
Oxidation of Cathepsin D by Hydroxy Radical: Its Effect on Enzyme Structure and Activity against Myofibrillar Proteins Extracted from Coregonus peled

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Activity of Cathepsin D in Hydroxyl Radical Oxidation System
2.2. Changes in the Carbonyl Content of Cathepsin D
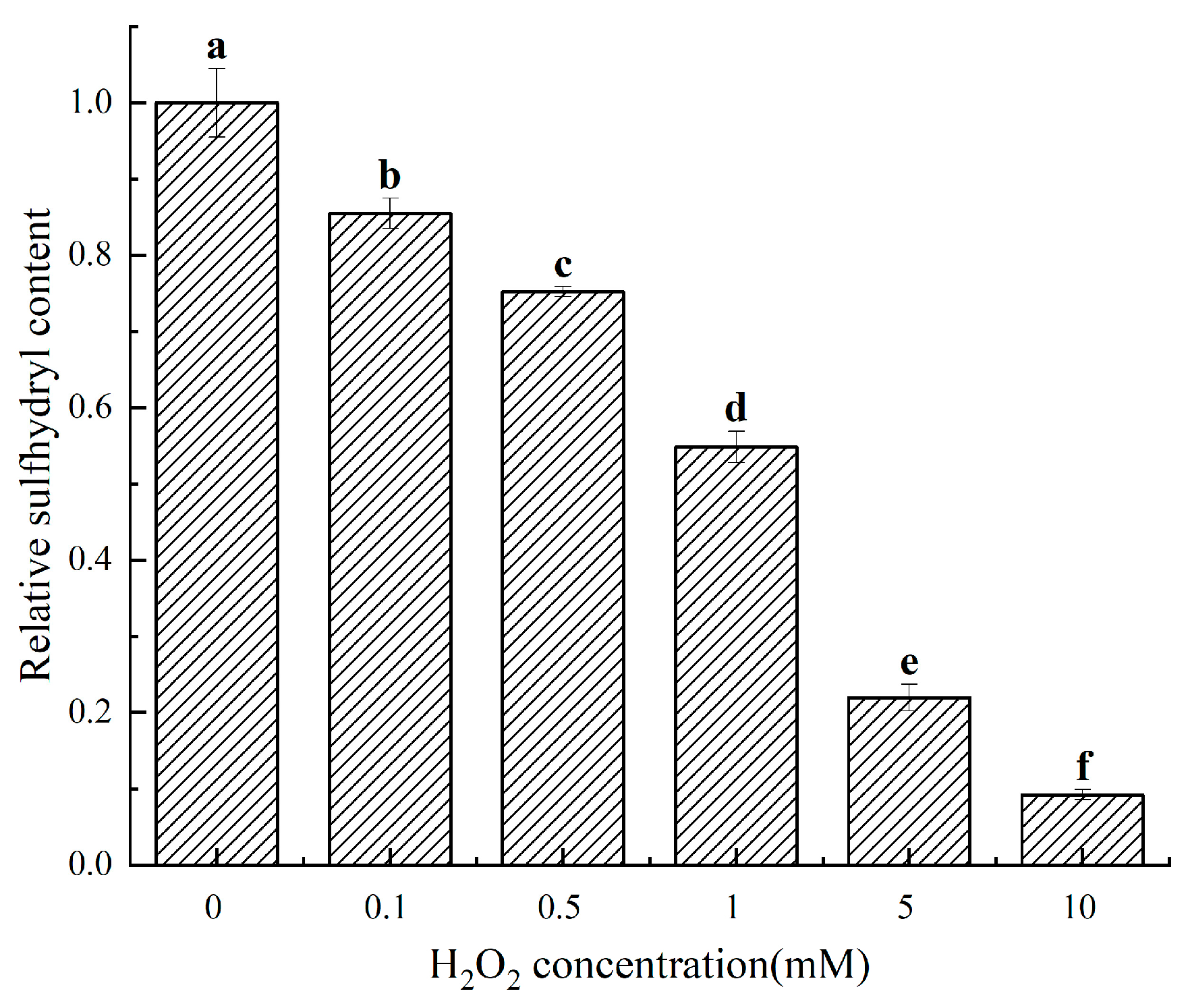
2.3. Changes in the Sulfhydryl Content of Cathepsin D
2.4. Structural Changes in Cathepsin D
2.4.1. Ultraviolet Absorption Spectrum Analysis of Cathepsin D
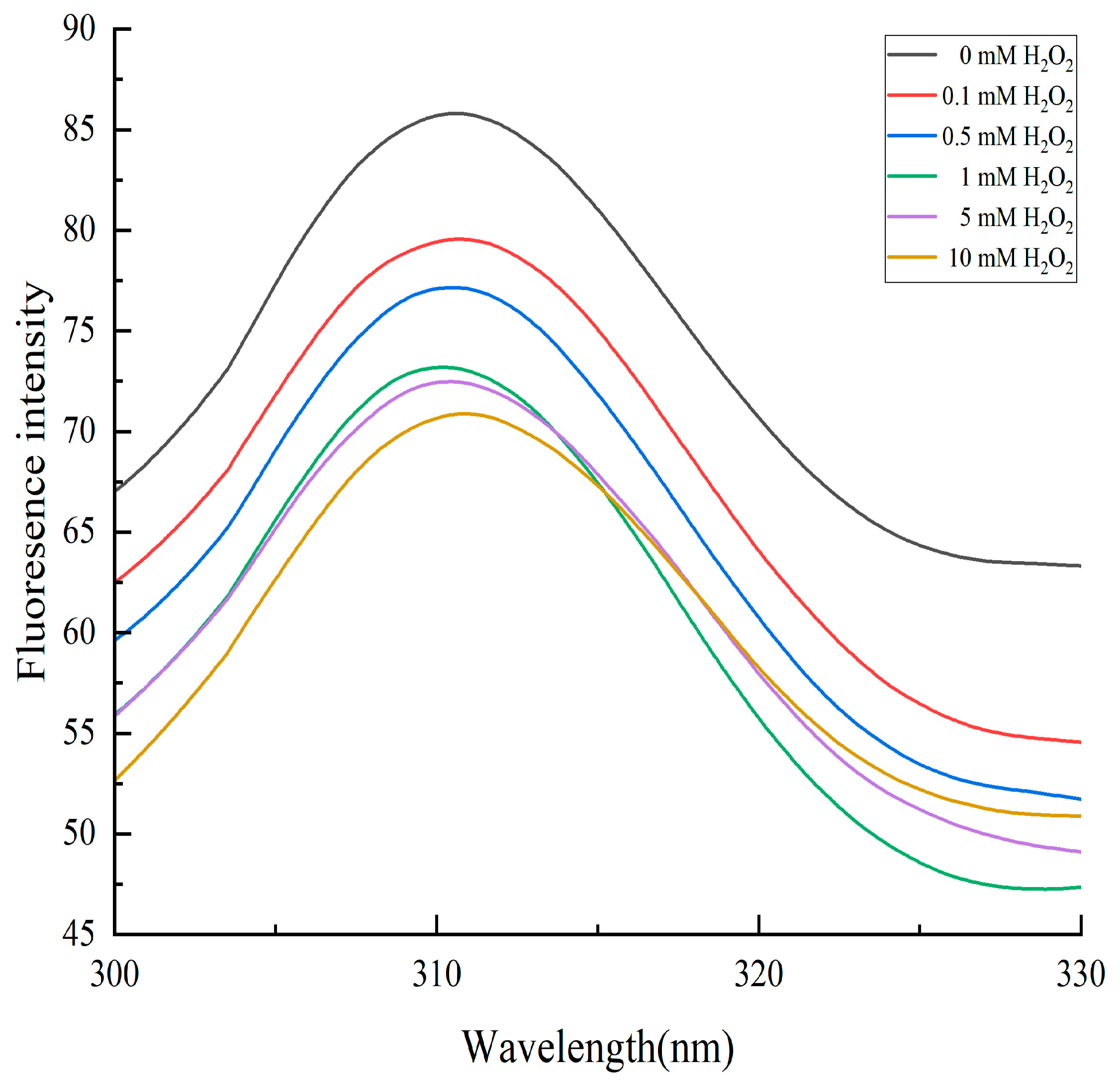
2.4.2. Changes in Endogenous Fluorescence Intensity of Cathepsin D
2.4.3. Circular Dichroism (CD) Analysis
2.5. Analysis of Myofibrillar Protein Degradation by Oxidized Cathepsin D In Vitro
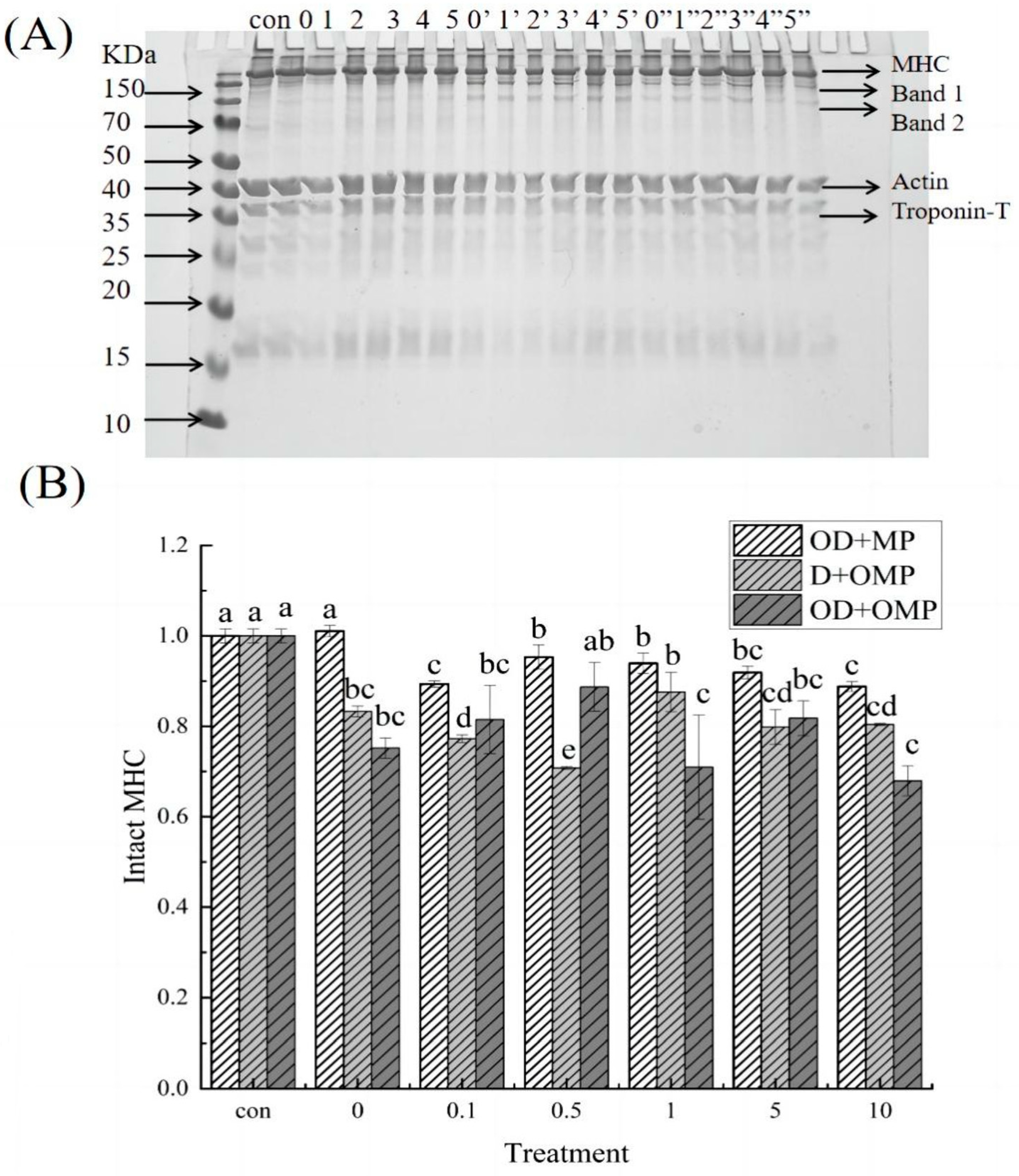
2.5.1. SDS-PAGE
2.5.2. Myosin Heavy Chain (MHC)
2.5.3. Actin
2.5.4. Desmin
2.5.5. Troponin-T
3. Materials and Methods
3.1. Sample Descriptions
3.2. Preparation of Myofibrillar Protein (MP)
3.3. Preparation of the Hydroxyl Radical Oxidation System
3.4. Measurement of the Activity of Cathepsin D
3.5. Measurement of the Carbonyl Content of Cathepsin D
3.6. Determination of the Sulfhydryl Content of Cathepsin D
3.7. Measurement of the Intrinsic Fluorescence of Cathepsin D
3.8. Circular Dichroism (CD) Analysis of Cathepsin D
3.9. Ultraviolet (UV) Absorption of Cathepsin D
3.10. Incubation of the Cathepsin D with the MP
3.11. SDS-PAGE and Western Blotting Analyses
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jiang, W.; He, Y.; Xiong, S.; Liu, Y.; Yin, T.; Hu, Y.; You, J. Effect of mild ozone oxidation on structural changes of silver carp (Hypophthalmichthys molitrix) Myosin. Food Bioprocess Technol. 2017, 10, 370–380. [Google Scholar] [CrossRef]
- Dulmaa, A.; Slynko, Y.V.; Gordon, N.Y.; Stolbunova, V.V.; Politov, D.V. Coregonus peled (Gmelin) transplanted into Ulaagchny Khar lake (western mongolia) showed no evidence of hybridization with other introduced Coregonus species. Contemp. Probl. Ecol. 2016, 9, 172–176. [Google Scholar] [CrossRef]
- Sentandreu, M.; Coulis, G.; Ouali, A. Role of muscle endopeptidases and their inhibitors in meat tenderness. Trends Food Sci. Technol. 2002, 13, 400–421. [Google Scholar] [CrossRef]
- Mukundan, M.; Antony, P.; Nair, M. A review on autolysis in fish. Fish. Res. 1986, 4, 259–269. [Google Scholar] [CrossRef]
- Szymczak, M.; Lepczyński, A. Occurrence of aspartyl proteases in brine after herring marinating. Food Chem. 2016, 194, 470–475. [Google Scholar] [CrossRef]
- Cho, W.L.; Raikhel, A.S. Cloning of cDNA for mosquito lysosomal aspartic protease. Sequence analysis of an insect lysosomal enzyme similar to cathepsins D and E. J. Biol. Chem. 1992, 267, 21823–21829. [Google Scholar] [CrossRef]
- Glickman, J.N.; Kornfeld, S. Mannose 6-phosphate-independent targeting of lysosomal enzymes in I-cell disease B lymphoblasts. J. Cell Biol. 1993, 123, 99–108. [Google Scholar] [CrossRef]
- Zhao, S.; Aviles, E.R., Jr.; Fujikawa, D.G. Nuclear translocation of mitochondrial cytochrome c, lysosomal cathepsins B and D, and three other death-promoting proteins within the first 60 minutes of generalized seizures. J. Neurosci. Res. 2010, 88, 1727–1737. [Google Scholar] [CrossRef]
- Bańkowska, A.; Gacko, M.; Chyczewska, E.; Worowska, A. Biological and diagnostic role of cathepsin D. Rocz. Akad. Med. W Bialymst. 1997, 42 (Suppl. S1), 79–85. [Google Scholar]
- Kim, K.; Homma, Y.; Ikeuchi, Y.; Suzuki, A. Cleavageof connectun by calpain ang cathepsin-D. Biosci. Biotechnol. Biochem. 1995, 59, 896–899. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, P.; Deng, X.; Guo, X.; Mao, X.; Guo, X.; Zhang, J. Effects of hydroxyl radical oxidation on myofibrillar protein and its susceptibility to μ-calpain proteolysis. LWT—Food Sci. Technol. 2021, 137, 110453. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Berlett, B.S. Reactive oxygen-mediated protein oxidation in aging and disease. Drug Metab. Rev. 1998, 30, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Huang, F.; Huang, M.; Zhou, G. Influence of oxidation on myofibrillar proteins degradation from bovine via mu-calpain. Food Chem. 2012, 134, 106–112. [Google Scholar] [CrossRef]
- Chan, J.T.; Omana, D.A.; Betti, M. Effect of ultimate pH and freezing on the biochemical properties of proteins in turkey breast meat. Food Chem. 2011, 127, 109–117. [Google Scholar] [CrossRef]
- Ladror, U.S.; Snyder, S.W.; Wang, G.T.; Holzman, T.F.; Krafft, G.A. Cleavege of the amino and carboxyl termini of alzheimers amyloid-beta by cathepsin-G. J. Biol. Chem. 1994, 269, 18422–18428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Scott, S.; Shelton, S.; Crutcher, K. Cathepsin D-mediated proteolysis of apolipoprotein E: Possible role in Alzheimer’s disease. Neuroscience 2006, 143, 689–701. [Google Scholar] [CrossRef]
- Kenessey, A.; Nacharaju, P.; Ko, L.W.; Yen, S.H. Degradation of tau by lysosomal enzyme cathepsin D: Implication for Alzheimer neurofibrillary degeneration. J. Neurochem. 1997, 69, 2026–2038. [Google Scholar] [CrossRef]
- Eymard, S.; Baron, C.P.; Jacobsen, C. Oxidation of lipid and protein in horse mackerel (Trachurus trachurus) mince and washed minces during processing and storage. Food Chem. 2009, 114, 57–65. [Google Scholar] [CrossRef]
- Morzel, M.; Gatellier, P.; Sayd, T.; Renerre, M.; Laville, E. Chemical oxidation decreases proteolytic susceptibility of skeletal muscle myofibrillar proteins. Meat Sci. 2006, 73, 536–543. [Google Scholar] [CrossRef]
- Zhang, D.; Li, H.; Emara, A.; Hu, Y.; Wang, Z.; Wang, M.; He, Z. Effect of in vitro oxidation on the water retention mechanism of myofibrillar proteins gel from pork muscles. Food Chem. 2020, 315, 126226. [Google Scholar] [CrossRef]
- Bao, Y.; Ertbjerg, P. Effects of protein oxidation on the texture and water-holding of meat: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3564–3578. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Liu, R.; Wang, H.; Hua, C.; Song, S.; Zhou, G.; Zhang, W. Effects of Oxidation in vitro on structures and functions of myofibrillar protein from beef muscles. J. Agric. Food Chem. 2019, 67, 5866–5873. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; He, Z.; Gan, X.; Li, H. Effect of peroxyl radicals on the structure and gel properties of isolated rabbit meat myofibrillar proteins. Int. J. Food Sci. Technol. 2018, 53, 2687–2696. [Google Scholar] [CrossRef]
- Tong, P.; Gao, J.; Chen, H.; Li, X.; Zhang, Y.; Jian, S.; Wichers, H.; Wu, Z.; Yang, A.; Liu, F. Effect of heat treatment on the potential allergenicity and conformational structure of egg allergen ovotransferrin. Food Chem. 2012, 131, 603–610. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Yuan, F.; Gao, Q.; Ma, J. Effect of 4-hydroxy-2-nonenal treatment on the IgE binding capacity and structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem. 2016, 212, 313–322. [Google Scholar] [CrossRef]
- Chizoba Ekezie, F.G.; Cheng, J.H.; Sun, D.W. Effects of mild oxidative and structural modifications induced by argon plasma on physicochemical properties of actomyosin from king prawn (Litopenaeus vannamei). J. Agric. Food Chem. 2018, 66, 13285–13294. [Google Scholar] [CrossRef]
- Guo, X.; Qiu, H.; Deng, X.; Mao, X.; Xu, C.; Zhang, J. Effect of chlorogenic acid on the physicochemical and functional properties of coregonus peled myofibrillar protein through hydroxyl radical oxidation. Molecules 2019, 24, 3205. [Google Scholar] [CrossRef]
- Rogers, D.M.; Jasim, S.B.; Dyer, N.T.; Auvray, F.; Réfrégiers, M.; Hirst, J.D. Electronic circular dichroism spectroscopy of proteins. Chemistry 2019, 5, 2751–2774. [Google Scholar] [CrossRef]
- Metcalf, P.; Fusek, M. Two crystal structures for cathepsin D: The lysosomal targeting signal and active site. EMBO J. 1993, 12, 1293–1302. [Google Scholar] [CrossRef]
- Chttpinttyol, S.; Crabbe, M.J.C. Chymosin and aspartic proteinases. Food Chem. 1998, 61, 395–418. [Google Scholar] [CrossRef]
- Sun, W.; Cui, C.; Zhao, M.; Zhao, Q.; Yang, B. Effects of composition and oxidation of proteins on their solubility, aggregation and proteolytic susceptibility during processing of Cantonese sausage. Food Chem. 2011, 124, 336–341. [Google Scholar] [CrossRef]
- Lametsch, R.; Roepstorff, P.; Møller, H.; Bendixen, E. Identification of myofibrillar substrates for μ-calpain. Meat Sci. 2004, 68, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Z.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.-D.A. Pulsed electric field operates enzymatically by causing early activation of calpains in beef during ageing. Chemistry 2019, 153, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.Y.; Strmoer, M.H.; Robson, R.M. Identification of the 30 kDa polypeptide in post mortem skeletal muscle as a degradation product of troponin-T. Biochimie 1994, 76, 369–375. [Google Scholar] [CrossRef]
- Lu, H.; Liu, X.; Zhang, Y.; Wang, H.; Luo, Y. Effects of chilling and partial freezing on rigor mortis changes of bighead carp (Aristichthys nobilis) fillets: Cathepsin activity, protein degradation and microstructure of myofibrils. J. Food Sci. 2015, 80, C2725–C2731. [Google Scholar] [CrossRef]
- Levin, R.; Brauer, R.W. The biuret reaction for the determination of proteins; an improved reagent and its application. J. Lab. Clin. Med. 1951, 38, 474–480. [Google Scholar]
- Park, D.; Xiong, Y.L.; Alderton, A.L. Concentration effects of hydroxyl radical oxidizing systems on biochemical properties of porcine muscle myofibrillar protein. Food Chem. 2007, 101, 1239–1246. [Google Scholar] [CrossRef]
- Minarowska, A.; Karwowska, A.; Gacko, M. Quantitative determination and localization of cathepsin D and its inhibitors. Folia Histochem. Cytobiol 2009, 47, 153–177. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Oliver, C.N.; Ahn, B.-W.; Moermant, E.J.; Goldstein, S.; Stadtman, E.R. Age-related changes in oxidized proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [CrossRef]
- Qin, J.; Deng, X.; Lei, Y.; Liu, P.; Lu, S.; Zhang, J. Effects of µ-calpain oxidation on Coregonus peled myofibrillar protein degradation in vitro. J. Food Sci. 2020, 85, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Lin, H.; Li, Z.; Wang, J.; Ahmed, I.; Chen, H. Changes of structure and IgE binding capacity of shrimp (Metapenaeus ensis) tropomyosin followed by acrolein treatment. Food Funct. 2017, 8, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Xia, W.; Jiang, Q. Pressure-induced changes of silver carp (Hypophthalmichthys molitrix) myofibrillar protein structure. Eur. Food Res. Technol. 2014, 238, 753–761. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Z.; Guo, X.; Zhu, X.; Mao, X.; Guo, X.; Deng, X.; Zhang, J. μ-Calpain oxidation and proteolytic changes on myofibrillar proteins from Coregonus Peled in vitro. Food Chem. 2021, 361, 130100. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavege of structural proteins during assembly of head od bacteriophage-T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, M.; Liu, P.; Wang, C.; Deng, X.; Zhang, L.; Zhang, J. Oxidation of Cathepsin D by Hydroxy Radical: Its Effect on Enzyme Structure and Activity against Myofibrillar Proteins Extracted from Coregonus peled. Molecules 2023, 28, 5117. https://doi.org/10.3390/molecules28135117
Ma M, Liu P, Wang C, Deng X, Zhang L, Zhang J. Oxidation of Cathepsin D by Hydroxy Radical: Its Effect on Enzyme Structure and Activity against Myofibrillar Proteins Extracted from Coregonus peled. Molecules. 2023; 28(13):5117. https://doi.org/10.3390/molecules28135117
Chicago/Turabian StyleMa, Mengjie, Pingping Liu, Chaoye Wang, Xiaorong Deng, Lianfu Zhang, and Jian Zhang. 2023. "Oxidation of Cathepsin D by Hydroxy Radical: Its Effect on Enzyme Structure and Activity against Myofibrillar Proteins Extracted from Coregonus peled" Molecules 28, no. 13: 5117. https://doi.org/10.3390/molecules28135117
APA StyleMa, M., Liu, P., Wang, C., Deng, X., Zhang, L., & Zhang, J. (2023). Oxidation of Cathepsin D by Hydroxy Radical: Its Effect on Enzyme Structure and Activity against Myofibrillar Proteins Extracted from Coregonus peled. Molecules, 28(13), 5117. https://doi.org/10.3390/molecules28135117