
New Biotinylated GHK and Related Copper(II) Complex: Antioxidant and Antiglycant Properties In Vitro against Neurodegenerative Disorders

Abstract
:1. Introduction
2. Results and Discussion
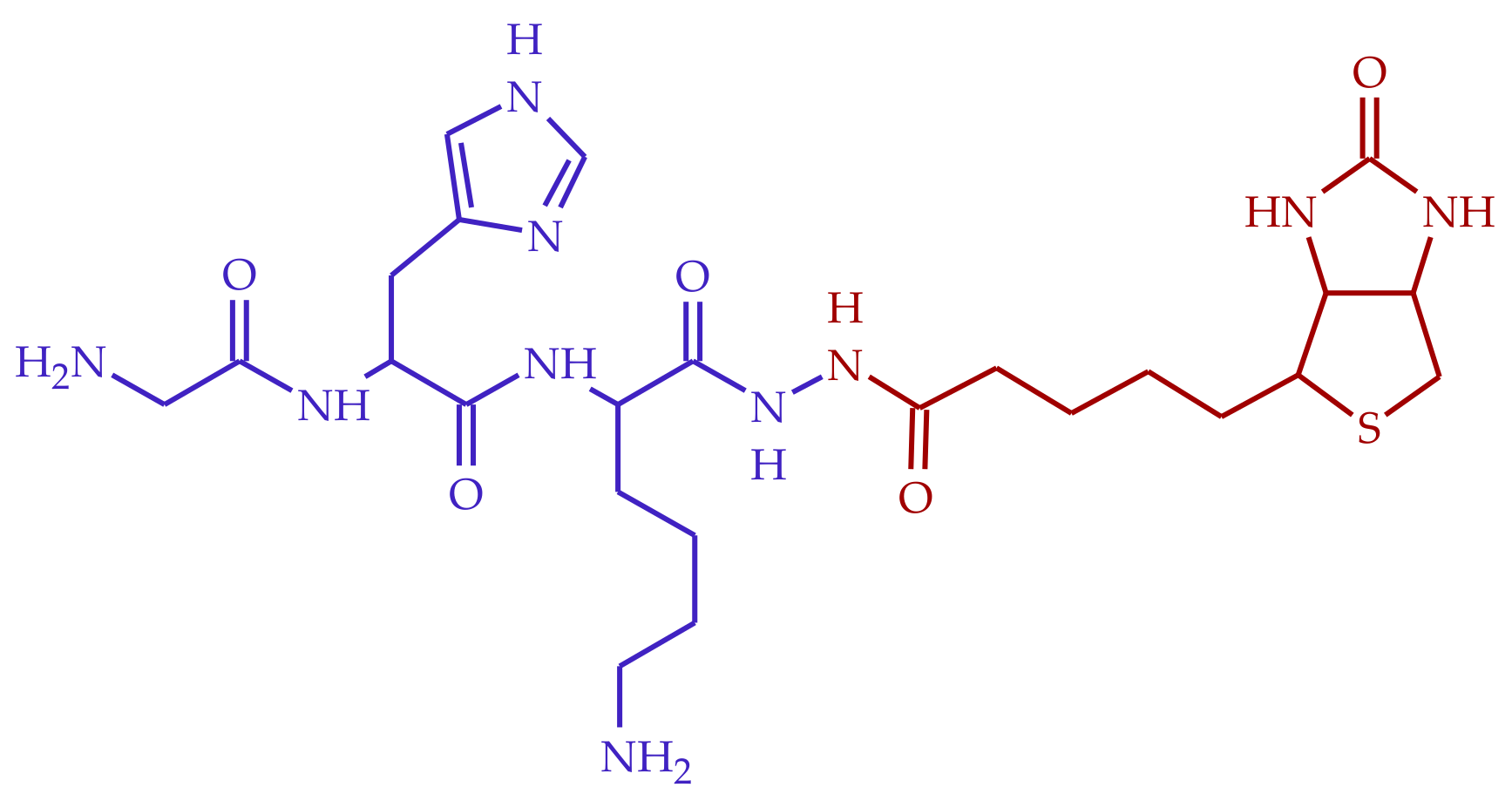
2.1. Synthesis and Purification of BioGHK
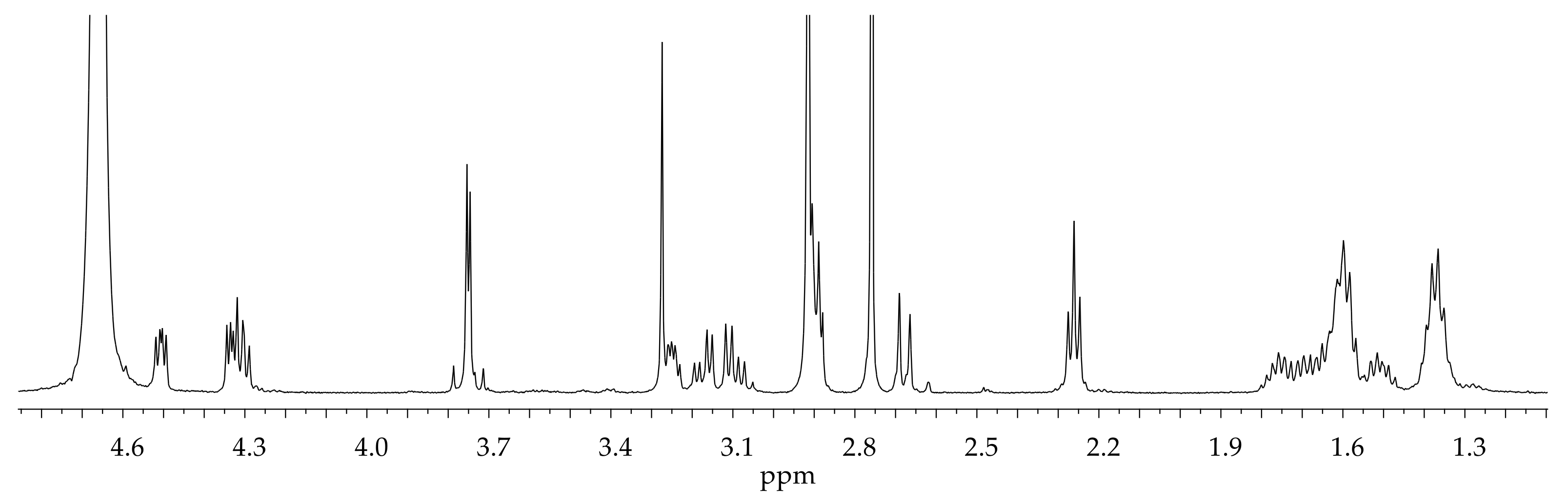
2.2. NMR Characterization of BioGHK
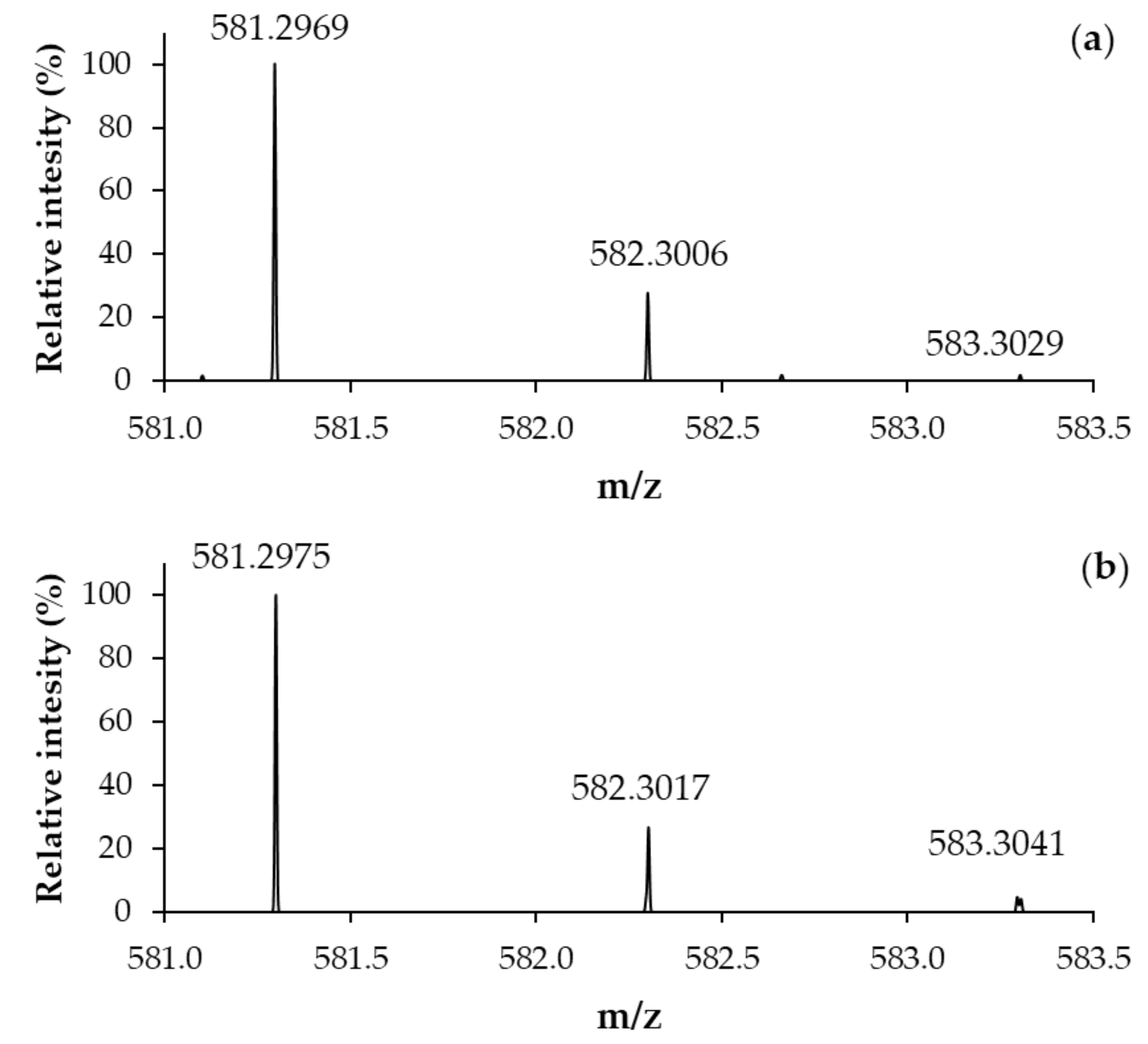
2.3. Structural Characterization of the Ligand
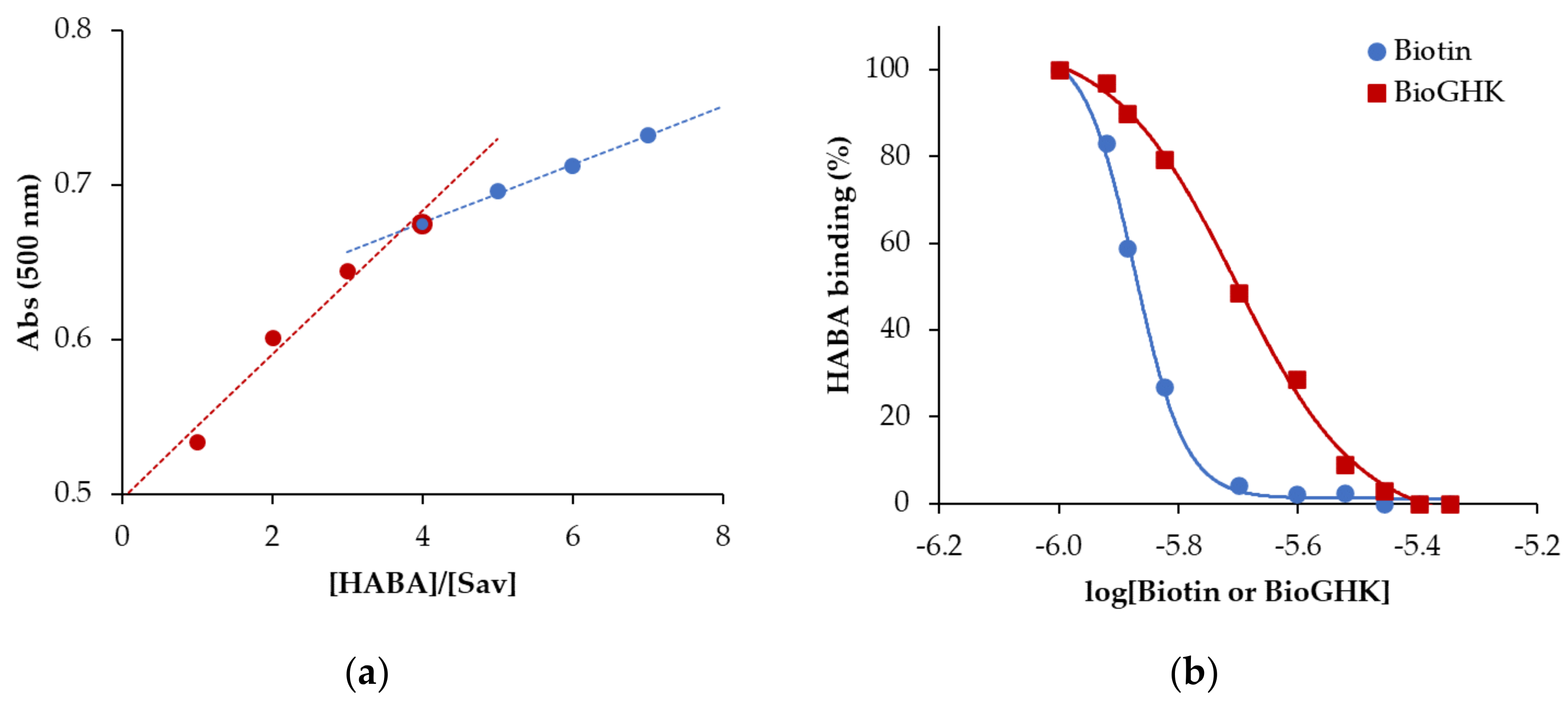
2.4. Interaction Sav/BioGHK
2.5. Characterization of the Cu-BioGHK System
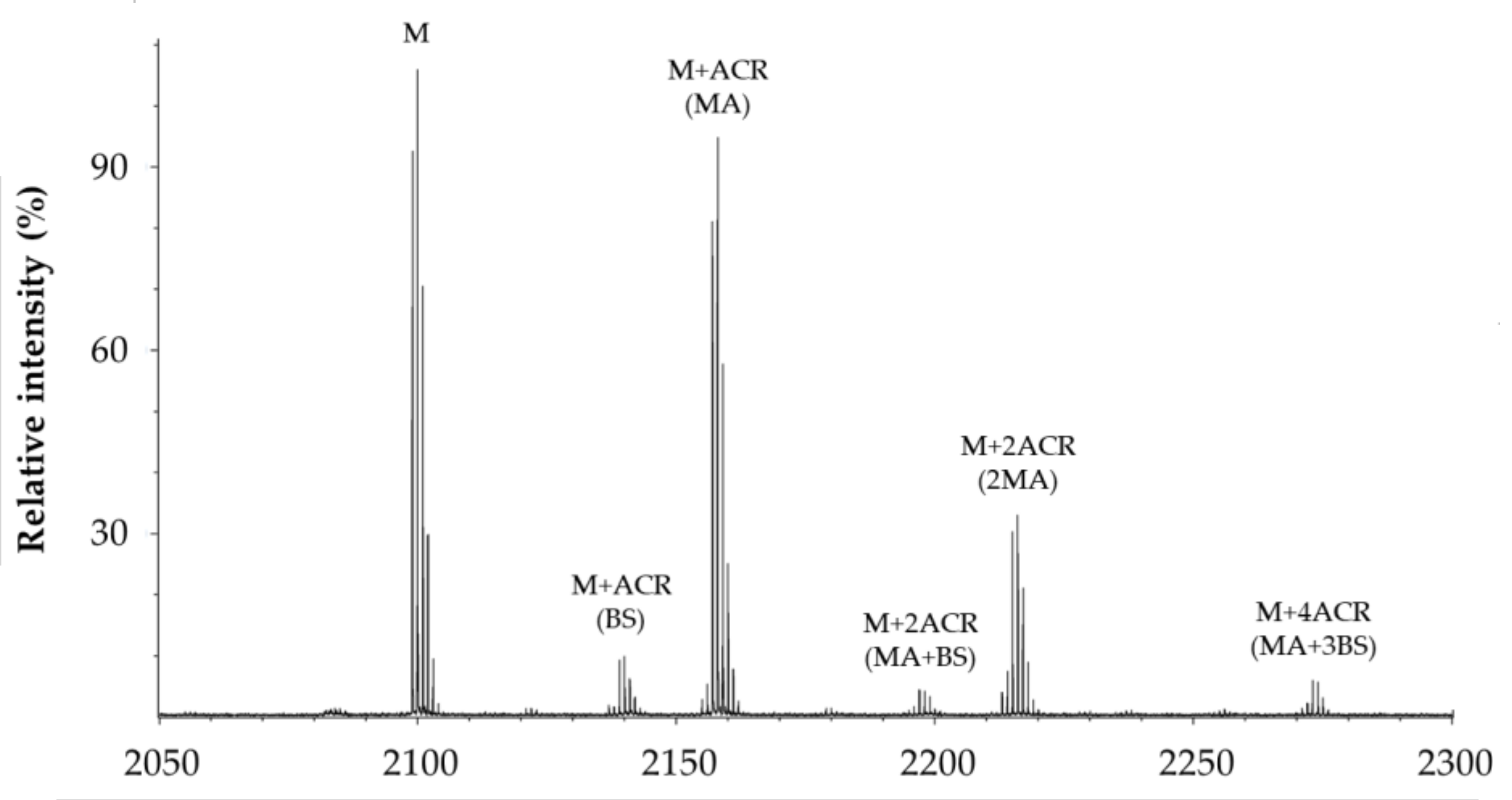
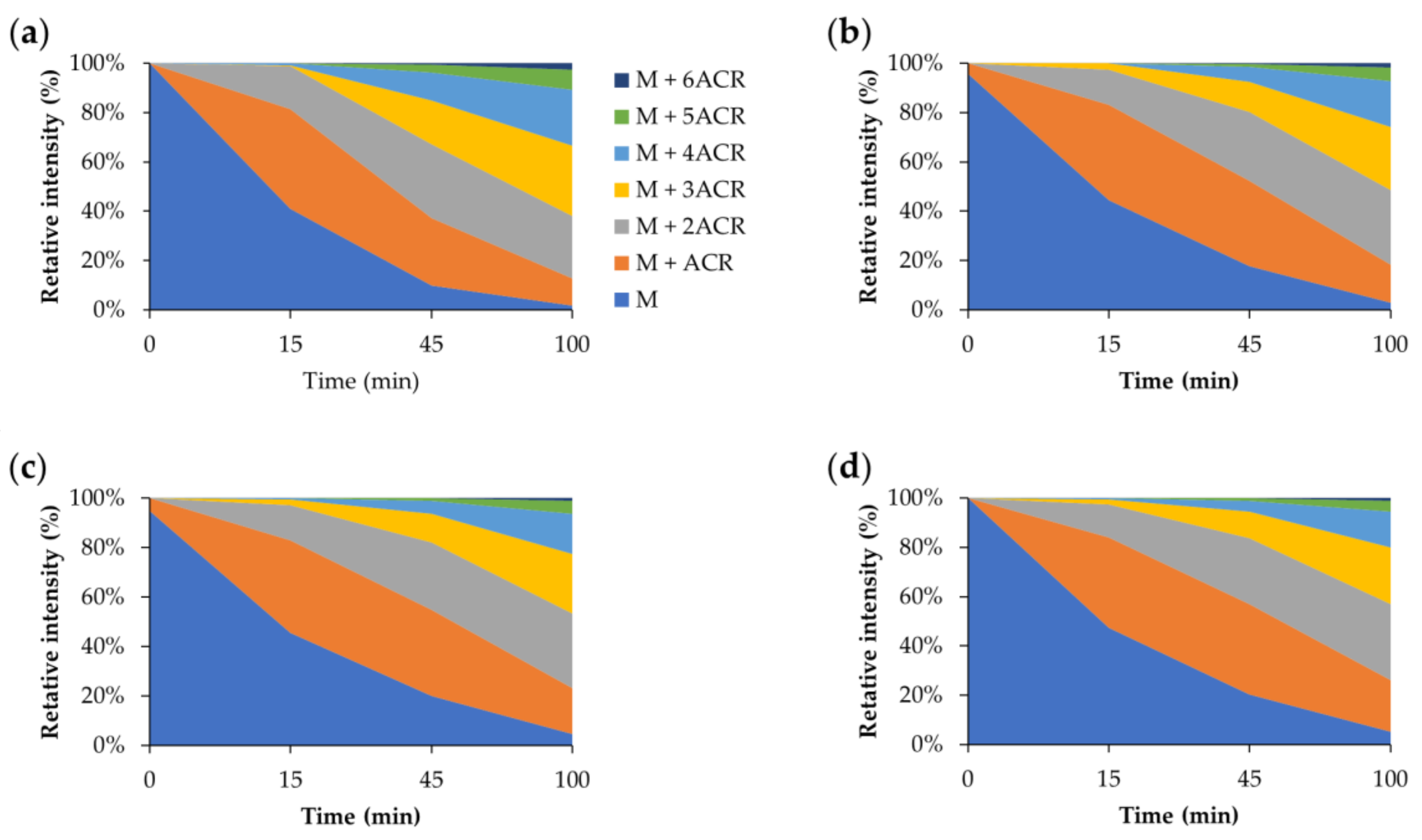
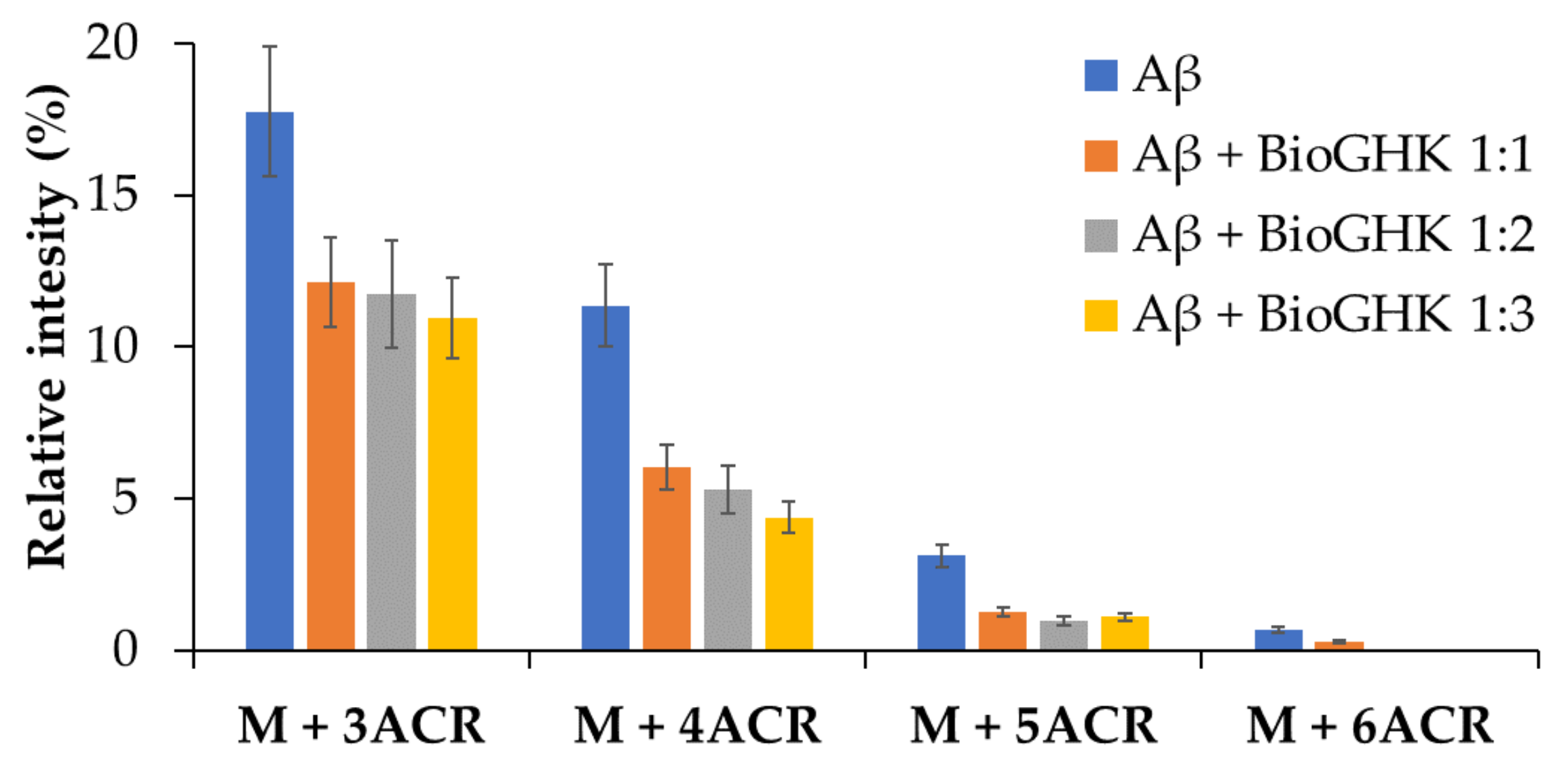
2.6. Effect of GHK and Derivative on Aβ Carbonylation
2.7. Effect of GHK and Derivative on Ascorbate Oxidation
2.8. Antiaggregant Activity towards Aβ
3. Materials and Methods
3.1. Synthesis of BioGHK
3.2. Interaction with Streptavidin
3.3. Copper(II) Complexes
3.4. Carbonylation of Aβ
3.5. Oxidation of Ascorbic Acid Catalyzed by Cu2+-Aβ
3.6. Aβ Aggregation Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Li, Z.; Sohrabi, H.R.; Sobrino, T.; Romaus-Sanjurjo, D. Editorial: New Challenges and Future Perspectives in Neurodegeneration. Front. Neurosci. 2022, 16, 1049721. [Google Scholar] [CrossRef]
- Wilson, D.M.; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Hallmarks of Neurodegenerative Diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Vallet, S.D. Proteases Decode the Extracellular Matrix Cryptome. Biochimie 2016, 122, 300–313. [Google Scholar] [CrossRef]
- Lane, T.F.; Iruela-Arispe, M.L.; Johnson, R.S.; Sage, E.H. SPARC Is a Source of Copper-Binding Peptides That Stimulate Angiogenesis. J. Cell Biol. 1994, 125, 929–943. [Google Scholar] [CrossRef] [PubMed]
- Pickart, L.; Margolina, A. Regenerative and Protective Actions of the GHK-Cu Peptide in the Light of the New Gene Data. Int. J. Mol. Sci. 2018, 19, 1987. [Google Scholar] [CrossRef] [PubMed]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The Human Tripeptide GHK-Cu in Prevention of Oxidative Stress and Degenerative Conditions of Aging: Implications for Cognitive Health. Oxid. Med. Cell. Longev. 2012, 2012, 324832. [Google Scholar] [CrossRef]
- Park, J.-R.; Lee, H.; Kim, S.-I.; Yang, S.-R. The Tri-Peptide GHK-Cu Complex Ameliorates Lipopolysaccharide-Induced Acute Lung Injury in Mice. Oncotarget 2016, 7, 58405–58417. [Google Scholar] [CrossRef]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. GHK Peptide as a Natural Modulator of Multiple Cellular Pathways in Skin Regeneration. BioMed Res. Int. 2015, 2015, 648108. [Google Scholar] [CrossRef]
- Budama-Kilinc, Y.; Kecel-Gunduz, S.; Cakir-Koc, R.; Aslan, B.; Bicak, B.; Kokcu, Y.; Ozel, A.E.; Akyuz, S. Structural Characterization and Drug Delivery System of Natural Growth-Modulating Peptide Against Glioblastoma Cancer. Int. J. Pept. Res. Ther. 2021, 27, 2015–2028. [Google Scholar] [CrossRef]
- Zoughaib, M.; Pavlov, R.V.; Gaynanova, G.A.; Garifullin, R.; Evtugyn, V.G.; Abdullin, T.I. Amphiphilic RGD and GHK Peptides Synergistically Enhance Liposomal Delivery into Cancer and Endothelial Cells. Mater. Adv. 2021, 2, 7715–7730. [Google Scholar] [CrossRef]
- Pickart, L.; Margolina, A. Modulation of Gene Expression in Human Breast Cancer Mcf7 and Prostate Cancer Pc3 Cells by the Human Copper-Binding Peptide Ghk-Cu. OBM Genet. 2021, 5, 128. [Google Scholar] [CrossRef]
- Kim, I.; Jin, S.M.; Han, E.H.; Ko, E.; Ahn, M.; Bang, W.Y.; Bang, J.K.; Lee, E. Structure-Dependent Antimicrobial Theranostic Functions of Self-Assembled Short Peptide Nanoagents. Biomacromolecules 2017, 18, 3600–3610. [Google Scholar] [CrossRef] [PubMed]
- Rajasekhar, K.; Madhu, C.; Govindaraju, T. Natural Tripeptide-Based Inhibitor of Multifaceted Amyloid β Toxicity. ACS Chem. Neurosci. 2016, 7, 1300–1310. [Google Scholar] [CrossRef]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The Effect of the Human Peptide GHK on Gene Expression Relevant to Nervous System Function and Cognitive Decline. Brain Sci. 2017, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, Q.; Wu, W.; Zhang, X.; Zuo, Z.; Lu, Y.; Zhao, H.; Wang, Z. Discovery of Novel Drug Candidates for Alzheimer’s Disease by Molecular Network Modeling. Front. Aging Neurosci. 2022, 14, 850217. [Google Scholar] [CrossRef]
- Ledwoń, P.; Errante, F.; Papini, A.M.; Rovero, P.; Latajka, R. Peptides as Active Ingredients: A Challenge for Cosmeceutical Industry. Chem. Biodivers. 2021, 18, e2000833. [Google Scholar] [CrossRef]
- Arul, V.; Gopinath, D.; Gomathi, K.; Jayakumar, R. Biotinylated GHK Peptide Incorporated Collagenous Matrix: A Novel Biomaterial for Dermal Wound Healing in Rats. J. Biomed. Mater. Res. B Appl. Biomater. 2005, 73, 383–391. [Google Scholar] [CrossRef]
- Jeong, S.; Yoon, S.; Kim, S.; Jung, J.; Kor, M.; Shin, K.; Lim, C.; Han, H.S.; Lee, H.; Park, K.Y.; et al. Anti-Wrinkle Benefits of Peptides Complex Stimulating Skin Basement Membrane Proteins Expression. Int. J. Mol. Sci. 2020, 21, 73. [Google Scholar] [CrossRef]
- Arul, V.; Kartha, R.; Jayakumar, R. A Therapeutic Approach for Diabetic Wound Healing Using Biotinylated GHK Incorporated Collagen Matrices. Life Sci. 2007, 80, 275–284. [Google Scholar] [CrossRef]
- Pardridge, W.M. Drug Transport across the Blood-Brain Barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1959–1972. [Google Scholar] [CrossRef]
- Waner, M.J.; Hiznay, J.M.; Mustovich, A.T.; Patton, W.; Ponyik, C.; Mascotti, D.P. Streptavidin Cooperative Allosterism upon Binding Biotin Observed by Differential Changes in Intrinsic Fluorescence. Biochem. Biophys. Rep. 2019, 17, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Grasso, G.; Magrì, A.; Bellia, F.; Pietropaolo, A.; La Mendola, D.; Rizzarelli, E. The Copper(II) and Zinc(II) Coordination Mode of HExxH and HxxEH Motif in Small Peptides: The Role of Carboxylate Location and Hydrogen Bonding Network. J. Inorg. Biochem. 2014, 130, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Grasso, G.I.; Arena, G.; Bellia, F.; Rizzarelli, E.; Vecchio, G. Copper(II)-Chelating Homocarnosine Glycoconjugate as a New Multifunctional Compound. J. Inorg. Biochem. 2014, 131, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Oliveri, V.; Rizzarelli, E.; Vecchio, G. New Derivative of Carnosine for Nanoparticle Assemblies. Eur. J. Med. Chem. 2013, 70, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Grasso, G.I.; Ahmed, I.M.M.; Oliveri, V.; Vecchio, G. Carnoquinolines Target Copper Dyshomeostasis, Aberrant Protein–Protein Interactions, and Oxidative Stress. Chem. Eur. J. 2020, 26, 16690–16705. [Google Scholar] [CrossRef]
- Huang, Y.J.; Jin, M.H.; Pi, R.B.; Zhang, J.J.; Ouyang, Y.; Chao, X.J.; Chen, M.H.; Liu, P.Q.; Yu, J.C.; Ramassamy, C.; et al. Acrolein Induces Alzheimer’s Disease-like Pathologies in Vitro and in Vivo. Toxicol. Lett. 2013, 217, 184–191. [Google Scholar] [CrossRef]
- Gómez-Ramos, A.; Díaz-Nido, J.; Smith, M.A.; Perry, G.; Avila, J. Effect of the Lipid Peroxidation Product Acrolein on Tau Phosphorylation in Neural Cells. J. Neurosci. Res. 2003, 71, 863–870. [Google Scholar] [CrossRef]
- Esmieu, C.; Guettas, D.; Conte-Daban, A.; Sabater, L.; Faller, P.; Hureau, C. Copper-Targeting Approaches in Alzheimer’s Disease: How to Improve the Fallouts Obtained from in Vitro Studies. Inorg. Chem. 2019, 58, 13509–13527. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Functional Amyloid, and Human Disease. Ann. Rev. Biochem. 2006, 75, 333–366. [Google Scholar] [CrossRef]
- Oliveri, V.; Bellia, F.; Vecchio, G. Cyclodextrin Nanoparticles Bearing 8-Hydroxyquinoline Ligands as Multifunctional Biomaterials. Chem. Eur. J. 2017, 23, 4442–4449. [Google Scholar] [CrossRef]
- Greco, V.; Naletova, I.; Ahmed, I.M.M.; Vaccaro, S.; Messina, L.; La Mendola, D.; Bellia, F.; Sciuto, S.; Satriano, C.; Rizzarelli, E. Hyaluronan-Carnosine Conjugates Inhibit Aβ Aggregation and Toxicity. Sci. Rep. 2020, 10, 15998. [Google Scholar] [CrossRef] [PubMed]
- Lanza, V.; Bellia, F.; Rizzarelli, E. An Inorganic Overview of Natural Aβ Fragments: Copper(II) and Zinc(II)-Mediated Pathways. Coord. Chem. Rev. 2018, 369, 1–14. [Google Scholar] [CrossRef]
- Cheignon, C.; Collin, F.; Faller, P.; Hureau, C. Is Ascorbate Dr Jekyll or Mr Hyde in the Cu(Aβ) Mediated Oxidative Stress Linked to Alzheimer’s Disease? Dalton Trans. 2016, 45, 12627–12631. [Google Scholar] [CrossRef] [PubMed]
- Grasso, G.I.; Bellia, F.; Arena, G.; Satriano, C.; Vecchio, G.; Rizzarelli, E. Multitarget Trehalose-Carnosine Conjugates Inhibit Aβ Aggregation, Tune Copper(II) Activity and Decrease Acrolein Toxicity. Eur. J. Med. Chem. 2017, 135, 447–457. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | m/z Calc. | m/z Exp. |
|---|---|---|
| M | 581.30 | 581.31 |
| M–H2O | 563.29 | 563.30 |
| M–2H2O | 545.28 | 545.27 |
| y3 | 524.28 | 524.26 |
| y2 | 387.22 | 387.23 |
| b3 | 323.18 | 323.17 |
| b3–H2O | 305.17 | 305.16 |
| b3–H2O–NH3 | 288.15 | 288.15 |
| y1 | 259.12 | 259.11 |
| b2 | 195.09 | 195.10 |
| b1 | 58.03 | 58.03 |
| Species | m/z Calc. | m/z Exp. | Δm |
|---|---|---|---|
| M | 2099.064 | 2098.969 | 0.095 |
| M + ACR (BS) | 2137.010 | 2136.985 | 0.025 |
| M + ACR (MA) | 2157.106 | 2157.011 | 0.095 |
| M + 2ACR (MP) | 2177.169 | 2178.023 | −0.854 |
| M + 2ACR (MA + BS) | 2195.121 | 2195.026 | 0.095 |
| M + 2ACR (2MA) | 2215.149 | 2215.053 | 0.096 |
| M + 3ACR (MP + MA) | 2235.207 | 2235.021 | 0.186 |
| M + 3ACR (2MA + BS) | 2253.166 | 2253.068 | 0.098 |
| M + 4ACR (1MA + 3BS) | 2271.179 | 2271.058 | 0.121 |
| M + 3ACR (3MA) | 2273.148 | 2273.094 | 0.054 |
| M + 4ACR (2MA + 2BS) | 2291.186 | 2291.048 | 0.138 |
| M + 4ACR (MP + 2MA) | 2293.252 | 2293.100 | 0.152 |
| M + 4ACR (3MA + BS) | 2311.207 | 2311.11 | 0.097 |
| M + 5ACR (2MA + 3BS) | 2329.140 | 2329.100 | 0.040 |
| M + 4ACR | 2331.229 | 2331.136 | 0.093 |
| M + 5ACR (MP + 3MA) | 2351.317 | 2351.140 | 0.177 |
| M + 5ACR (4MA + BS) | 2369.337 | 2369.152 | 0.185 |
| M + 6ACR (3MA + 3BS) | 2387.343 | 2387.142 | 0.201 |
| CTRL | Bio | GHK | Bio + GHK | BioGHK | |
|---|---|---|---|---|---|
| Fmax − F0 | 23 ± 3 | 18 ± 4 | 15 ± 4 | 14 ± 2 | 6 ± 2 |
| tlag | 26 ± 6 | 25 ± 4 | 25 ± 5 | 23 ± 3 | 24 ± 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tosto, R.; Vecchio, G.; Bellia, F. New Biotinylated GHK and Related Copper(II) Complex: Antioxidant and Antiglycant Properties In Vitro against Neurodegenerative Disorders. Molecules 2023, 28, 6724. https://doi.org/10.3390/molecules28186724
Tosto R, Vecchio G, Bellia F. New Biotinylated GHK and Related Copper(II) Complex: Antioxidant and Antiglycant Properties In Vitro against Neurodegenerative Disorders. Molecules. 2023; 28(18):6724. https://doi.org/10.3390/molecules28186724
Chicago/Turabian StyleTosto, Rita, Graziella Vecchio, and Francesco Bellia. 2023. "New Biotinylated GHK and Related Copper(II) Complex: Antioxidant and Antiglycant Properties In Vitro against Neurodegenerative Disorders" Molecules 28, no. 18: 6724. https://doi.org/10.3390/molecules28186724
APA StyleTosto, R., Vecchio, G., & Bellia, F. (2023). New Biotinylated GHK and Related Copper(II) Complex: Antioxidant and Antiglycant Properties In Vitro against Neurodegenerative Disorders. Molecules, 28(18), 6724. https://doi.org/10.3390/molecules28186724