
In Vitro Pharmacological Screening of Essential Oils from Baccharis parvidentata and Lippia origanoides Growing in Brazil

 ,
,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
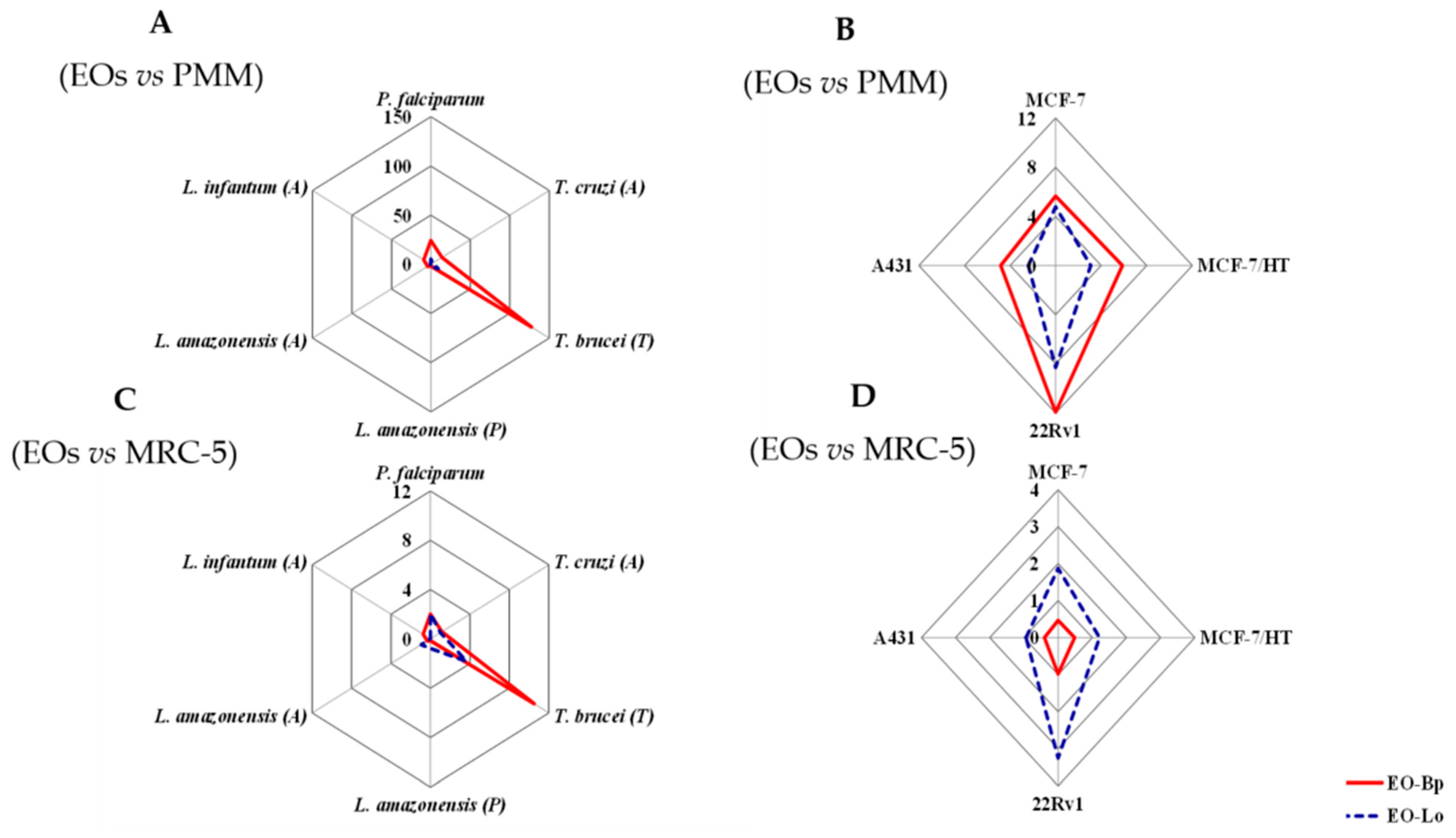
2. Results and Discussion
3. Materials and Methods
3.1. Essential Oils
3.2. Microorganisms and Cells
3.3. Antimicrobial Screening
3.4. Antiproliferative Assay on Malignant Cells
3.5. Cytotoxicity Test on Non-Malignant Cells
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ekiert, H.M.; Szopa, A. Biological activities of natural products. Molecules 2020, 25, 5769. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, A.T.; Ferreira, S.; Duarte, A.P. Genus Ruta: A natural source of high value products with biological and pharmacological properties. J. Ethnopharmacol. 2020, 260, 113076. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.M.; Sun, S.; Kim, M.J.; Phan, N.D.; Tawila, A.M.; Awale, S. Benzophenones from Betula alnoides with antiausterity activities against the PANC-1 human pancreatic cancer cell line. J. Nat. Prod. 2021, 84, 1607–1616. [Google Scholar] [CrossRef]
- Salam, S.; Harneti, D.; Maharani, R.; Nurlelasari; Safari, A.; Hidayat, A.T.; Lesmana, R.; Nafiah, M.A.; Supratman, U.; Prescott, T.A.K.; et al. Cytotoxic triterpenoids from Chisocheton pentandrus. Phytochemistry 2021, 187, 112759. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Edris, A.; Kot, A.M.; Piwowarek, K. Biological activity of some aromatic plants and their metabolites, with an emphasis on health-promoting properties. Molecules 2020, 25, 2478. [Google Scholar] [CrossRef]
- Küçükbay, F.Z.; Kuyumcu, E.; Bilenler, T.; Yıldız, B. Chemical composition and antimicrobial activity of essential oil of Achillea cretica L. (Asteraceae) from Turkey. Nat. Prod. Res. 2012, 26, 1668–1675. [Google Scholar] [CrossRef]
- Pérez Zamora, C.M.; Torres, C.A.; Nuñez, M.B. Antimicrobial activity and chemical composition of essential oils from Verbenaceae species growing in South America. Molecules 2018, 23, 544. [Google Scholar] [CrossRef] [Green Version]
- Perera, W.H.; Bizzo, H.R.; Gama, P.E.; Alviano, C.S.; Salimena, F.R.G.; Alviano, D.S.; Leitão, S.G. Essential oil constituents from high altitude Brazilian species with antimicrobial activity: Baccharis parvidentata Malag., Hyptis monticola Mart. ex Benth. and Lippia origanoides Kunth. J. Essent. Oil Res. 2017, 29, 109–116. [Google Scholar] [CrossRef]
- Ramos Campos, F.; Bressan, J.; Godoy Jasinski, V.C.; Zuccolotto, T.; da Silva, L.E.; Bonancio Cerqueira, L. Baccharis (Asteraceae): Chemical constituents and biological activities. Chem. Biodivers. 2016, 13, 1–17. [Google Scholar] [CrossRef]
- Budel, J.M.; Wang, M.; Raman, V.; Zhao, J.; Khan, S.I.; Rehman, J.U.; Techen, N.; Tekwani, B.; Monteiro, L.M.; Heiden, G.; et al. Essential oils of five Baccharis species: Investigations on the chemical composition and biological activities. Molecules 2018, 23, 2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terblanché, F.C.; Kornelius, G. Essential oil constituents of the genus Lippia (Verbenaceae)—A literature review. J. Essent. Oil Res. 1996, 8, 471–485. [Google Scholar] [CrossRef]
- Pascual, M.E.; Slowing, K.; Carretero, E.; Sánchez Mata, D.; Villar, A. Lippia: Traditional uses, chemistry and pharmacology: A review. J. Ethnopharmacol. 2001, 76, 201–214. [Google Scholar] [CrossRef]
- Ribeiro, A.F.; Andrade, E.H.A.; Salimena, F.R.G.; Maia, J.G.S. Circadian and seasonal study of the cinnamate chemotype from Lippia origanoides Kunth. Biochem. Syst. Ecol. 2014, 55, 249–259. [Google Scholar] [CrossRef]
- Valli, M.; Pivatto, M.; Danuello, A.; Castro-Gamboa, I.; Silva, D.H.S.; Cavalheiro, A.J.; Araujo, A.R.; Furlan, M.; Lopes, M.N.; Bolzani, V.D.S. Tropical biodiversity: Has it been a potential source of secondary metabolites useful for medicinal chemistry? Química Nova 2012, 35, 2278–2287. [Google Scholar] [CrossRef]
- D’agostino, M.; Tesse, N.; Frippiat, J.P.; Machouart, M.; Debourgogne, A. Essential oils and their natural active compounds presenting antifungal properties. Molecules 2019, 24, 3713. [Google Scholar] [CrossRef] [Green Version]
- Luna, E.C.; Silva Luna, I.; Scotti, L.; Monteiro, A.F.M.; Scotti, M.T.; de Moura, R.O.; de Araújo, R.S.A.; Monteiro, K.L.C.; de Aquino, T.M.; Ribeiro, F.F.; et al. Active essential oils and their components in use against neglected diseases and arboviruses. Oxidative Med. Cell. Longev. 2019, 2019, 6587150. [Google Scholar] [CrossRef] [Green Version]
- Blowman, K.; Magalhães, M.; Lemos, M.F.L.; Cabral, C.; Pires, I.M. Anticancer properties of essential oils and other natural products. Evid. Based Complement. Alternat. Med. 2018, 2018, e3149362. [Google Scholar] [CrossRef]
- Spisni, E.; Petrocelli, G.; Imbesi, V.; Spigarelli, R.; Azzinnari, D.; Donati Sarti, M.; Campieri, M.; Valerii, M.C. Antioxidant, anti-inflammatory, and microbial-modulating activities of essential oils: Implications in colonic pathophysiology. Int. J. Mol. Sci. 2020, 21, 4152. [Google Scholar] [CrossRef]
- de Lima, G.P.G.; de Souza, T.M.; de Paula Freire, G.; Farias, D.F.; Cunha, A.P.; Ricardo, N.M.P.S.; de Morais, S.M.; Carvalho, A.F.U. Further insecticidal activities of essential oils from Lippia sidoides and Croton species against Aedes aegypti L. Parasitol. Res. 2013, 112, 1953–1958. [Google Scholar] [CrossRef]
- Masalha, M.; Borovok, I.; Schreiber, R.; Aharonowitz, Y.; Cohen, G. Analysis of transcription of the Staphylococcus aureus aerobic class Ib and anaerobic class III ribonucleotide reductase genes in response to oxygen. J. Bacteriol. 2001, 183, 7260–7272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timbe, P.P.R.; de Souza da Motta, A.; Stincone, P.; Pinilla, C.M.B.; Brandelli, A. Antimicrobial activity of Baccharis dracunculifolia DC and its synergistic interaction with nisin against food-related bacteria. J. Food Sci. Technol. 2021, 58, 3010–3018. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.T.; Maia, B.H.L.D.N.S.; Ferriani, A.P.; Santos, V.A.Q.; Da Cunha, M.A.A.; Teixeira, S.D. Chemical Characterization, Antioxidant Capacity and Antimicrobial Potential of Essential Oil from the Leaves of Baccharis oreophila Malme. Chem. Biodivers. 2019, 16, e1800372. [Google Scholar] [CrossRef]
- Bottieau, E.; Clerinx, J. Human African trypanosomiasis: Progress and stagnation. Infect. Dis. Clin. N. Am. 2019, 33, 61–77. [Google Scholar] [CrossRef]
- Simarro, P.P.; Cecchi, G.; Paone, M.; Franco, J.R.; Diarra, A.; Ruiz, J.A.; Fèvre, E.M.; Courtin, F.; Mattioli, R.C.; Jannin, J.G. The Atlas of Human African trypanosomiasis: A contribution to global mapping of neglected tropical diseases. Int. J. Health Geogr. 2010, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.-J.; Zhang, L.; Ittmann, M.M.; Xin, L. Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proc. Natl. Acad. Sci. USA 2014, 111, E592–E600. [Google Scholar] [CrossRef] [Green Version]
- Juan-Rivera, M.C.; Martínez-Ferrer, M. Integrin inhibitors in prostate cancer. Cancers 2018, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Roubaud, G.; Liaw, B.C.; Oh, W.K.; Mulholland, D.J. Strategies to avoid treatment-induced lineage crisis in advanced prostate cancer. Nat. Rev. Clin. Oncol. 2017, 14, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Dembic, Z. Antitumor drugs and their targets. Molecules 2020, 25, 5776. [Google Scholar] [CrossRef]
- Baldissera, M.D.; de Freitas Souza, C.; Mourão, R.H.V.; da Silva, L.V.F.; Monteiro, S.G. Trypanocidal action of Lippia alba and Lippia origanoides essential oils against Trypanosoma evansi in vitro and in vivo used mice as experimental model. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2017, 41, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.R.; Aires, J.R.D.A.; Higino, T.M.M.; Medeiros, M.D.G.F.D.; Citó, A.M.D.G.L.; Lopes, J.A.D.; de Figueiredo, R.C.B.Q. Trypanocidal and cytotoxic activities of essential oils from medicinal plants of Northeast of Brazil. Exp. Parasitol. 2012, 132, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Escobar, P.; Milena Leal, S.; Herrera, L.V.; Martinez, J.R.; Stashenko, E. Chemical composition and antiprotozoal activities of Colombian Lippia spp. essential oils and their major components. Mem. Inst. Oswaldo Cruz 2010, 105, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.; Fuentes Lorenzo, J.L.; Stashenko, E.E.; Levy, M.; Levy, M.M.; Camarillo, I.G. Lippia origanoides extract induces cell cycle arrest and apoptosis and suppresses NF-ΚB signaling in triple-negative breast cancer cells. Int. J. Oncol. 2017, 51, 1801–1808. [Google Scholar] [CrossRef] [Green Version]
- Heiden, G.; Ribas, O.D.S. Baccharis umbellata (Asteraceae, Astereae): A new species endemic to the highest summits of Paraná, Southern Brazil. Phytotaxa 2012, 49, 23. [Google Scholar] [CrossRef]
- Heiden, G.; Leoni, L.; Nakajima, J. Baccharis magnifica (Asteraceae, Astereae): A striking new species endemic to the summits of Serra Do Caparaó, southeastern Brazil. Phytotaxa 2014, 162, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Sieniawska, E.; Sawicki, R.; Swatko-Ossor, M.; Napiorkowska, A.; Przekora, A.; Ginalska, G.; Augustynowicz-Kopec, E. The effect of combining natural terpenes and antituberculous agents against reference and clinical Mycobacterium tuberculosis strains. Molecules 2018, 23, 176. [Google Scholar] [CrossRef] [Green Version]
- Vimal, A.; Pal, D.; Tripathi, T.; Kumar, A. Eucalyptol, sabinene and cinnamaldehyde: Potent inhibitors of Salmonella yarget protein l-asparaginase. 3 Biotech 2017, 7, 258. [Google Scholar] [CrossRef]
- Mikus, J.; Harkenthal, M.; Steverding, D.; Reichling, J. In vitro effect of essential oils and isolated mono- and sesquiterpenes on Leishmania major and Trypanosoma brucei. Planta Med. 2000, 66, 366–368. [Google Scholar] [CrossRef]
- Chowdhry, L.; Khan, Z.K.; Kulshrestha, D.K. Evaluation of himachalol in murine invasive aspergillosis. Indian J. Exp. Biol. 1996, 39, 449–452. [Google Scholar] [CrossRef]
- Shebaby, W.; Elias, A.; Mroueh, M.; Nehme, B.; El Jalbout, N.D.; Iskandar, R.; Daher, J.C.; Zgheib, M.; Ibrahim, P.; Dwairi, V.; et al. Himachalol induces apoptosis in B16-F10 murine melanoma cells and protects against skin carcinogenesis. J. Ethnopharmacol. 2020, 253, 112545. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.C.R.; Lopes, P.M.; de Azevedo, M.B.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Activities of a-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardo, N.I.; Sainz, P.; González-Coloma, A.; Burillo, J.; Martinez-Díaz, R.A. Trypanocidal effects of essential oils from selected medicinal plants. Synergy among the main components. Nat. Prod. Commun. 2017, 12, 709–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, T.Q.; Felisberto, J.R.S.; Guimarães, E.F.; de Queiroz, G.A.; da Fonseca, A.C.C.; Ramos, Y.J.; Marques, A.M.; Moreira, D.D.L.; Robbs, B.K. Apoptotic effect of β-pinene on oral squamous cell carcinoma as one of the major compounds from essential oil of medicinal plant Piper rivinoides Kunth. Nat. Prod. Res. 2021, 36, 1636–1640. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Vlietinck, A.J.; Vanden, D.; Maesa, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Räz, B.; Iten, M.; Grether-Bühler, Y.; Kaminsky, R.; Brun, R. The alamar blue assay to determine drug sensitivity of African trypanosomes (T.b. rhodesiense and T.b. gambiense) in vitro. Acta Trop. 1997, 68, 139–147. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human Malaria Parasites in Continuous Culture. J. Parasitol. 2005, 91, 484–486. [Google Scholar] [CrossRef]
- Buckner, F.S.; Verlinde, C.L.; La Flamme, A.C.; Van Voorhis, W.C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing β-galactosidase. Antimicrob. Agents Chemother. 1996, 40, 2592–2597. [Google Scholar] [CrossRef] [Green Version]
- Hirumi, H.; Hirumi, K. Continuous cultivation of Trypanosoma brucei blood stream forms in a medium containing a low concentration of serum protein without feeder cell layers. J. Parasitol. 1989, 75, 985–989. [Google Scholar] [CrossRef] [Green Version]
- Sladowski, D.; Steer, S.J.; Clothier, R.H.; Balls, M. An improved MTT assay. J. Immunol. Methods 1993, 157, 203–207. [Google Scholar] [CrossRef]
- Torres-Santos, E.C.; Moreira, D.L.; Kaplan, M.A.; Meirelles, M.N.; Rossi-Bergmann, B. Selective effect of 2′,6′-dihydroxy-4′-methoxychalcone isolated from Piper aduncum on Leishmania amazonensis. Antimicrob. Agents Chemother. 1999, 43, 1234–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komendantova, A.S.; Scherbakov, A.M.; Komkov, A.V.; Chertkova, V.V.; Gudovanniy, A.O.; Chernoburova, E.I.; Sorokin, D.V.; Dzichenka, Y.U.; Shirinian, V.Z.; Volkova, Y.A.; et al. Novel steroidal 1,3,4-thiadiazines: Synthesis and biological evaluation in androgen receptor-positive prostate cancer 22Rv1 Cells. Bioorg. Chem. 2019, 91, 103142. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Infectious Agent | IC50 (μg/mL) | ||
|---|---|---|---|
| EO-Bp | EO-Lo | Rd | |
| E. coli | >64 | >64 | 0.8 (0.76–0.84) |
| S. aureus | 9.2 (8.7–9.7) | >64 | 8.3 (7.9–8.7) |
| C. albicans | >64 | >64 | 2.0 (1.9–2.1) |
| P. falciparum | 3.0 (2.9–3.2) | 14.4 (13.7–15.1) | 0.02 (0.01–0.03) |
| T. cruzi (A) | 5.2 (4.9–5.5) | 29.6 (28.1–31.1) | 0.8 (0.7–0.9) |
| T. brucei (T) | 0.6 (0.5–0.7) | 8.1 (7.7–8.5) | 0.05 (0.02–0.08) |
| L. amazonensis (P) | 39.7 (37.7–41.7) | 37.8 (35.9–39.7) | 0.4 (0.3–0.5) |
| L. amazonensis (A) | 17.4 (16.5–18.3) | 31.8 (30.2–33.4) | 1.3 (1.2–1.4) |
| L. infantum (A) | 8.1 (7.7–8.5) | >64 | 3.7 (3.5–3.9) |
| Malignant Cell Line | IC50 (μg/mL) | ||
|---|---|---|---|
| EO-Bp | EO-Lo | Rd | |
| MCF-7 | 12.9 (12.2–13.5) | 15.8 (15.0–16.6) | 1.9 (1.8–2.0) |
| MCF-7/HT | 12.4 (11.8–13.0) | 24.4 (23.2–25.6) | 3.8 (3.6–4.1) |
| 22Rv1 | 6.1 (5.8–6.4) | 9.1 (8.6–9.5) | 0.81 (0.77–0.85) |
| A431 | 15.1 (14.3–15.8) | 31.5 (30.8–32.1) | 1.2 (1.1–1.3) |
| Cell | CC50 (μg/mL) | |
|---|---|---|
| EO-Bp | EO-Lo | |
| PMM | 72.8 (69.2–72.9) | 75.5 (71.7–79.3) |
| MRC-5 | 6.0 (5.7–6.3) | 29.6 (28.1–31.1) |
| MCF-10A | 15.4 (13.9–16.9) | 29.9 (27.1–32.7) |
| Major Compounds | Pharmacological Property | Target (Results) | Reference |
|---|---|---|---|
Sabinene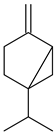 | Antimicrobial |
| [37] |
| [38] | ||
| Antiparasitic |
| [39] | |
| |||
| Antiproliferative | No reports were found | - | |
Himachalol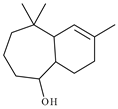 | Antimicrobial |
| [40] |
| Antiparasitic | No reports were found | - | |
| Antiproliferative |
| [41] | |
β-Pinene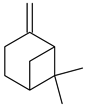 | Antimicrobial |
| [42] |
| Antiparasitic |
| [43] | |
| Antiproliferative |
| [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perera, W.H.; Scherbakov, A.M.; Buravchenko, G.I.; Mikhaevich, E.I.; Leitão, S.G.; Cos, P.; Shchekotikhin, A.E.; Monzote, L.; Setzer, W.N. In Vitro Pharmacological Screening of Essential Oils from Baccharis parvidentata and Lippia origanoides Growing in Brazil. Molecules 2022, 27, 1926. https://doi.org/10.3390/molecules27061926
Perera WH, Scherbakov AM, Buravchenko GI, Mikhaevich EI, Leitão SG, Cos P, Shchekotikhin AE, Monzote L, Setzer WN. In Vitro Pharmacological Screening of Essential Oils from Baccharis parvidentata and Lippia origanoides Growing in Brazil. Molecules. 2022; 27(6):1926. https://doi.org/10.3390/molecules27061926
Chicago/Turabian StylePerera, Wilmer H., Alexander M. Scherbakov, Galina I. Buravchenko, Ekaterina I. Mikhaevich, Suzana Guimarães Leitão, Paul Cos, Andrey E. Shchekotikhin, Lianet Monzote, and William N. Setzer. 2022. "In Vitro Pharmacological Screening of Essential Oils from Baccharis parvidentata and Lippia origanoides Growing in Brazil" Molecules 27, no. 6: 1926. https://doi.org/10.3390/molecules27061926
APA StylePerera, W. H., Scherbakov, A. M., Buravchenko, G. I., Mikhaevich, E. I., Leitão, S. G., Cos, P., Shchekotikhin, A. E., Monzote, L., & Setzer, W. N. (2022). In Vitro Pharmacological Screening of Essential Oils from Baccharis parvidentata and Lippia origanoides Growing in Brazil. Molecules, 27(6), 1926. https://doi.org/10.3390/molecules27061926