrane Proteins Using Non-linear Spin Label EPR Spectroscopy. Cell. Mol. Biol. Lett. 2002, 7, 87–91. [Google Scholar] [PubMed]
- Bordignon, E. EPR Spectroscopy of Nitroxide Spin Probes. eMagRes 2017, 6, 6235–6254. [Google Scholar] [CrossRef]
- Bujacz, A. Structures of Bovine, Equine and Leporine Serum Albumin. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 1278–1289. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Bujacz, A.; Bujacz, G. Crystal Structure of Bovine Serum Albumin. Transp. Protein 2012. [Google Scholar] [CrossRef]
- Celej, M.S.; Montich, G.G.; Fidelio, G.D. Protein Stability Induced by Ligand Binding Correlates with Changes in Protein Flexibility. Protein Sci. 2003, 12, 1496–1506. [Google Scholar] [CrossRef]
- Togashi, D.M.; Ryder, A.G.; Mc Mahon, D.; Dunne, P.; McManus, J. Fluorescence Study of Bovine Serum Albumin and Ti and Sn Oxide Nanoparticles Interactions. In Diagnostic Optical Spectroscopy in Biomedicine IV, Proceedings of the SPIE-OSA Biomedical Optics, Munich, Germany 17–21 June 2007; Schweitzer, D., Fitzmaurice, M., Eds.; Optica Publishing Group: Washington, DC, USA, 2007; Volume 6628, p. 6628_61. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Jain, N.; Bhasne, K.; Kumari, V.; Mukhopadhyay, S. pH-Induced Conformational Isomerization of Bovine Serum Albumin Studied by Extrinsic and Intrinsic Protein Fluorescence. J. Fluoresc. 2011, 21, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.T.; Callis, P.R. Mechanisms of Tryptophan Fluorescence Shifts in Proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Rodrigues, F.H.; Delgado, G.G.; Santana da Costa, T.; Tasic, L. Applications of Fluorescence Spectroscopy in Protein Conformational Changes and Intermolecular Contacts. BBA Adv. 2023, 3, 100091. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, G.; Mei, Z. Spectroscopic and Dynamic Light Scattering Studies of the Interaction between Pterodontic Acid and Bovine Serum Albumin. Acta Pharm. Sin. B 2012, 2, 53–59. [Google Scholar] [CrossRef]
- Polat, H.; Kutluay, G.; Polat, M. Analysis of Dilution Induced Disintegration of Micellar Drug Carriers in the Presence of Inter and Intra Micellar Species. Colloids Surf. A 2020, 601, 124989. [Google Scholar] [CrossRef]
- Burgstaller, C.; Etchart, N.N. Dimerization of Bovine Serum Albumin as Evidenced by Particle Size and Molecular Mass Measurement. Anton Paar GmbH. D51|A044EN-A. 2018. Available online: https://www.anton-paar.com/corp-en/services-support/document-finder/application-reports/dimerization-of-bovine-serum-albumin-as-evidenced-by-particle-size-and-molecular-mass-measurement/ (accessed on 31 August 2023).
- Howe, K.J.; Clark, M.M. Fouling of Microfiltration and Ultrafiltration Membranes by Natural Waters. Environ. Sci. Technol. 2002, 36, 3571–3576. [Google Scholar] [CrossRef]
- Bacova, J.; Knotek, P.; Kopecka, K.; Hromádko, L.; Čapek, J.; Nývltová, P.; Bruckova, L.; Schroterova, L.; Sestakova, B.; Palarcik, J.; et al. Evaluating the Use of TiO2 Nanoparticles for Toxicity Testing in Pulmonary A549 Cells. Int. J. Nanomed. 2022, 17, 4211–4225. [Google Scholar] [CrossRef]
- Teale, F.W.J.; Weber, G. Ultraviolet Fluorescence of the Aromatic Amino Acids. Biochem. J. 1957, 65, 476–482. [Google Scholar] [CrossRef]
- Pocanschi, C.L.; Popot, J.L.; Kleinschmidt, J.H. Folding and Stability of Outer Membrane Protein A (OmpA) from Escherichia Coli in an Amphipathic Polymer, Amphipol A8-35. Eur. Biophys. J. 2013, 42, 103–118. [Google Scholar] [CrossRef]
- Koch, D.; Manzhos, S. On the Charge State of Titanium in Titanium Dioxide. J. Phys. Chem. Lett. 2017, 8, 1593–1598. [Google Scholar] [CrossRef]
- Pandit, S.; Kundu, S. Fluorescence quenching and related interactions among globular proteins (BSA and lysozyme) in presence of titanium dioxide nanoparticles. Colloids Surf. 2021, 628, 127253. [Google Scholar] [CrossRef]
- Freed, J.H. Theory of Slow Tumbling ESR Spectra of Nitroxides. In Spin Labeling. Theory and Applications; Berliner, L.J., Ed.; Academic Press: New York, NY, USA, 1976; pp. 53–132. [Google Scholar]
- Hubbell, W.L.; Lopez, C.J.; Altenbach, C.; Yang, Z. Technological Advances in Site-directed Spin Labeling of Proteins. Curr. Opin. Struct. Biol. 2013, 23, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.J.; Fleissner, M.R.; Brooks, E.K.; Hubbell, W.L. Stationary-phase EPR for Exploring Protein Structure, Conformation, and Dynamics in Spin-labeled Proteins. Biochemistry 2014, 53, 7067–7075. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Spin-label Order Parameter Calibrations for Slow Motion. Appl. Magn. Reson. 2018, 49, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Páli, T.; Kóta, Z. Studying Lipid-Protein Interactions with Electron Paramagnetic Resonance Spectroscopy of Spin-labeled Lipids. Methods Mol. Biol. 2019, 2013, 529–561. [Google Scholar] [CrossRef]
- Kivelson, D. Theory of ESR Linewidths of Free Radicals. J. Chem. Phys. 1960, 33, 1094–1107. [Google Scholar] [CrossRef]
- Páli, T.; Pesti, M. Chapter V: Phase Transition of Membrane Lipids. In Manual on Membrane Lipids; Prasad, R., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1996; pp. 80–111. [Google Scholar] [CrossRef]
- Griffith, O.H.; Jost, P.C. Lipid Spin Labels in Biological Membranes. In Spin Labeling. Theory and Applications; Berliner, L.J., Ed.; Academic Press: New York, NY, USA, 1976; pp. 453–523. [Google Scholar] [CrossRef]
- Peters, T., Jr. Ligand Binding in Albumin. In All about Albumin; Academic Press: Cambridge, MA, USA, 1995; pp. 76–132. [Google Scholar] [CrossRef]
- Hull, H.H.; Chang, R.; Kaplan, L.J. On the Location of the Sulfhydryl Group in Bovine Plasma Albumin. Biochim. Biophys. Acta 1975, 400, 132–136. [Google Scholar] [CrossRef]
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal Structure of Human Serum Albumin at 2.5 A Resolution. Protein Eng. 1999, 12, 439–446. [Google Scholar] [CrossRef]
- Stewart, A.J.; Blindauer, C.A.; Berezenko, S.; Sleep, D.; Tooth, D.; Sadler, P.J. Role of Tyr84 in Controlling the Reactivity of Cys34 of Human Albumin. FEBS J. 2005, 272, 353–362. [Google Scholar] [CrossRef]
- Griffith, O.H.; McConnell, H.M. A Nitroxide-maleimide Spin Label. Proc. Natl. Acad. Sci. USA 1966, 55, 8–11. [Google Scholar] [CrossRef]
- Lange, A.; Marsh, D.; Wassmer, K.H.; Meier, P.; Kothe, G. Electron Spin Resonance Study of Phospholipid Membranes Employing a Comprehensive Line-Shape Model. Biochemistry 1985, 24, 4383–4392. [Google Scholar] [CrossRef] [PubMed]
- Eaton, S.S.; Woodcock, L.B.; Eaton, G.R. Continuous Wave Electron Paramagnetic Resonance of Nitroxide Biradicals in Fluid Solution. Concepts Magn. Reson. Part A 2018, 47A, 21426. [Google Scholar] [CrossRef] [PubMed]
- Mahy, J.G.; Carcel, C.; Man, M.W.C. Evonik P25 photoactivation in the visible range by surface grafting of modified porphyrins for p-nitrophenol elimination in water. AIMS Mater. 2023, 10, 437–452. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Taneva, S.G.; Krumova, S.; Bogár, F.; Kincses, A.; Stoichev, S.; Todinova, S.; Danailova, A.; Horváth, J.; Násztor, Z.; Kelemen, L.; et al. Insights into Graphene Oxide Interaction with Human Serum Albumin in Isolated State and in Blood Plasma. Int. J. Biol. Macromol. 2021, 175, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Vriend, G. YASARA View—Molecular Graphics for All Devices—From Smartphones to Workstations. Bioinform 2014, 30, 2981–2982. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | [BSA]/M | Membrane | R% |
|---|---|---|---|
| BSA | 1.5 × 10−5 | PVDF | 92 |
| BSA | 1.5 × 10−5 | PVDF/TiO2 | 28 |
| BSA(TiO2) | 1.5 × 10−5 | PVDF | 91 |
| BSA(TiO2) | 4.9 × 10−6 | PVDF | 37 |
| BSA | 4.9 × 10−6 | PVDF | 37 |
| [BSA]/10−6 M | d1/nm | d2/nm | Particle Type | Medium | Reference |
|---|---|---|---|---|---|
| 0.6 | 11 | dimers (d2) | H2O | own result | |
| 0.65 | 8.9 | compact aggregates (d2) | pH 7.2, 10 mM phosphate buffer | [16] | |
| 4.5 | 10 | monomer (d1) | pH 7.0, H2O | [17] | |
| 5 | 7.3 | 13.5 | monomer (d1), dimer (d2) | pH 7.4, H2O | [46] |
| 10 | 3.4 | monomer (d1) | pH 7.0, H2O | [18] | |
| 25 | 10.6 | compact aggregates (d2) | pH 7.2, 10 mM phosphate buffer | [16] | |
| 60 | 1.7 | 24–200 | unfolded monomers (d1) large aggregates (d2) | H2O | own result |
| 100 | 5.2 | undefined particle (d1) | pH 7.4, H2O | [47] | |
| 120.4 | 2 | 22 | denaturated monomers (d1), aggregation of denaturated monomers (d2) | H2O | [48] |
| 120.4 | 12.4 | dimer (d2) | pH 7.4, 0.01 M PBS buffer | [48] |
| Sample | [BSA]/M | Fitted Emission Maximum (λmax)/nm | Mean Emission Maximum (<λF>)/nm |
|---|---|---|---|
| BSA | 6 × 10−5 | 338.9 | 352.3 |
| Filtrated BSA | 1.8 × 10−5 | 340.2 | 356.9 |
| BSA+TiO2 | 6 × 10−5 | - | - |
| BSA | 2 × 10−6 | 338.4 | 352.3 |
| Filtrated BSA | 6 × 10−7 | 340.7 | 355.0 |
| BSA+TiO2 | 2 × 10−6 | 355.6 | 363.3 |
| Sample | Mobile Component/% | Immobile Component/% | τ/ns | 2Azz/G |
|---|---|---|---|---|
| Water | 100 | - | 0.1206 | - |
| BSA | 2.4 | 97.6 | - | 64.62 |
| Filtrated BSA | 71 | 29 | 0.1312 | 62.3–64.1 |
| BSA+TiO2 | 1.0 | 99.0 | - | 65.05 |
| Sample | Mobile Component/% | Immobile Component/% | τ/ns | 2Azz/G |
|---|---|---|---|---|
| Water | 100 | - | 0.0043 | - |
| BSA | 71 | 29 | 0.0082 | 63.92 |
| Filtrated BSA | 93 | 7 | 0.0056 | cc. 61 G * |
| BSA+TiO2 | 67 | 33 | 0.0084 | 64.20 |
| Sample | Mobile Component/% | Immobile Component/% | τ/ns | 2Azz/G |
|---|---|---|---|---|
| Water | 100 | - | 0.0094 | - |
| BSA | 97–98 | 2–3 | 0.0303 | - |
| Filtrated BSA | 100 | - | 0.0224 | - |
| BSA+TiO2 | 97–98 | 2–3 | 0.0536 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sebők-Nagy, K.; Kóta, Z.; Kincses, A.; Fazekas, Á.F.; Dér, A.; László, Z.; Páli, T. Spin-Label Electron Paramagnetic Resonance Spectroscopy Reveals Effects of Wastewater Filter Membrane Coated with Titanium Dioxide Nanoparticles on Bovine Serum Albumin. Molecules 2023, 28, 6750. https://doi.org/10.3390/molecules28196750
Sebők-Nagy K, Kóta Z, Kincses A, Fazekas ÁF, Dér A, László Z, Páli T. Spin-Label Electron Paramagnetic Resonance Spectroscopy Reveals Effects of Wastewater Filter Membrane Coated with Titanium Dioxide Nanoparticles on Bovine Serum Albumin. Molecules. 2023; 28(19):6750. https://doi.org/10.3390/molecules28196750
Chicago/Turabian StyleSebők-Nagy, Krisztina, Zoltán Kóta, András Kincses, Ákos Ferenc Fazekas, András Dér, Zsuzsanna László, and Tibor Páli. 2023. "Spin-Label Electron Paramagnetic Resonance Spectroscopy Reveals Effects of Wastewater Filter Membrane Coated with Titanium Dioxide Nanoparticles on Bovine Serum Albumin" Molecules 28, no. 19: 6750. https://doi.org/10.3390/molecules28196750
APA StyleSebők-Nagy, K., Kóta, Z., Kincses, A., Fazekas, Á. F., Dér, A., László, Z., & Páli, T. (2023). Spin-Label Electron Paramagnetic Resonance Spectroscopy Reveals Effects of Wastewater Filter Membrane Coated with Titanium Dioxide Nanoparticles on Bovine Serum Albumin. Molecules, 28(19), 6750. https://doi.org/10.3390/molecules28196750