
TNKPVI, a Putative Bioaccessible Pharmacophore of Anti-Inflammatory Potato Patatin-Derived Decapeptide DIKTNKPVIF

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
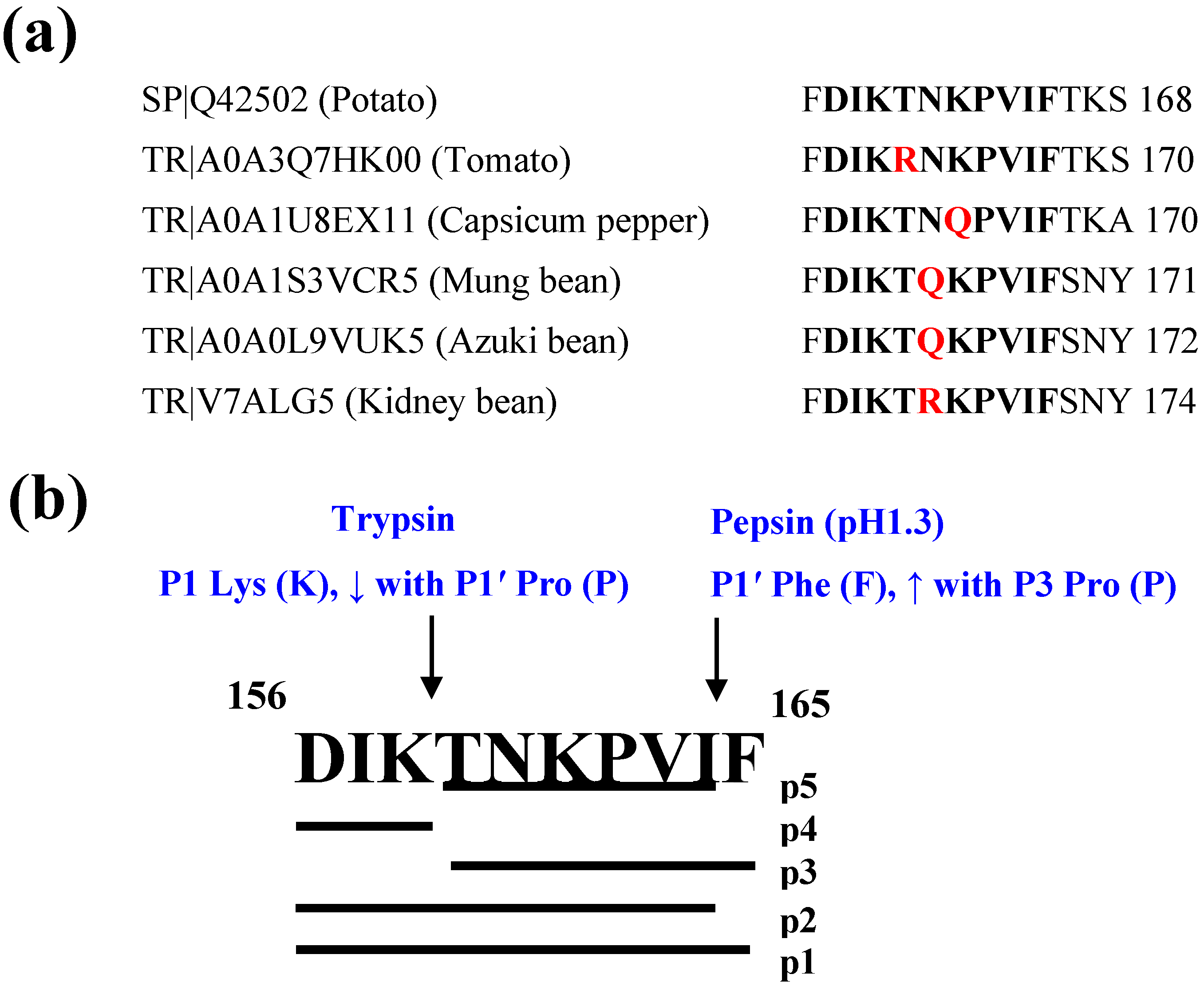
2.1. In Silico Gastrointestinal Hydrolysis
2.2. Physicochemical Properties, Pharmacokinetics and Target Predictions
2.3. Peptide Synthesis
2.4. Chemicals and Cell Culture
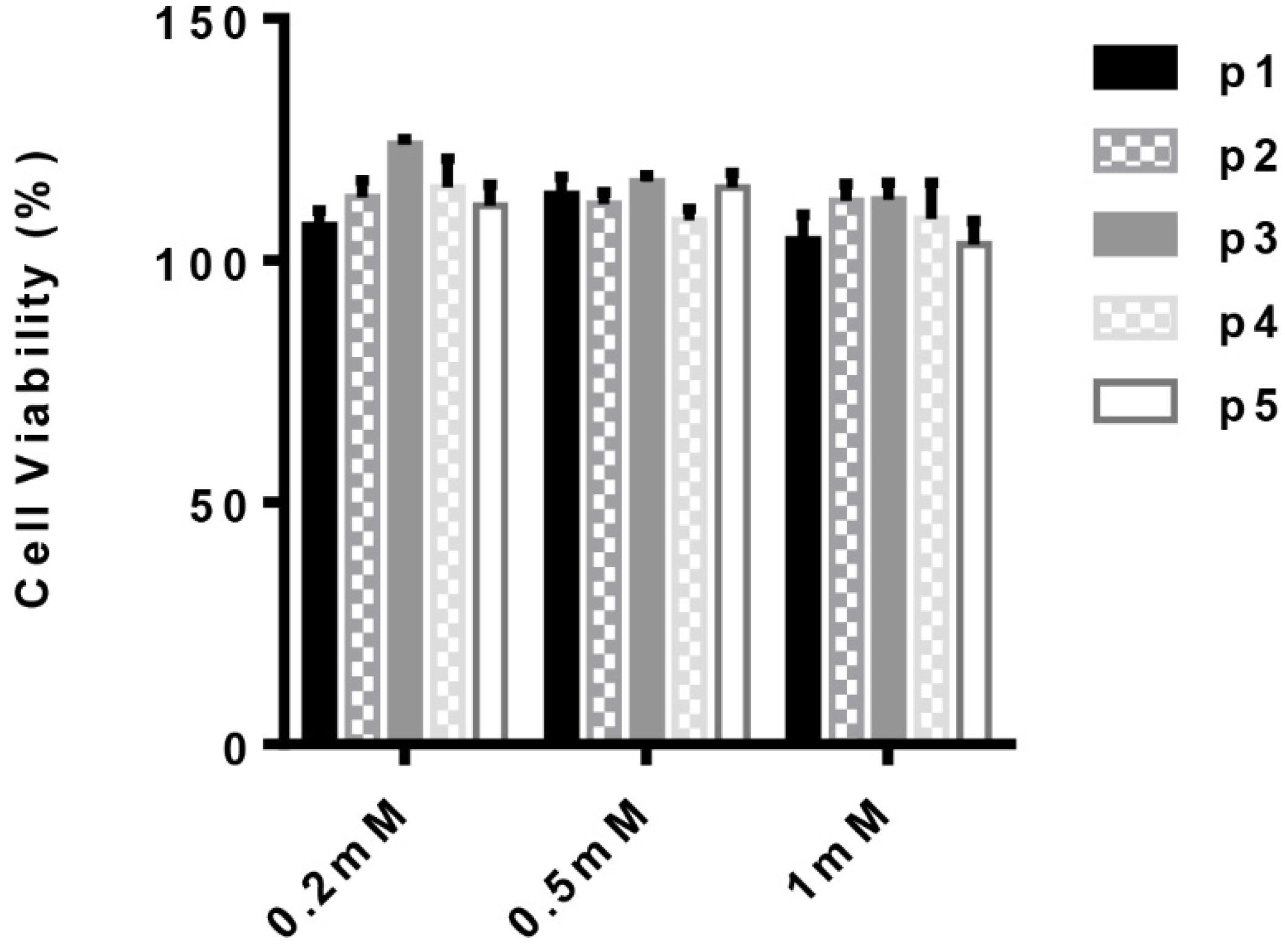
2.5. Human Monocyte-Derived Macrophages and Cytotoxicity Assay
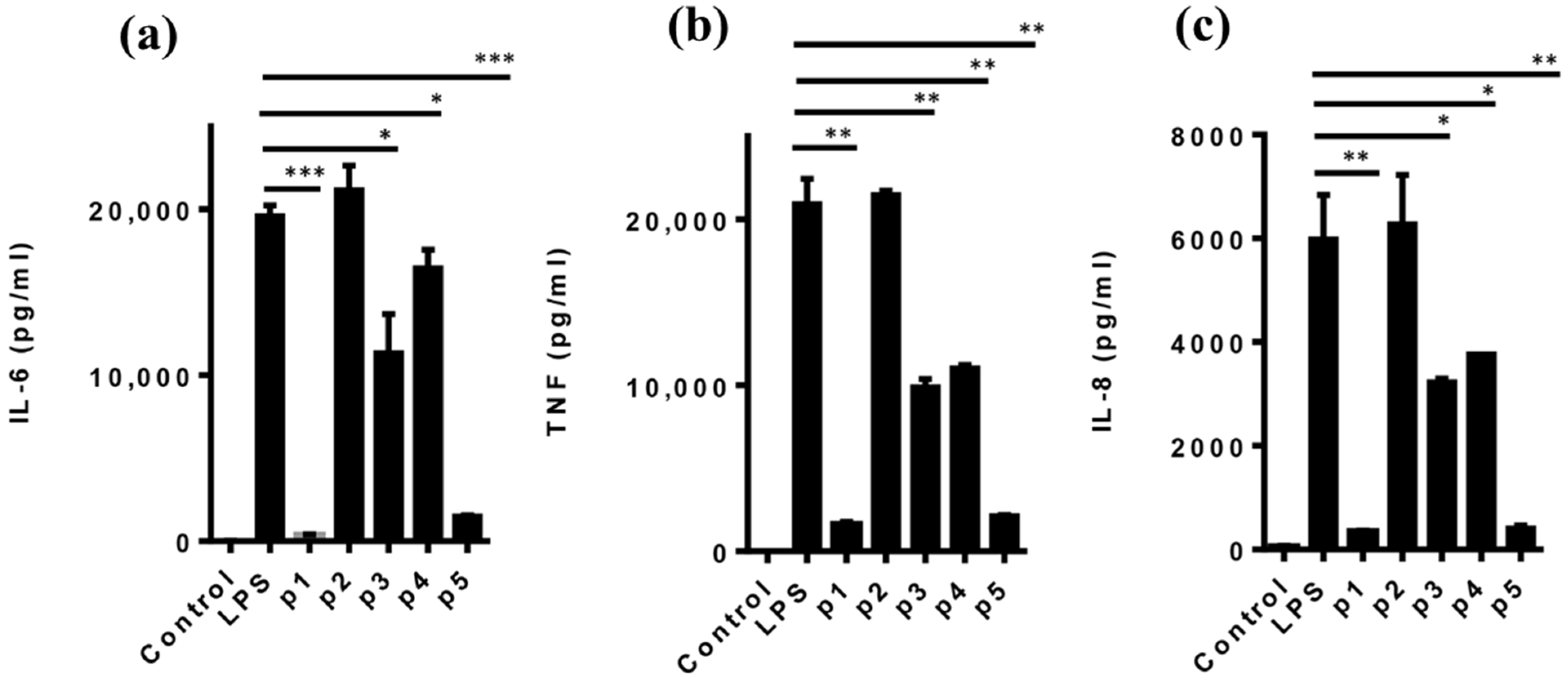
2.6. Cytokine Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. In Silico Gastrointestinal Biostability and Physicochemical Properties
3.2. Pharmacokinetic Properties of the Parent Peptide and Fragments
3.3. Anti-Inflammatory Activity of the Parent Peptide and Fragments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, A.; Rajendran, S.R.C.K.; He, Q.S.; Bazinet, L.; Udenigwe, C.C. Encapsulation of food protein hydrolysates and peptides: A review. RSC Adv. 2015, 5, 79270–79278. [Google Scholar] [CrossRef]
- Rafferty, J.; Nagaraj, H.P.; McCloskey, A.; Huwaitat, R.; Porter, S.; Albadr, A.; Laverty, G. Peptide therapeutics and the pharmaceutical industry: Barriers encountered translating from the laboratory to patients. Curr. Med. Chem. 2016, 29, 4231–4259. [Google Scholar] [CrossRef]
- Ahmed, T.; Sun, X.; Udenigwe, C.C. Role of structural properties of bioactive peptides in their stability during simulated gastrointestinal digestion: A systematic review. Trends Food Sci. Technol. 2022, 120, 265–273. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef]
- Huang, C.Y.; Nithiyanantham, S.; Liao, J.Y.; Lin, W.T. Bioactive peptides attenuate cardiac hypertrophy and fibrosis in spontaneously hypertensive rat hearts. J. Food Drug Anal. 2020, 28, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Dumeus, S.; Shibu, M.A.; Lin, W.T.; Wang, M.F.; Lai, C.H.; Shen, C.Y.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Bioactive peptide improves diet-induced hepatic fat deposition and hepatocyte proinflammatory response in SAMP8 ageing mice. Cell. Physiol. Biochem. 2018, 48, 1942–1952. [Google Scholar] [CrossRef]
- Liao, W.; Fan, H.; Liu, P.; Wu, J. Identification of angiotensin converting enzyme 2 (ACE2) up-regulating peptides from pea protein hydrolysate. J. Funct. Foods 2019, 60, 103395. [Google Scholar] [CrossRef]
- Daliri, E.B.M.; Lee, B.H.; Kim, J.H.; Oh, D.H. Novel angiotensin I-converting enzyme inhibitory peptides from soybean protein isolates fermented by Pediococcus pentosaceus SDL1409. LWT 2018, 93, 88–93. [Google Scholar] [CrossRef]
- Asledottir, T.; Picariello, G.; Mamone, G.; Ferranti, P.; Røseth, A.; Devold, T.; Vegarud, G. Degradation of ß-casomorphin-7 through in vitro gastrointestinal and jejunal brush border membrane digestion. J. Dairy Sci. 2019, 102, 8622–8629. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Consortium, O.S.D.D.; Raghava, G.P.S. In silico approach for predicting toxicity of peptides and proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.B.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Han, C.H.; Lee, M.H.; Hsu, F.L.; Hou, W.C. Patatin, the tuber storage protein of potato (Solanum tuberosum L.), exhibits antioxidant activity in vitro. J. Agric. Food Chem. 2003, 51, 4389–4393. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Udechukwu, M.C.; Yiridoe, C.; Gibson, A.; Gong, M. Antioxidant mechanism of potato protein hydrolysates against in vitro oxidation of reduced glutathione. J. Funct. Foods 2016, 20, 195–203. [Google Scholar] [CrossRef]
- Keil, B. Specificity of Proteolysis; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Udenigwe, C.C.; Abioye, R.O.; Okagu, I.U.; Obeme-Nmom, J.I. Bioaccessibility of bioactive peptides: Recent advances and perspectives. Curr. Opin. Food Sci. 2021, 39, 182–189. [Google Scholar] [CrossRef]
- Ao, J.; Li, B. Stability and antioxidative activities of casein peptide fractions during simulated gastrointestinal digestion in vitro: Charge properties of peptides affect digestive stability. Food Res. Int. 2013, 52, 334–341. [Google Scholar] [CrossRef]
- Boman, H.G. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef]
- Guruprasad, K.; Reddy, B.V.B.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. Des. Sel. 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ikai, A. Thermostability and aliphatic index of globular proteins. J. Biochem. 1980, 88, 1895–1898. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.B. Drugs as P-glycoprotein substrates, inhibitors, and inducers. Drug Metab. Rev. 2002, 34, 47–54. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Martin, Y.C. A bioavailability score. J. Med. Chem. 2005, 48, 3164–3170. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Delaney, J.S. ESOL: Estimating aqueous solubility directly from molecular structure. J. Chem. Inf. Comput. Sci. 2004, 44, 1000–1005. [Google Scholar] [CrossRef]
- Liu, B.; Shao, Y.; Fu, R. Current research status of HLA in immune-related diseases. Immun. Inflamm. Dis. 2021, 9, 340–350. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peptide or Fragment | MW (Da) | Hydrophobicity Index | Net Charge (pH 1.2) | Net Charge (pH 7) | Boman Index | Instability Index | Aliphatic Index |
|---|---|---|---|---|---|---|---|
| DIKTNKPVIF (p1) | 1174.4 | −0.11 | 2.99 | 0.76 | 1.22 | 20.88 | 107.0 |
| DIKTNKPVI (p2) | 1027.2 | −0.43 | 2.99 | 0.76 | 1.68 | 22.09 | 118.9 |
| TNKPVIF (p3) | 817.98 | 0.26 | 2.00 | 0.76 | 0.40 | 37.67 | 97.14 |
| DIK (p4) | 374.43 | −0.97 | 1.99 | −0.24 | 3.12 | −21.63 | 130.0 |
| TNKPVI (p5) | 670.81 | −0.17 | 2.00 | 0.76 | 0.97 | 42.28 | 113.3 |
| Peptide Sequence | Physicochemical Properties | Toxicity | Lipophilicity | Drug-Likeness | Pharmacokinetics | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROTB (n) < 10 | HBA (n) < 10 | HBD (n) < 5 | ESOL Log S | SVM Score (<0.0) | TPSA (Å2) < 140 | ClogP (o/w) < 5 | Bioavailability Score | Lipinski Filter | GIA | P-Gly Substrate | CYP3A4 Inhibitor | BBB Permeability | |
| DIKTNKPVIF | 47 | 18 | 15 | −0.58 (VS) | −1 Non-toxin | 469.09 | −1.69 | 0.17 | 3 | Low | Yes | No | No |
| DIKTNKPVI | 42 | 17 | 14 | 0.89 (HS) | −0.83 Non-toxin | 439.99 | −2.16 | 0.17 | 3 | Low | Yes | No | No |
| TNKPVIF | 30 | 12 | 10 | −1.3 (VS) | −0.92 Non-toxin | 318.47 | −0.93 | 0.17 | 3 | Low | Yes | No | No |
| DIK | 15 | 8 | 6 | 2.34 (HS) | −0.83 Non-toxin | 184.84 | −1.4 | 0.55 | 1 | Low | Yes | No | No |
| TNKPVI | 25 | 11 | 9 | 0.2 (HS) | −0.78 Non-toxin | 289.37 | −1.58 | 0.17 | 3 | Low | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okeke, E.B.; Abioye, R.O.; Ventura-Santana, E.; Sun, X.; Udenigwe, C.C. TNKPVI, a Putative Bioaccessible Pharmacophore of Anti-Inflammatory Potato Patatin-Derived Decapeptide DIKTNKPVIF. Molecules 2022, 27, 3869. https://doi.org/10.3390/molecules27123869
Okeke EB, Abioye RO, Ventura-Santana E, Sun X, Udenigwe CC. TNKPVI, a Putative Bioaccessible Pharmacophore of Anti-Inflammatory Potato Patatin-Derived Decapeptide DIKTNKPVIF. Molecules. 2022; 27(12):3869. https://doi.org/10.3390/molecules27123869
Chicago/Turabian StyleOkeke, Emeka B., Raliat O. Abioye, Esmeiry Ventura-Santana, Xiaohong Sun, and Chibuike C. Udenigwe. 2022. "TNKPVI, a Putative Bioaccessible Pharmacophore of Anti-Inflammatory Potato Patatin-Derived Decapeptide DIKTNKPVIF" Molecules 27, no. 12: 3869. https://doi.org/10.3390/molecules27123869
APA StyleOkeke, E. B., Abioye, R. O., Ventura-Santana, E., Sun, X., & Udenigwe, C. C. (2022). TNKPVI, a Putative Bioaccessible Pharmacophore of Anti-Inflammatory Potato Patatin-Derived Decapeptide DIKTNKPVIF. Molecules, 27(12), 3869. https://doi.org/10.3390/molecules27123869