
Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus


Abstract
1. Introduction
2. Pathophysiology of T2DM
3. Oxidative Stress in T2DM Pathogenesis
3.1. Glucose Influx through the Polyol Pathway
- (i)
- The reduction of glucose to sorbitol by aldose reductase (AR) with the help of its cofactor NADPH. In the state of hyperglycemia, about 30% of glucose is metabolized by the polyol pathway. The increased activity of AR in converting glucose has caused a depletion in its cofactor NADPH, which is also essential for the production of glutathione (GSH). GSH is an important cellular antioxidant that is capable of preventing ROS damage to cellular components. The decrease in GSH level has weakened the antioxidant capacity, thus favoring the condition for oxidative stress.
- (ii)
3.2. Intracellular Production of AGEs
3.3. PKC Activation Pathway
3.4. Hexosamine Pathway
3.5. Relationship between Oxidative Stress and T2DM
4. Lycopene
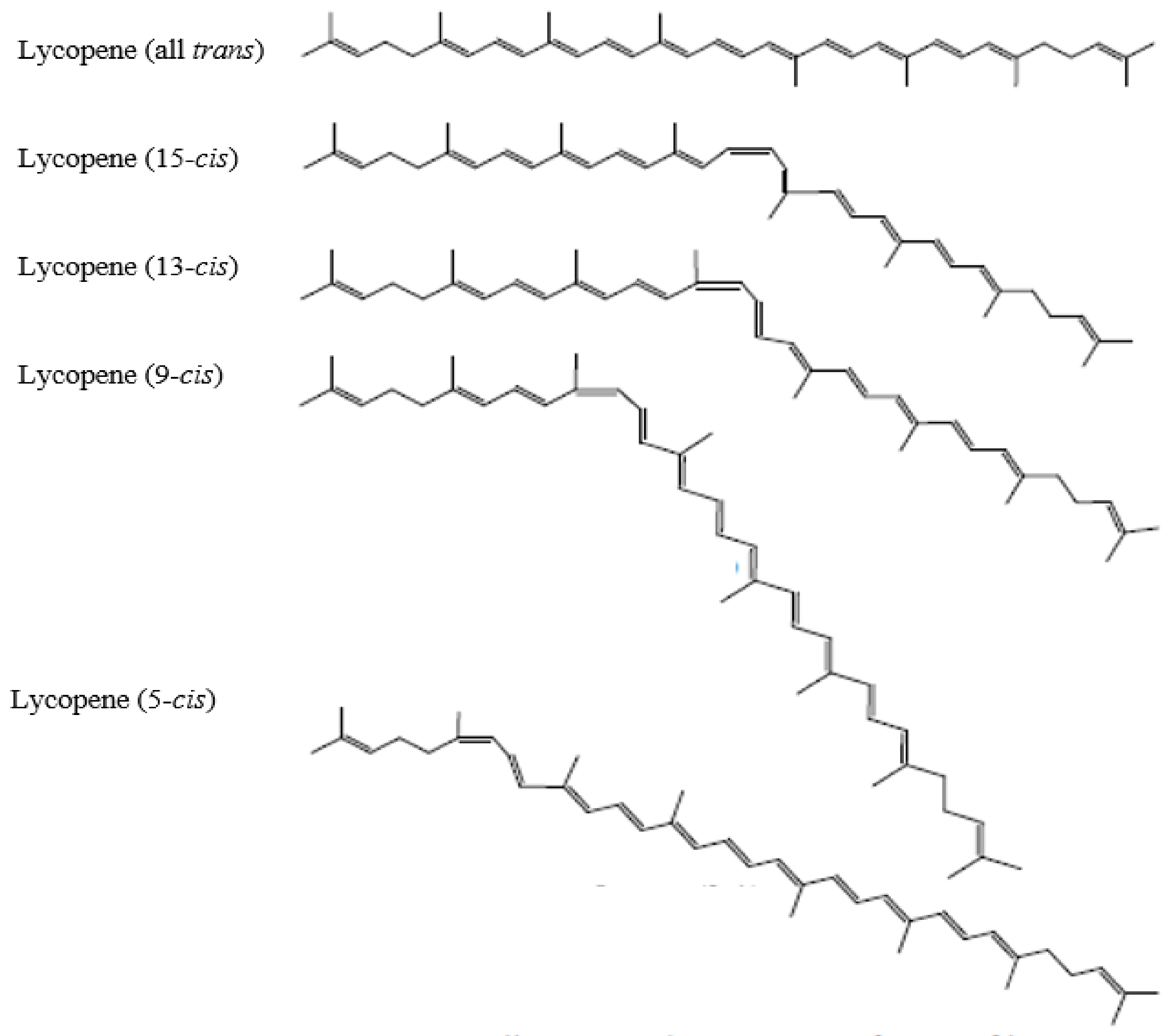
4.1. Chemistry and Physical Properties
4.2. Dietary Sources
4.2.1. Natural Lycopene
4.2.2. Synthetic Lycopene
4.2.3. Lycopene from Blakeslea trispora
4.3. The Use of Lycopene and Acceptable Daily Intake (ADI)
5. Lycopene as Antioxidant
5.1. The Mechanisms of Action of Lycopene in Scavenging Reactive Species
5.2. Synergistic Effect of Lycopene with Other Antioxidants
6. Lycopene Consumption and T2DM
6.1. Lycopene Status in T2DM Patients
6.2. Animal Studies: Lycopene Effects on Glycemic Control and Oxidative Stress Biomarkers
6.3. Human Studies: Lycopene Effects on Glycemic Control
6.4. Human Studies: Lycopene Effects on Oxidative Stress Biomarkers and Risk of T2DM
7. Mechanisms of Action of Lycopene in T2DM
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas. Available online: https://diabetesatlas.org/ (accessed on 18 January 2022).
- Skyler, J.S.; Bakris, G.L.; Bonifacio, E.; Darsow, T.; Eckel, R.H.; Groop, L.; Groop, P.H.; Handelsman, Y.; Insel, R.A.; Mathieu, C.; et al. Differentiation of diabetes by pathophysiology, natural history, and prognosis. Diabetes 2017, 66, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Ganasegeran, K.; Hor, C.P.; Jamil, M.; Loh, H.C.; Noor, J.M.; Hamid, N.A.; Suppiah, P.D.; Abdul Manaf, M.R.; Ch’ng, A.; Looi, I. A systematic review of the economic burden of type 2 diabetes in Malaysia. Int. J. Env. Res. Public Health 2020, 17, 5723. [Google Scholar] [CrossRef] [PubMed]
- Upritchard, J.E.; Sutherland, W.H.F.; Mann, J.I. Effect of supplementation with tomato juice, vitamin E, and vitamin C on LDL oxidation and products of inflammatory activity in type 2 diabetes. Diabetes Care 2000, 23, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Neyestani, T.; Shariatzadeh, N.; Gharavi, A.; Kalayi, A.; Khalaji, N. Physiological dose of lycopene suppressed oxidative stress and enhanced serum levels of immunoglobulin M in patients with Type 2 diabetes mellitus: A possible role in the prevention of long-term complications. J. Endocrinol. Investig. 2007, 30, 833–838. [Google Scholar] [CrossRef]
- Ozmutlu, S.; Dede, S.; Ceylan, E. The effect of lycopene treatment on ACE activity in rats with experimental diabetes. J. Renin-Angiotensin-Aldosterone Syst. 2012, 13, 328–333. [Google Scholar] [CrossRef]
- Yegin, S.Ç.; Yur, F.; Ceylan, E. Effect of lycopene application in rats with experimental diabetes using lipoprotein, paraoxonase and cytokines. J. Membr. Biol. 2013, 246, 621–626. [Google Scholar] [CrossRef]
- Xia, X.; Xiao, J. Natural ingredients from medicine food homology as chemopreventive reagents against type 2 diabetes mellitus by modulating gut microbiota homoeostasis. Molecules 2021, 26, 6934. [Google Scholar] [CrossRef]
- Asif, M. The prevention and control the type-2 diabetes by changing lifestyle and dietary pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Nur Kaplan, D.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M.; et al. Lycopene: Food sources, biological activities, and human health benefits. Oxid. Med. Cell. Longev. 2021, 2021, 2713511. [Google Scholar] [CrossRef]
- Montonen, J.; Knekt, P.; Jarvinen, R.; Reunanen, A. Dietary antioxidant intake and risk of type 2 diabetes: Response to Granado-Lorencio and Olmedilla-Alonso. Diabetes Care 2004, 27, 1845–1846. [Google Scholar] [CrossRef][Green Version]
- Rani, A.J.; Mythili, S.V. Study on total antioxidant status in relation to oxidative stress in type 2 diabetes mellitus. J. Clin. Diagnos. Res. 2014, 8, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, S.; Manson, J.; Gaziano, J.; Buring, J.; Sesso, H. The consumption of lycopene and tomato-based food products is not associated with the risk of type 2 diabetes in women. J. Nutr. 2006, 136, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Bal, B.S.; Chopra, S.; Singh, S.; Malhotra, N. Ameliorative effect of lycopene on lipid peroxidation and certain antioxidant enzymes in diabetic patients. J. Diabetes Metab. 2012, 3, 1–5. [Google Scholar] [CrossRef]
- DeFronzo, R.A. The triumvirate: β-cell, muscle, liver. A collusion responsible for NIDDM. Diabetes 1988, 37, 667–687. [Google Scholar] [CrossRef]
- Meyer, C.; Dostou, J.M.; Welle, S.L.; Gerich, J.E. Role of human liver, kidney, and skeletal muscle in postprandial glucose homeostasis. Am. J. Physiol.-Endocrinol. Metab. 2002, 282, 419–427. [Google Scholar] [CrossRef]
- Holt, R.I.G. Diagnosis, epidemiology and pathogenesis of diabetes mellitus: An update for psychiatrists. Br. J. Psychiatry 2004, 184, 55–64. [Google Scholar] [CrossRef]
- Belfort, R.; Mandarino, L.; Kashyap, S.; Wirfel, K.; Pratipanawatr, T.; Berria, R.; Defronzo, R.A.; Cusi, K. Dose-response effect of elevated plasma free fatty acid on insulin signaling. Diabetes 2005, 4, 1640–1648. [Google Scholar] [CrossRef]
- Bigagli, E.; Lodovici, M. Circulating oxidative stress biomarkers in clinical studies on type 2 diabetes and its complications. Oxid. Med. Cell. Longev. 2019, 2019, 5953685. [Google Scholar] [CrossRef]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Du, X.; Edelstein, D.; Rossetti, L.; Fantus, I.; Goldberg, H.; Ziyadeh, F.; Wu, J.; Brownlee, M. Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. USA 2000, 9, 12222–12226. [Google Scholar] [CrossRef]
- Chung, S.; Ho, E.; Lam, K.; Chung, S. Contribution of polyol pathway to diabetes-induced oxidative stress. J. Am. Soc. Nephrol. 2003, 14, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Araki, N.; Koh, N.; Nakamura, J.; Horiuchi, S.; Hotta, N. Rapid formation of advanced glycation end products by intermediate metabolites of glycolytic pathway and polyol pathway. Biochem. Biophys. Res. Commun. 1996, 228, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Martin, K.A.; Hwa, J. Aldose reductase, oxidative stress, and diabetic mellitus. Front. Pharmacol. 2012, 3, 87. [Google Scholar] [CrossRef] [PubMed]
- Monnier, V.M.; Nagaraj, R.H.; Portero-Otin, M.; Glomb, M.; Elgawish, A.H.; Sell, D.R.; Friedlander, M.A. Structure of advanced Maillard reaction products and their pathological role. Nephrol. Dial. Transplant. 1996, 11, 20–26. [Google Scholar] [CrossRef]
- Mohamed, A.; Bierhaus, A.; Schiekofer, S.; Tritschler, H.; Ziegler, R.; Nawroth, P. The role of oxidative stress and NF-κB activation in late diabetic complications. BioFactors 1999, 10, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Hori, O.; Brett, J.; Yan, S.; Wautier, J.; Stern, D. Cellular receptors for advanced glycation end products. Implications for induction of oxidant stress and cellular dysfunction in the pathogenesis of vascular lesions. Arterioscler. Thromb. J. Vasc. Biol. 1994, 10, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Bali, A.; Singh, N.; Jaggi, A. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef]
- Xia, P.; Inoguchi, T.; Kern, T.S.; Engerman, R.L.; Oates, P.J.; King, G.L. Characterization of the mechanism for the chronic activation of diacylglycerol-protein kinase C pathway in diabetes and hypergalactosemia. Diabetes 1994, 43, 1122–1129. [Google Scholar] [CrossRef]
- Newton, A.C. Regulation of the ABC kinases by phosphorylation: Protein kinase C as a paradigm. Biochem. J. 2003, 370, 361–371. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diab. 2015, 6, 456. [Google Scholar] [CrossRef]
- Wells, L.; Vosseller, K.; Hart, G.W. Glycosylation of nucleocytoplasmic proteins: Signal transduction and O-GlcNAc. Science 2001, 291, 2376–2378. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Noble, N.A.; Cohen, A.H.; Nast, C.C.; Hishida, A.; Gold, L.I.; Border, W.A. Expression of transforming growth factor-β isoforms in human glomerular diseases. Kidney Int. 1996, 49, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Al-Jiffri, E. Association between adipocytokines, systemic inflammation and oxidative stress biomarkers among obese type 2 diabetic patients. Adv. Res. Gastroenterol. Hepatol. 2017, 5, 80–85. [Google Scholar] [CrossRef]
- Aouacheri, O.; Saka, S.; Krim, M.; Messaadia, A.; Maidi, I. The investigation of the oxidative stress-related parameters in type 2 diabetes mellitus. Can. J. Diabetes 2015, 39, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Lasisi, I.A.; Adedokun, K.A.; Oyenike, M.A.; Muhibi, M.A.; Kamorudeen, R.T.; Oluogun, W.A. Glycemic control and its impact on oxidative stress biomarkers in type 2 diabetic patients treated with metformin: A cross-sectional analysis. Sci. Med. 2019, 29, e33630. [Google Scholar] [CrossRef]
- Ozmutlu, A.; Petcu, L.; Ştefan, S.; Mitu, M.; Lixandru, D.; Ionescu-Tîrgovişte, C.; Pîrcălăbioru, G.G.; Ciulu-Costinescu, F.; Bubulica, M.V.; Chifiriuc, M.C. Markers of oxidative stress and antioxidant defense in romanian patients with type 2 diabetes mellitus and obesity. Molecules 2017, 22, 714. [Google Scholar]
- Whiting, P.; Kalansooriya, A.; Holbrook, I.; Haddad, F.; Jennings, P. The relationship between chronic glycaemic control and oxidative stress in type 2 diabetes mellitus. Br. J. Biomed. Sci. 2008, 65, 71–74. [Google Scholar] [CrossRef]
- Mandal, M.; Varghese, A.; Gaviraju, V.; Talwar, S.; Malini, S. Impact of hyperglycaemia on molecular markers of oxidative stress and antioxidants in type 2 diabetes mellitus. Clin. Diabetol. 2019, 8, 215–222. [Google Scholar] [CrossRef]
- George, G.; Ganjifrockwala, F.A.; Joseph, J.T. Biomarkers of oxidative stress and antioxidant status in type 2 diabetes patients-a study among African diabetic patients on treatment. Diabetes Obes. Metab. 2015, 6, 39. [Google Scholar]
- Casoinic, F.; Sampelean, D.; Buzoianu, A.D.; Hancu, N.; Baston, D. Serum levels of oxidative stress markers in patients with type 2 diabetes mellitus and non-alcoholic steatohepatitis. Rom. J. Intern. Med. 2016, 54, 228–236. [Google Scholar] [CrossRef]
- Čolak, E.; Majkić-Singh, N.; Stanković, S.; Srecković-Diimitrijević, V.; Djordjević, P.; Lalić, K.; Lalić, N. Parameters of antioxidative defense in type 2 diabetic patients with cardiovascular complications. Ann. Med. 2005, 37, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.M.; Paiva, S.A.R. β-carotene and other carotenoids as antioxidants. J. Am. Coll. Nutr. 1999, 8, 426–433. [Google Scholar]
- Gerster, H. The potential role of lycopene for human health. J. Am. Coll. Nutr. 1997, 16, 109–126. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Use of lycopene as a food colour. EFSA J. 2008, 674, 1–66. [Google Scholar]
- Zechmeister, L. Isomerization and stereochemistry. Chem. Rev. 1994, 34, 267–344. [Google Scholar] [CrossRef]
- Klipstein-Grobusch, K.; Launer, L.; Geleijnse, J.; Boeing, H.; Hofman, A.; Witteman, J. Serum carotenoids and atherosclerosis. Atherosclerosis 2000, 148, 49–56. [Google Scholar] [CrossRef]
- Shi, J.; Le Maguer, M. Lycopene in tomatoes: Chemical and physical properties affected by food processing. Crit. Rev. Biotechnol. 2000, 20, 293–334. [Google Scholar] [CrossRef]
- Agarwal, A.; Shen, H.; Agarwal, S.; Rao, A.V. Lycopene content of tomato products: Its stability, bioavailability and in vivo antioxidant properties. J. Med. Food 2001, 4, 9–15. [Google Scholar] [CrossRef]
- Chasse, G.; Mak, M.; Deretey, E.; Farkas, I.; Torday, L.; Papp, J.; Sarma, D.; Argawal, A.; Chakaravarthi, S.; Argawal, S.; et al. An ab initio computational study on selected lycopene isomers. J. Mol. Struct. 2001, 571, 27–37. [Google Scholar] [CrossRef]
- Rao, A.V.; Agarwal, S. Role of antioxidant lycopene in cancer and heart disease. J. Am. Coll. Nutr. 2000, 19, 563–569. [Google Scholar] [CrossRef]
- Roldán-Gutiérrez, J.; Luque de Castro, M.D. Lycopene: The need for better methods for characterization and determination. Trac. Trends Anal. Chem. 2007, 26, 163–170. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Revised exposure assessment for lycopene as a food colour on request from the European Commission. EFSA J. 2010, 8, 1444. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Shixian, Q.; Dai, Y.; Kakuda, Y.; Shi, J.; Mittal, G.; Yeung, D.; Jiang, Y. Synergistic anti-oxidative effects of lycopene with other bioactive compounds. Food Rev. Int. 2005, 21, 295–311. [Google Scholar] [CrossRef]
- Preedy, V.; Watson, R. Tomatoes and Tomato Products; Enfield, N.H., Ed.; The Science Publishers: Rawalpindi, Pakistan, 2008. [Google Scholar]
- Journal, T.E. Opinion of the Scientific Panel on food additives, flavourings, processing aids and materials in contact with food (AFC) related to an application on the use of alpha-tocopherol containing oil suspensions and cold water dispersible forms of lycopene from. EFSA J. 2005, 3, 275. [Google Scholar]
- Mantzouridou, F.; Tsimidou, M.Z. Lycopene formation in Blakeslea trispora. Chemical aspects of a bioprocess. Trends Food Sci. Technol. 2008, 19, 363–371. [Google Scholar] [CrossRef]
- Pegklidou, K.; Mantzouridou, F.; Tsimidou, M.Z. Lycopene production using blakeslea trispora in the presence of 2-methyl imidazole: Yield, selectivity, and safety aspects. J. Agr. Food Chem. 2008, 56, 4482–4490. [Google Scholar] [CrossRef]
- Matulka, R.A.; Hood, A.M.; Griffiths, J.C. Safety evaluation of a natural tomato oleoresin extract derived from food-processing tomatoes. Regul. Toxicol. Pharmacol. 2004, 39, 390–402. [Google Scholar] [CrossRef]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef]
- Shi, J.; Le Maguer, M.; Bryan, M. Lycopene from Tomatoes. In Functional Foods-Biochemical and Processing Aspects; Shi, J., Mazza, G., Le Maguer, M., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 135–168. [Google Scholar]
- Olempska-Beer, Z. 67th JECFA-Chemical and technical assessment (CTA). Assess. Technol. Eur. W. 2006, 1, 1–21. [Google Scholar]
- Cantrell, A.; McGarvey, D.J.; Truscott, T.G.; Rancan, F.; Böhm, F. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch. Biochem. Biophys. 2003, 412, 47–54. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Liu, Y.; Lv, Q.Z. DFT study on the quenching mechanism of singlet oxygen by lycopene. RSC Adv. 2016, 98498–98505. [Google Scholar] [CrossRef]
- Tinkler, J.; Böhm, F.; Schalch, W.; Truscott, T. Dietary carotenoids protect human cells from damage. J. Photochem. Photobiol. B Biol. 1994, 26, 283–285. [Google Scholar] [CrossRef]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids that Exhibit both Anti- and Pro-Oxidative Mechanisms-A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef]
- Burton, G.W.; Ingold, K.U. β-Carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Lowe, G. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Conn, P.F.; Lambert, C.; Land, E.J.; Schalch, W.; Truscott, T.G. Carotene-oxygen radical interactions. Free Radic. Res. 1992, 16, 401–408. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Yeum, K.J. Carotenoid-radical interactions. Biochem. Biophys. Res. Commun. 2003, 305, 754–760. [Google Scholar] [CrossRef]
- Rao, A.V.; Shen, H. Effect of low dose lycopene intake on lycopene bioavailability and oxidative stress. Nutr. Res. 2002, 22, 1125–1131. [Google Scholar] [CrossRef]
- Fuhrman, B.; Volkova, N.; Rosenblat, M.; Aviram, M. Lycopene synergistically inhibits ldl oxidation in combination with vitamin E, glabridin, rosmarinic acid, carnosic acid, or garlic. Antioxid. Redox Signal. 2000, 2, 491–506. [Google Scholar] [CrossRef]
- Truscott, T. β-carotene and disease: A suggested pro-oxidant and anti-oxidant mechanism and speculations concerning its role in cigarette smoking. J. Photochem. Photobiol. B Biol. 1996, 35, 233–235. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Clevidence, B.; Bieri, J. Association of carotenoids with human plasma lipoproteins. Methods Enzymol. 1993, 214, 33–46. [Google Scholar] [PubMed]
- Al-Delaimy, W.K.; van Kappel, A.L.; Ferrari, P.; Slimani, N.; Steghens, J.P.; Bingham, S.; Johansson, I.; Wallström, P.; Overvad, K.; Tjønneland, A.; et al. Plasma levels of six carotenoids in nine European countries: Report from the European Prospective Investigation into Cancer and Nutrition (EPIC). Public Health Nutr. 2004, 7, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Imrhan, V. Tomatoes versus lycopene in oxidative stress and carcinogenesis: Conclusions from clinical trials. Eur. J. Clin. Nutr. 2007, 61, 295–303. [Google Scholar] [CrossRef]
- Quansah, D.Y.; Ha, K.; Jun, S.; Kim, S.A.; Shin, S.; Wie, G.A.; Joung, H. Associations of dietary antioxidants and risk of type 2 diabetes: Data from the 2007–2012 Korea national health and nutrition examination survey. Molecules 2017, 22, 1664. [Google Scholar] [CrossRef]
- Li, W.; Wang, G.; Lu, X.; Jiang, Y.; Xu, L.; Zhao, X. Lycopene ameliorates renal function in rats with streptozotocin-induced diabetes. Int. J. Clin. Exp. Pathol. 2014, 7, 5008–5015. [Google Scholar]
- Li, Z.; Lu, X.Z.; Ma, C.C.; Chen, L. Serum lycopene in diabetic retinopathy. Eur. J. Ophthalmol. 2010, 20, 719–723. [Google Scholar] [CrossRef]
- Assis, R.P.; Arcaro, C.A.; Gutierres, V.O.; Oliveira, J.O.; Costa, P.I.; Baviera, A.M.; Brunetti, I.L. Combined effects of curcumin and lycopene or bixin in yoghurt on inhibition of LDL oxidation and increases in HDL and paraoxonase levels in streptozotocin-diabetic rats. Int. J. Mol. Sci. 2017, 18, 322. [Google Scholar] [CrossRef]
- Brazionis, L.; Rowley, K.; Itsiopoulos, C.; O’dea, K. Plasma carotenoids and diabetic retinopathy. Br. J. Nutr. 2009, 101, 270–277. [Google Scholar] [CrossRef]
- Ford, E.S.; Mokdad, A.H.; Giles, W.H.; Brown, D.W. The metabolic syndrome and antioxidant concentrations: Findings from the Third National Health and Nutrition Examination Survey. Diabetes 2003, 52, 2346–2352. [Google Scholar] [CrossRef] [PubMed]
- Polidori, M.; Mecocci, P.; Stahl, W.; Parente, B.; Cecchetti, R.; Cherubini, A.; Cao, P.; Sies, H.; Senin, U. Plasma levels of lipophilic antioxidants in very old patients with Type 2 diabetes. Diabetes Metab. Res. Rev. 2000, 16, 15–19. [Google Scholar] [CrossRef]
- Malekiyan, R.; Abdanipour, A.; Sohrabi, D.; Jafari Anarkooli, I. Antioxidant and neuroprotective effects of lycopene and insulin in the hippocampus of streptozotocin-induced diabetic rats. Biomed. Rep. 2019, 10, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Akinnuga, A.M.; Bamidele, O.; Ebunlomo, O.A.; Adeniyi, O.S.; Adeleyea, G.S.; Ebomuche, L.C. Hypoglycaemic effects of dietary intake of ripe and unripe Lycopersicon esculentum (Tomatoes) on streptozotocin-induced diabetes mellitus in rats. Online J. Biol. Sci. 2010, 10, 50–53. [Google Scholar] [CrossRef][Green Version]
- Sharma, P.K.; Saxena, P.; Jaswanth, A.; Chalamaiah, M.; Balasubramaniam, A. Anti- diabetic activity of lycopene niosomes: Experimental observation. J. Pharm. Drug Dev. 2017, 4, 103. [Google Scholar]
- Saad, A.; Ahmad, W.; Haque, A.E.; Al-Jassabi, S. The role of lycopene from zizyphus spina-christi in the prevention of streptozotocin-induced diabetes mellitus in balb/c mice. Middle-E. J. Sci. Res. 2013, 16, 952–956. [Google Scholar]
- Ozmen, O.; Topsakal, S.; Haligur, M.; Aydogan, A.; Dincoglu, D. Effects of caffeine and lycopene in experimentally induced diabetes mellitus. Pancreas 2016, 45, 579–583. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, C.; Xu, Y. Lycopene attenuates endothelial dysfunction in streptozotocin-induced diabetic rats by reducing oxidative stress. Pharm. Biol. 2011, 49, 1144–1149. [Google Scholar] [CrossRef]
- Baş, H.; Pandir, D.; Kalender, S. Furan-induced hepatotoxic and Hematologic changes in diabetic rats: The protective role of lycopene. Arh. Hig. Rada Toksikol. 2016, 67, 194–203. [Google Scholar] [CrossRef]
- Duzguner, V.; Kucukgul, A.; Erdogan, S.; Celik, S.; Sahin, K. Effect of lycopene administration on plasma glucose, oxidative stress and body weight in streptozotocin diabetic rats. J. Appl. Anim. Res. 2008, 33, 17–20. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Zhang, H.; He, X.; Bai, A. Lycopene ameliorates erectile dysfunction in streptozotocin-induced diabetic rats. Pharmazie 2012, 67, 256–259. [Google Scholar] [PubMed]
- Bayramoglu, A.; Bayramoglu, G.; Senturk, H. Lycopene partially reverses symptoms of diabetes in rats with streptozotocin-induced diabetes. J. Med. Food 2013, 16, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Eze, E.D.; Tanko, Y.; Abubakar, A.; Sulaiman, S.O.; Rabiu, K.M.; Mohammed, A. Lycopene Ameliorates Diabetic-Induced Changes in Erythrocyte Osmotic Fragility and Lipid Peroxidation in Wistar Rats. J. Diab. Mell. 2017, 7, 71–85. [Google Scholar] [CrossRef]
- Yin, Y.; Zheng, Z.; Jiang, Z. Effects of lycopene on metabolism of glycolipid in type 2 diabetic rats. Biomed. Pharmacother. 2019, 109, 2070–2077. [Google Scholar] [CrossRef]
- Pollack, A.; Oren, P.; Stark, A.H.; Eisner, Z.; Nyska, A.; Madar, Z. Cataract development in sand and galactosemic rats fed a natural tomato extract. J. Agric. Food Chem. 1999, 47, 5122–5126. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Wang, Y. Beneficial effect of lycopene on anti-diabetic nephropathy through diminishing inflammatory response and oxidative stress. Food Funct. 2015, 6, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Uçar, S.; Pandir, D. Furan induced ovarian damage in non-diabetic and diabetic rats and cellular protective role of lycopene. Arch. Gynecol. Obstet. 2017, 296, 1027–1037. [Google Scholar] [CrossRef]
- Zheng, Z.; Yin, Y.; Lu, R.; Jiang, Z. Lycopene ameliorated oxidative stress and inflammation in type 2 diabetic rats. J. Food Sci. 2019, 84, 1194–1200. [Google Scholar] [CrossRef]
- Coyne, T.; Ibiebele, T.I.; Baade, P.D.; Dobson, A.; McClintock, C.; Dunn, S.; Leonard, D.; Shaw, J. Diabetes mellitus and serum carotenoids: Findings of a population-based study in Queensland, Australia. Am. J. Clin. Nutr. 2005, 82, 685–693. [Google Scholar] [CrossRef]
- She, C.; Shang, F.; Zhou, K.; Liu, N. Serum Carotenoids and Risks of Diabetes and Diabetic Retinopathy in a Chinese Population Sample. Curr. Mol. Med. 2017, 7, 287–297. [Google Scholar] [CrossRef]
- Bose, K.; Agrawal, B. Effect of long term supplementation of tomatoes (cooked) on levels of antioxidant enzymes, lipid peroxidation rate, lipid profile and glycated haemoglobin in Type 2 diabetes mellitus. W. Indian Med. J. 2006, 55, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Leh, H.E.; Lee, L.K. The role of lycopene for the amelioration of glycaemic status and peripheral antioxidant capacity among the Type II diabetes mellitus patients: A case–control study. Ann. Med. 2021, 53, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Sluijs, I.; Cadier, E.; Beulens, J.; van der A, D.; Spijkerman, A.; van der Schouw, Y. Dietary intake of carotenoids and risk of type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2015, 254, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Consitt, L.A.; Bell, J.A.; Houmard, J.A. Intramuscular lipid metabolism, insulin action, and obesity. IUBMB Life 2009, 61, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.C.; Mu, G.P.; Huang, S.F.; Zeng, X.H.; Cheng, H.; Li, Z.X. Effects of lycopene on number and function of human peripheral blood endothelial progenitor cells cultivated with high glucose. Nutr. Res. Pract. 2014, 8, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.C.; Peng, L.S.; Zou, L.; Huang, S.F.; Xie, Y.; Mu, G.P.; Zeng, X.H.; Zhou, X.L.; Zeng, Y.C. Protective effect and mechanism of lycopene on endothelial progenitor cells (EPCs) from type 2 diabetes mellitus rats. Biomed. Pharmacother. 2017, 92, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Icel, E.; Icel, A.; Uçak, T.; Karakurt, Y.; Elpeze, B.; Keskin Çimen, F.; Süleyman, H. The effects of lycopene on alloxan induced diabetic optic neuropathy. Cutan. Ocul. Toxicol. 2019, 38, 88–92. [Google Scholar] [CrossRef]
- Bazyel, B.; Dede, S.; Cetin, S.; Yuksek, V. In vitro evaluation of the effects of lycopene on caspase system and oxidative DNA damage in high-glucose condition. Pharmacogn. Mag. 2019, 15, 30–33. [Google Scholar]
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef]


{kind=link}
| No. | Study Population | Sample | Biomarkers | Observations | Reference |
|---|---|---|---|---|---|
| 1 | T2DM with CVD (n = 69) T2DM without CVD (n = 48) Control (n = 42) | erythrocyte | GPx SOD | ↓ Decreased GPx and SOD in T2DM with CVD | [42] |
| 2 | T2DM (n = 20) Control (n = 20) | plasma and erythrocyte | GPx | ↓ Decreased erythrocyte-GPx and plasma-GPx in T2DM | [38] |
| 3 | T2DM (n = 57) Control (n = 41) | serum | SOD TBARS | ↑ Increased TBARS in T2DM No differences in SOD between T2DM and control | [40] |
| 4 | T2DM (n = 59) Control (n = 48) | serum | GPx SOD MDA | ↓ Decreased GPx in T2DM ↑ Increased SOD in T2DM ↑ Increased MDA in T2DM | [35] |
| 5 | T2DM and NASH (n = 60) T2DM without NASH (n = 55) Control (n = 50) | serum | MDA | ↑ Higher MDA in T2DM and NASH | [41] |
| 6 | T2DM (n = 115) Control (n = 32) | plasma | GPx | ↓ Lower GPx level in T2DM | [37] |
| 7 | T2DM (n = 100) Control (n = 100) | serum | GPx SOD MDA | ↓ Decreased GPx and SOD level in T2DM ↑ Increased MDA in T2DM | [34] |
| 8 | T2DM (n = 83) Control (n = 81) | serum | SOD MDA | ↓ Lower SOD level in T2DM ↑ Higher MDA level in T2DM | [39] |
| 9 | Controlled-T2DM (n = 80) Uncontrolled T2DM (n = 80) Control (n = 100) | serum | GPx MDA | ↓ Reduced GPx in controlled and uncontrolled T2DM; ↑ Increased MDA in controlled and uncontrolled T2DM | [36] |
| Source of Lycopene | Average (mg/day) | High (mg/day) | Reference |
|---|---|---|---|
| Naturally occurring | 0.5–5 | 8–20 | [57] |
| Fortified foods | 8–19 | 23–37 | [45] |
| Supplements | 0 (no supplement use) | 8–15 | [45] |
| Food color | 2–6 | 11–23 | [57] |
| Compound a | Number of Conjugated Carbon-Carbon Double Bonds b | Terminal Rings | Quenching Rate Constant, (kq L mol−1 s−1) | Relative Rates |
|---|---|---|---|---|
| Lycopene | 11 | 0 | 3.1 × 1010 | 103 |
| γ-Carotene | 11 | 1 | 2.5 × 1010 | 83 |
| Astaxanthin | 11(2) | 2 | 2.4 × 1010 | 80 |
| Canthaxanthin | 11(2) | 2 | 2.1 × 1010 | 70 |
| α-Carotene | 10 | 2 | 1.9 × 1010 | 63 |
| Bixin | 9(2) | 0 | 1.4 × 1010 | 47 |
| β-Carotene | 11 | 2 | 1.4 × 1010 | 47 |
| Zeaxanthin | 11 | 2 | 1.0 × 1010 | 33 |
| Lutein | 10 | 2 | 0.8 × 1010 | 27 |
| Cryptoxanthin | 11 | 2 | 0.6 × 1010 | 20 |
| Crocin | 7(2) | 0 | 0.11 × 1010 | 3.7 |
| α-Tocopherol | n.c.c | n.c. | 0.03 × 1010 | 1 |
| Lipoic acid | n.c. | n.c. | 0.0138 × 1010 | 0.46 |
| Glutathione | n.c. | n.c. | 0.00024 × 1010 | 0.008 |
| No. | Population | Treatment/Method | Effects | Reference |
|---|---|---|---|---|
| 1 | STZ-induced diabetic Wistar-Albino male rats | Oral administration of 10 mg/kg/day lycopene in corn oil for 28 days | Reduction of blood glucose level and HbA1c % | [6] |
| 2 | STZ-induced diabetic Wistar-Albino Rats | Administration of 10/mg/kg/day of lycopene for 28 days | Reduction in HbA1c % | [7] |
| 3 | STZ-induced diabetic mice | Lycopene supplementation (40, 80 mg) | Decreased serum blood glucose concentration | [80] |
| 4 | STZ-induced Albino Wistar rats | Supplementation of ripe and unripe tomato (10%) for 14 days | Increased insulin level (37%) Decreased glucose concentration (33%) | [86] |
| 5 | STZ-induced diabetic rats | Single dose of 90 mg/kg/bw tomato-extract lycopene | Decreased FBG levels | [87] |
| 6 | Wistar Rats | Lycopene niosomes (100 and 200 mg/kg/bw for 14 days) | Decreased FBG levels | [88] |
| 7 | STZ-induced diabetic male Balb/c mice | Oral administration of lycopene dose (100, 200 mg/kg/bw) for 10 days | Decreased in FBG levels; No changes in serum insulin levels | [89] |
| 8 | STZ-induced diabetic rats | Lycopene + caffeine administration by oral gavages for 1 month | Decreased blood and urine FBG levels; Increased serum insulin levels | [90] |
| 9 | STZ-induced diabetic rats | Administration of lycopene (10, 30, 60 mg/kg/bw) for 30 days | Decreased FBG levels | [91] |
| 10 | STZ-induced diabetic rats | Administration of 4 mg/kg/bw of lycopene | No significant changes in HbA1c levels; Decreased in FBG levels | [92] |
| 11 | STZ-induced diabetic rats | Oral administration of 10 mg/kg/bw lycopene for 3 weeks | Reduction of blood glucose levels by 25% | [93] |
| 12 | STZ-induced male Sprague-Dawley rats | Administration of lycopene (10, 30, 60 mg/kg/d) for 8 weeks | Decreased FBG levels | [94] |
| 13 | STZ-induced diabetic rats | Administration of lycopene at the dose of 2.5 mg/kg/bw for 7 days | Reduction in serum glucose levels; Increased serum insulin levels | [95] |
| 14 | STZ-induced diabetic Wistar rats | Oral administration of lycopene in sunflower oil at a dose of 4mg/kg/bw for 8 weeks | Decreased FBG levels; Increased plasma insulin concentration | [96] |
| 15 | STZ-induced diabetic Wistar Rats | Oral administration of lycopene (10, 20 and 40 mg/kg/bw) for 4 weeks | Decreased FBG levels | [97] |
| 16 | STZ-induced diabetic rats | Oral administration of lycopene oil solution (10 mg/kg or 20 mg/kg/bw) for 10 weeks | Decreased FBG levels | [98] |
| No. | Population | Treatment | Effects on Oxidative Stress Biomarkers | Reference |
|---|---|---|---|---|
| 1 | 24 STZ-induced diabetic Wistar rats | Oral administration of lycopene in sunflower oil at a dose of 4 mg/kg/bw for 8 weeks | Increased SOD, CAT and GPx activities in erythrocytes Decreased GSH and NO levels (plasma) and GSH levels (brain tissue); Decreased in brain tissue MDA levels but no significant effect in plasma MDA levels | [6] |
| 2 | 40 STZ-induced diabetic male Balb/c mice | Oral administration of lycopene (100, 200 mg/kg/bwt) for 10 days | Decreased in ROS levels in serum, liver and pancreas tissues; Decreased in SOD, CAT, and GPx; Prevent increase in LPO level (liver, pancreas) | [7] |
| 3 | STZ-induced rats | Single dose of 90 mg/kg/bw of tomato-extract lycopene | Decreased H2O2 formation; Increased CAT, SOD and GPx | [78] |
| 4 | STZ-induced diabetic Wistar Rats | Oral administration of 90 mg/kg curcumin + 45 mg/kg lycopene in yogurt | Increased CAT levels; Decreased serum Ox-LDL and liver TBARS | [82] |
| 5 | STZ-induced diabetic rats | Single dose of 90 mg/kg/bw tomato-extract lycopene | Increased CAT, SOD and GPx | [87] |
| 6 | STZ-induced diabetic rats | Administration of lycopene (10, 30, 60 mg/kg) for 30 days | Increased aortic SOD activity; Decreased MDA levels | [91] |
| 7 | STZ-induced male diabetic rats | Administration of 4 mg kg−1 bw lycopene for 28 days | Increased GPx, SOD, CAT and GST levels in liver tissue Decreased MDA level in liver tissue | [92] |
| 8 | STZ-induced diabetic rats | Oral administration of 10mg/kg/bw lycopene for 3 weeks | Reduction of LPO rate and NO in the plasma | [93] |
| 9 | STZ-induced male Sprague–Dawley rats | Administration of lycopene (10, 30, 60 mg/kg/d) for 8 weeks | Decreased MDA levels and increased SOD activities | [94] |
| 10 | 60 STZ-induced diabetic male Sprague–Dawley rats | Administration of 20 mg/kg/day lycopene by oral gavage tube for 8 weeks | Increased SOD activity; Decreased kidney MDA levels | [95] |
| 11 | STZ-induced diabetic nephropathy mice | Lycopene supplementation (40, 80 mg) | Augmented bioactivities of SOD, GPx; Reduction of MDA level | [96] |
| 12 | STZ-induced diabetic Wistar Rats | Oral administration of lycopene oil solution (10 mg/kg or 20 mg/kg/bw) for 10 weeks | Increased GPx and SOD Decreased MDA level in pancreas | [97] |
| 13 | Diabetic sand rats | Administration of natural tomato extract at 0.2% in the diet for 5 weeks | Increased GSH levels | [98] |
| 14 | STZ-induced diabetic Wistar Rats | Oral administration of graded dose of lycopene (10, 20 and 40 mg/kg bw) for 4 weeks | Decreased erythrocyte MDA concentration | [99] |
| 15 | STZ-induce female Wistar-Albino diabetic rats | Administration of 4 mg/kg/bw lycopene for 28 days | Increased CAT, SOD, GPx and GST enzymes activities; Decreased MDA level | [100] |
| 16 | STZ-induced diabetic Wistar Rats | Oral administration of lycopene dose (0, 5, 10 and 15 mg/kg/bw) for 10 weeks | Increased CAT, SOD and GPx Decreased MDA level in pancreas | [101] |
| No. | Study Design | Subjects Characteristics | Intervention | Effects on T2DM | References |
|---|---|---|---|---|---|
| 1 | Randomized placebo-controlled trial | T2DM (n = 57) Mean age 63 ± 8 years | Tomato juice (500 mL/day) + vitamin E (800 U/day) and vitamin C (500 mg/day) for 4 weeks | No changes in plasma glucose concentration | [4] |
| 2 | Case-control study | T2DM (n = 71) Non-T2DM (n = 23) Age > 50 years | Dietary Intake | Negative correlation between HbA1c and serum lycopene | [81] |
| 3 | Prospective cohort study | Total, n = 1597 T2DM (n = 132) Age ≥ 25 years | Dietary Intake | Reduced plasma glucose and fasting insulin concentrations with increased serum lycopene | [102] |
| 4 | Cross-sectional study | T2DM (n = 190) T2DM + DR (n = 272) Control (n = 285) | Dietary Intake | No significant association between HbA1c and lycopene | [103] |
| 5 | Case-control study | T2DM (n = 40) Control (n = 50) Age 35–55 years | Ripe tomatoes (cooked) (200 g/day) for 30 days | No significant changes in fasting blood sugar and HbA1c levels | [104] |
| 6 | Case-control study | T2DM (n = 87) Control (n = 122) | Dietary intake | HbA1c and FBG levels decreased significantly with higher lycopene intake | [105] |
| No | Subjects Characteristics | Intervention | Effect after Lycopene Supplementation | References |
|---|---|---|---|---|
| 1 | T2DM (n = 57) >75 years | Tomato juice (500 mL/day) + vitamin E (800 U/day) and vitamin C (500 mg/day) for 4 weeks | ↓ LDL oxidation | [4] |
| 2 | T2DM (n = 35) 54 ± 9 years | Lycopene supplementation (10 mg/d) or placebo for 8 weeks | ↑ TAC levels; Inhibit MDA-LDL formation | [5] |
| 3 | T2DM (n = 4304) 40–69 years | Dietary intake | No association between lycopene and risk of T2DM | [11] |
| 4 | Total (n = 35,784) ≥45 years | 131-item-validated semi-quantitative FFQ | No association between either dietary lycopene or lycopene-containing foods and the risk of T2DM | [13] |
| 5 | T2DM (n = 50) T2DM + Lycopene (n = 50) Control (n = 50) 48 ± 6 years | Oral administration of lycopene (4 mg once daily for 3 months) | ↑ SOD, GPx, GR and GSH levels in T2DM + lycopene ↓ MDA and XOD in T2DM + lycopene | [14] |
| 6 | T2DM (n = 603) Control (n = 23,774) 19–74 years | 24-h dietary recall | No association between lycopene intake and reduced risk of T2DM | [79] |
| 7 | T2DM (n = 40) Control (n = 50) 35–55 years | Supplementation with cooked tomato, 200 g for 30 days | ↑ SOD, GPx, GR, GSH ↓ MDA | [104] |
| 8 | Total (n = 37,846) 49.1 years | Validated FFQ | No association between lycopene intake and reduced risk of T2DM | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leh, H.E.; Lee, L.K. Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus. Molecules 2022, 27, 2335. https://doi.org/10.3390/molecules27072335
Leh HE, Lee LK. Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus. Molecules. 2022; 27(7):2335. https://doi.org/10.3390/molecules27072335
Chicago/Turabian StyleLeh, Hui Eng, and Lai Kuan Lee. 2022. "Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus" Molecules 27, no. 7: 2335. https://doi.org/10.3390/molecules27072335
APA StyleLeh, H. E., & Lee, L. K. (2022). Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus. Molecules, 27(7), 2335. https://doi.org/10.3390/molecules27072335