
Chitosan Sensitivity of Fungi Isolated from Mango (Mangifera indica L.) with Anthracnose

 , , , and
, , , and 
Abstract
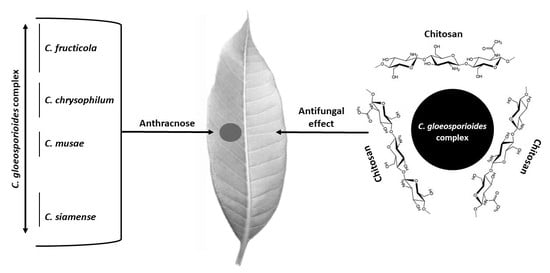
:
1. Introduction
2. Results and Discussion
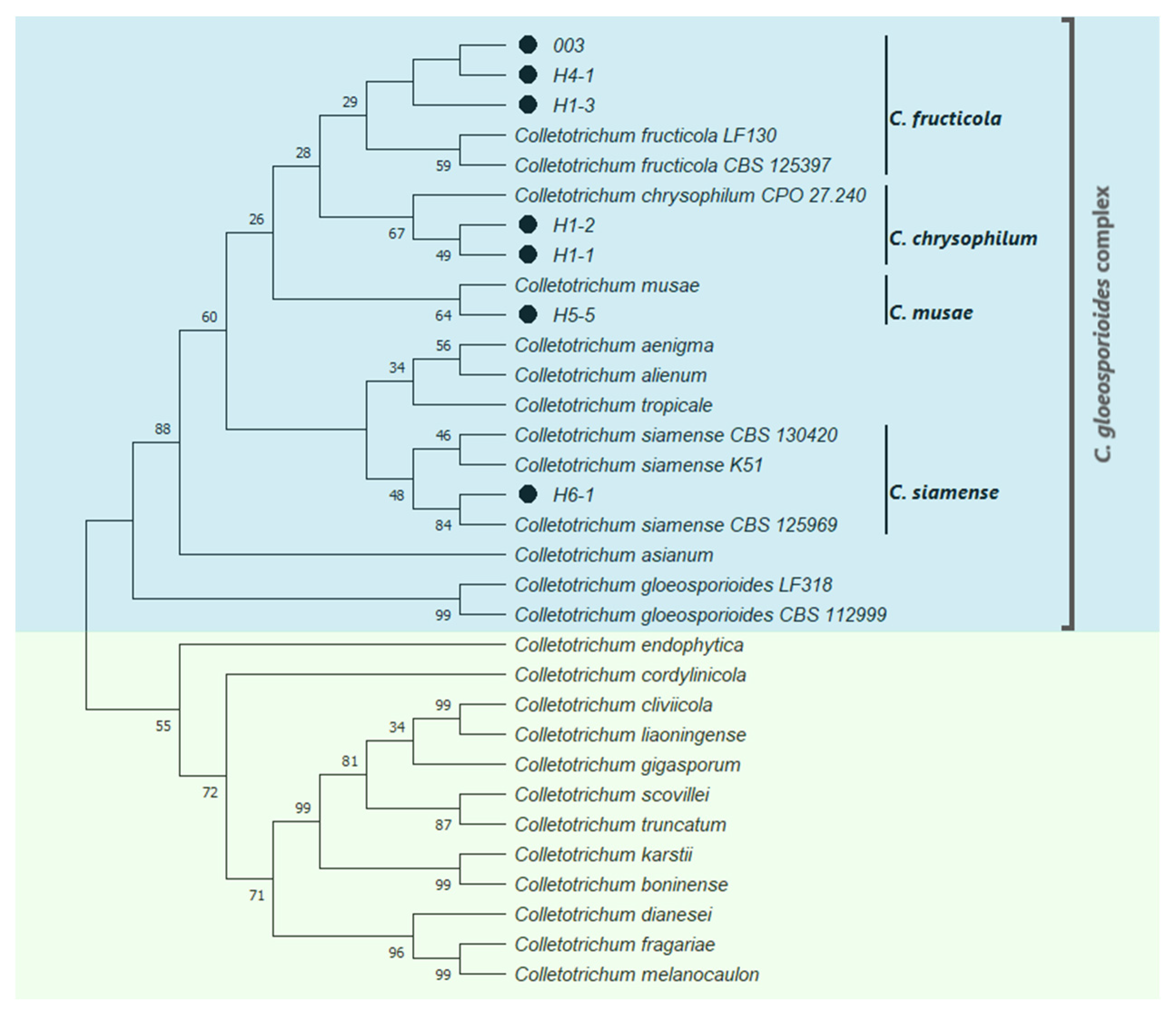
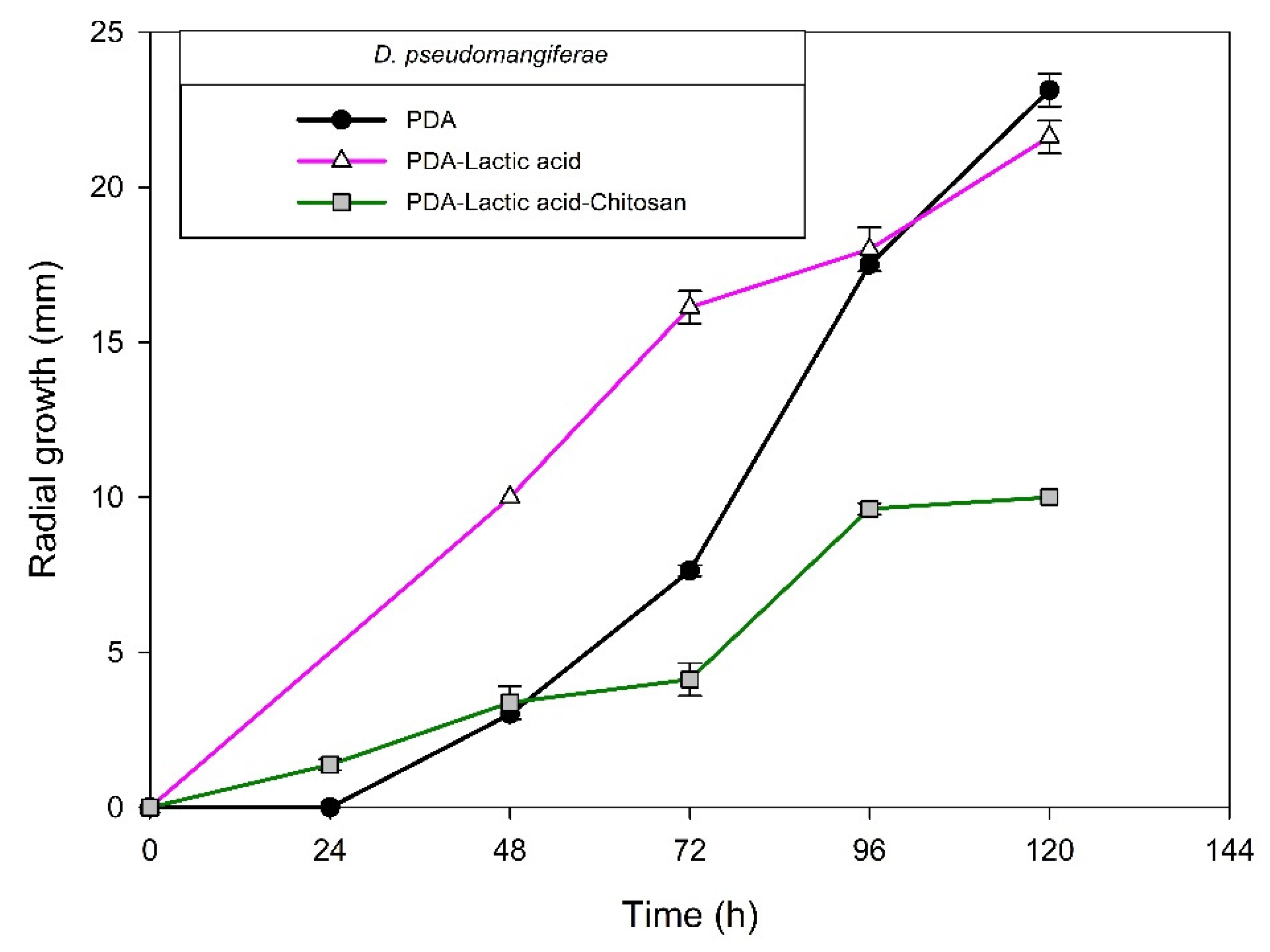
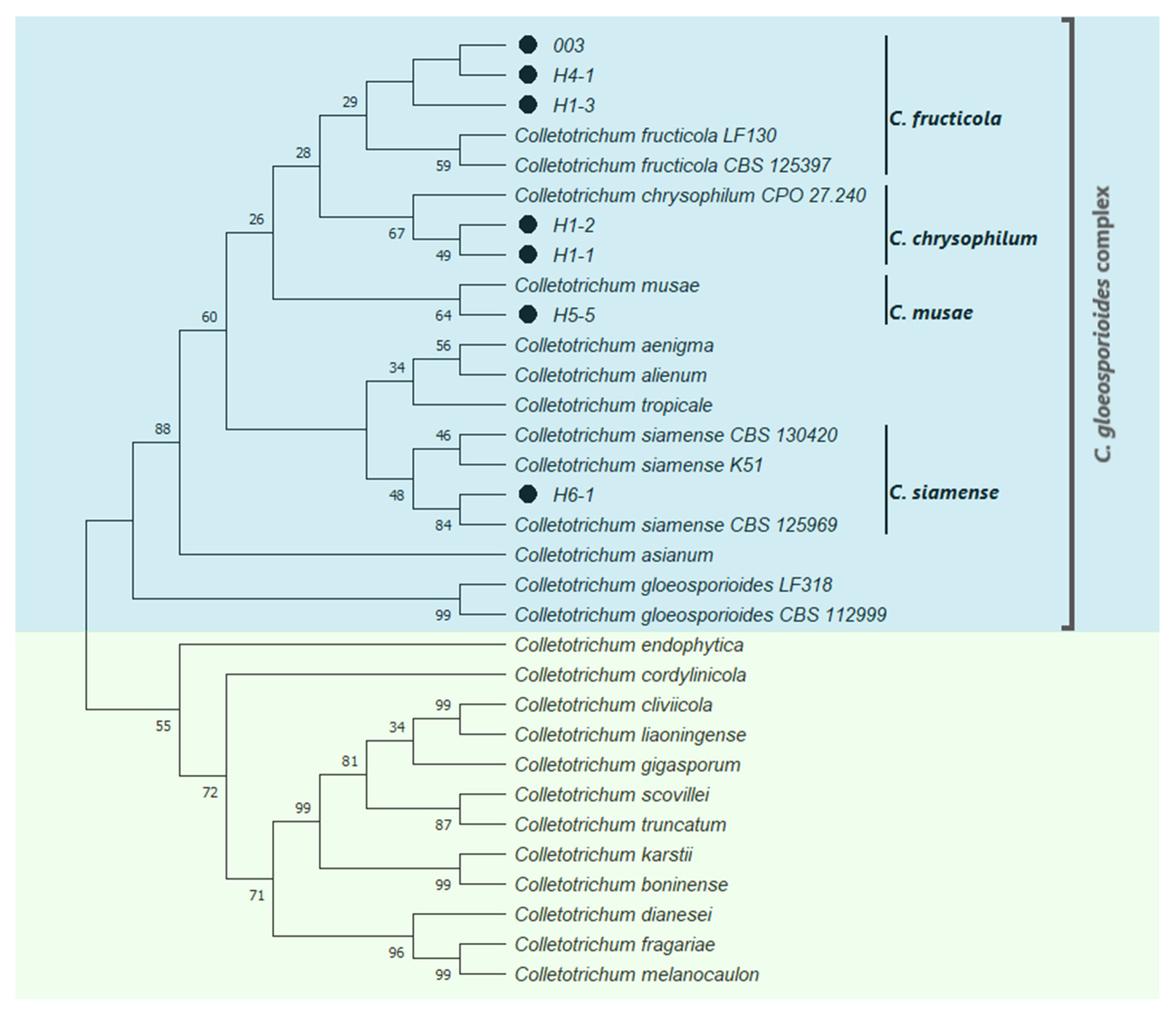
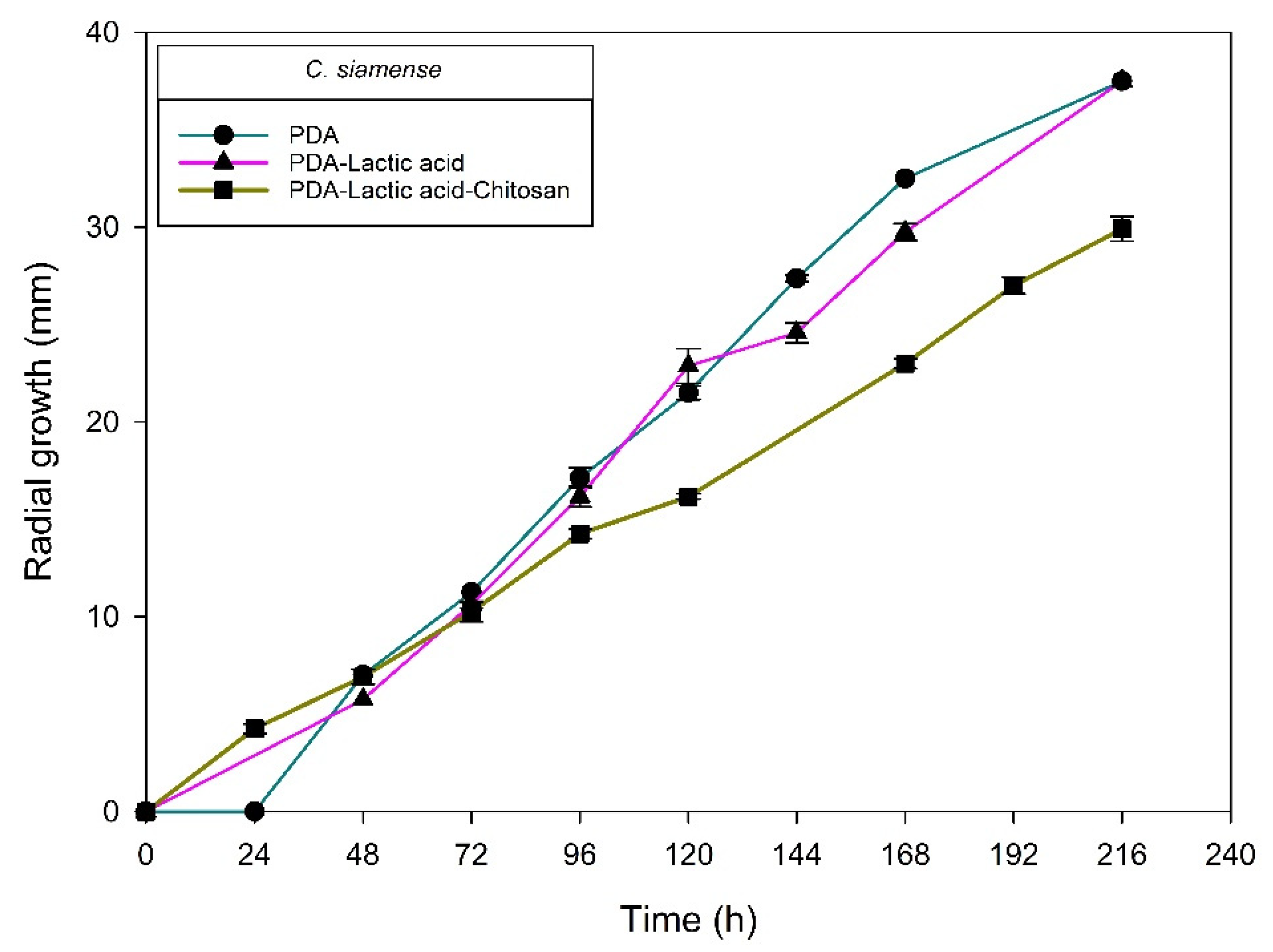
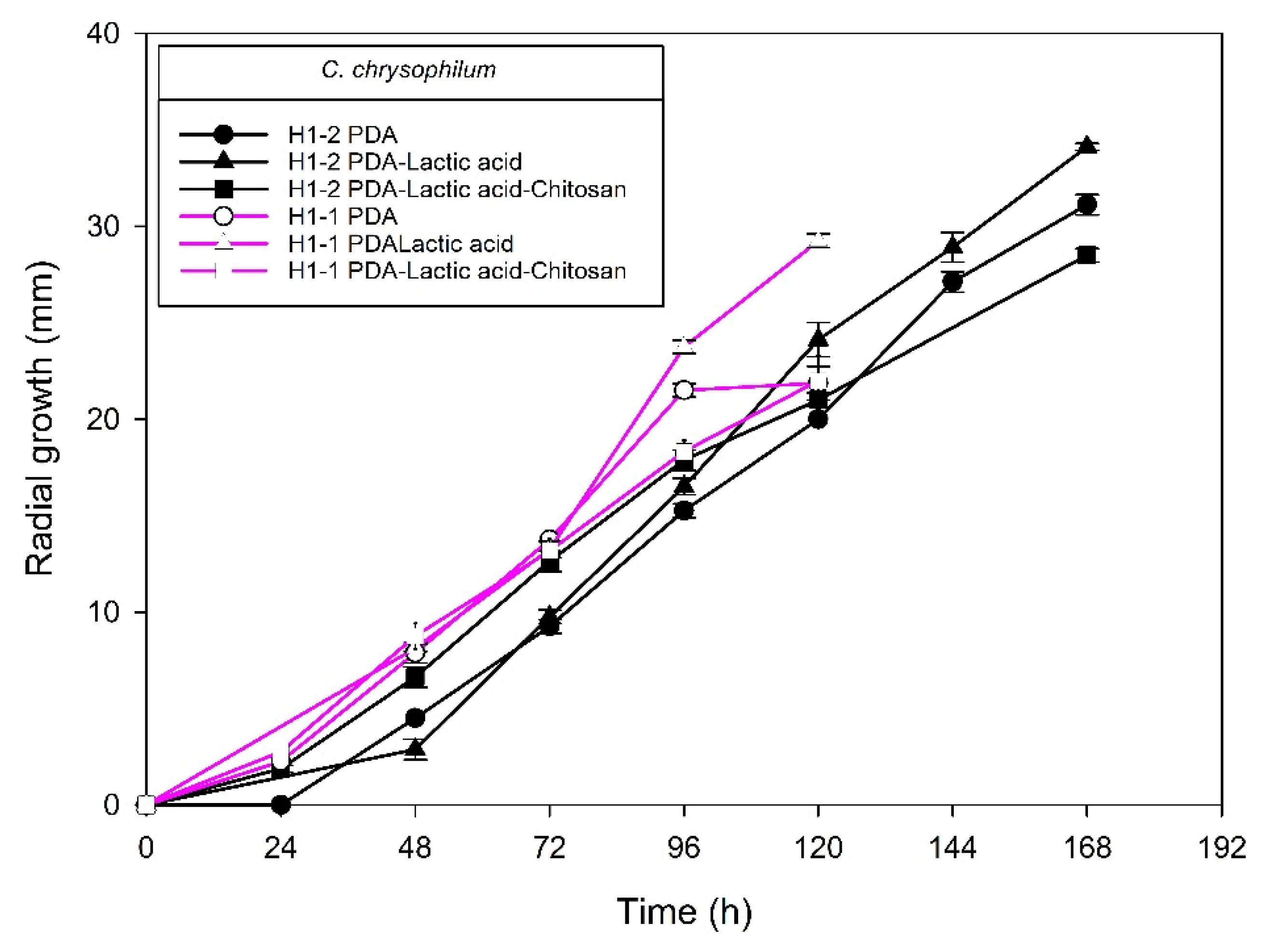
2.1. Identification of Fungal Isolates
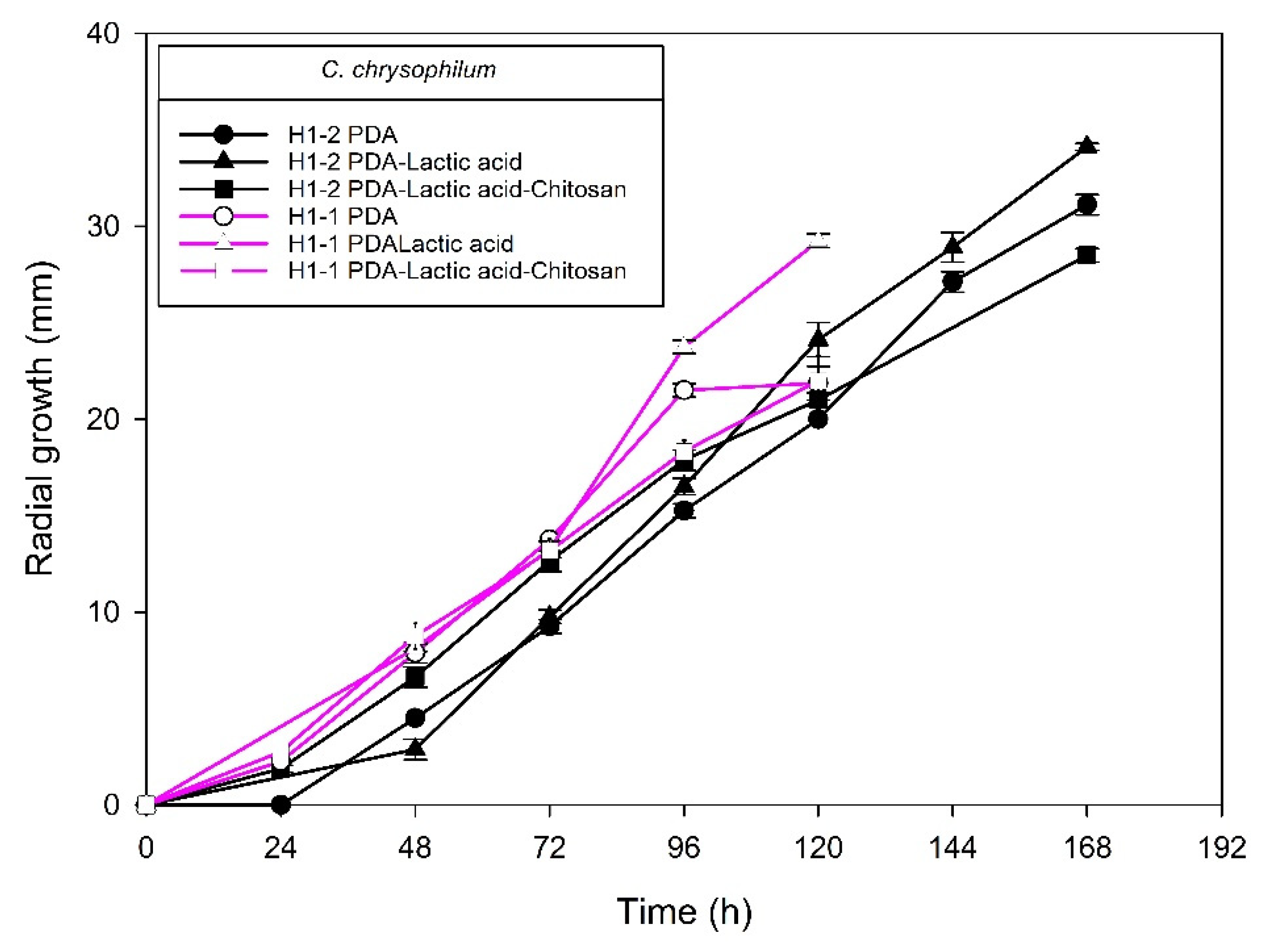
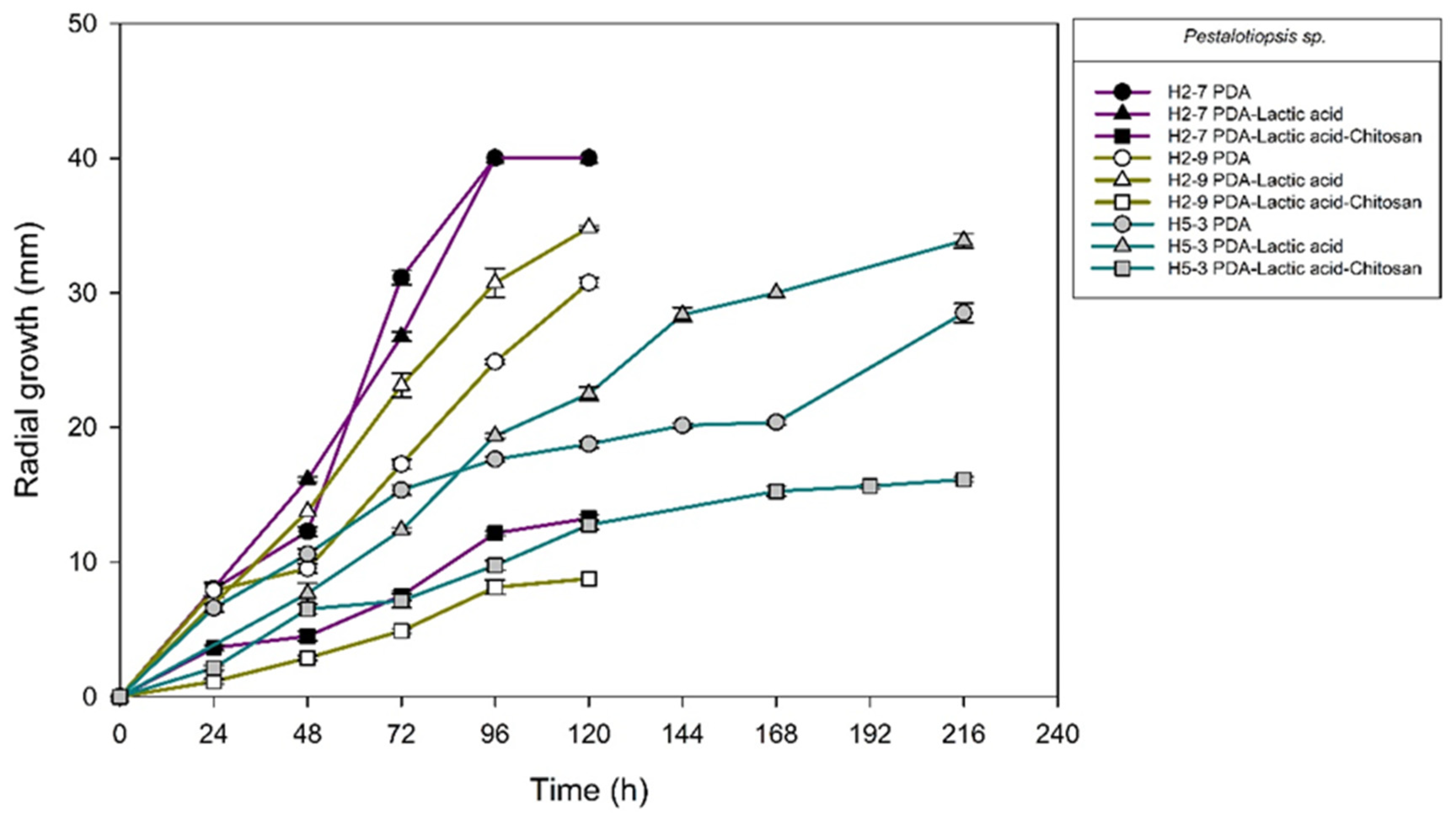
2.2. Chitosan Characterization and Sensitivity of Isolated Fungi
3. Materials and Methods
3.1. Identification of Fungal Isolates
3.2. Chitosan Characterization and Sensitivity of Isolated Fungi
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Herrera-González, J.A.; Bautista-Baños, S.; Salazar-García, S.; Gutiérrez-Martínez, P. Situación actual del manejo poscosecha y de enfermedades fungosas del aguacate ‘Hass’ para exportación en Michoacán. REMEXCA 2000, 11, 1647–1660. [Google Scholar] [CrossRef]
- González-Álvarez, M.; Moreno-Limón, S.; Salcedo-Martínez, S.; Pérez-Rodríguez, E. In vitro Evaluation of Antifungal Activity of Agave (Agave scabra, Salm Dyck) Extracts against Post-Harvest Mushrooms. Phyton (B Aires) 2015, 84, 427–434. [Google Scholar]
- García-Mateos, M.R.; Acosta-Ramos, M.; Rodríguez-Pérez, E.; Vásquez-Sánchez, J.; Hernández-Ramos, L. Extractos vegetales para el control de Colletotrichum gloeosporioides in vitro, en periodo de floración de Carica papaya. Polibotanica 2021, 51, 213–228. [Google Scholar]
- Palou, L.; Montesinos-Herrero, C. Uso de fungicidas en poscosecha de frutos cítricos. Horticultura 2015, 321, 72–76. [Google Scholar]
- Shahid, M.; Zaidi, A.; Rizvi, A.; Saif, S.; Ahmed, B. Recent advances in management strategies of vegetable diseases. In Microbial Strategies for Vegetable Production; Zaidi, A., Khan, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 197–226. [Google Scholar]
- Gan, H.; Wickings, K. Soil ecological responses to pest management in golf turf vary with management intensity, pesticide identity, and application program. Agric. Ecosyst. Environ. 2017, 246, 66–77. [Google Scholar] [CrossRef]
- Mesa, A.M.; Marin, P.A.; Ocampo, O.; Calle, J.; Monsalve, F.Z.I. Fungicidas a partir de extractos vegetales: Una alternativa en el manejo integrado de hongos fitopatógenos. RIA 2019, 45, 23–30. [Google Scholar]
- Landero-Valenzuela, N.; Lara-Viveros, F.M.; Nieto-Angel, D.; Aguado-Rodríguez, J.; Callejas-Hernández, J. Mulberry (Morus alba) leaf extracts as a control alternative of blue mold on postharvest apple fruit. Rev. Mex. Fitopatol. 2017, 35, 1–19. [Google Scholar]
- Maia, T.F.; Donato, A.D.; Fraga, M.E. Review atividade antifúngica de oleos essenciais de plantas. Rev. Bras. Prod. Agroind. 2015, 17, 105–116. [Google Scholar]
- Yari, O.M.; Morales, J.S.; Ramírez-Navas, J.S. Fungicidal effect of chitosan on rust inoculated on coffea. UGCiencia 2016, 22, 45–56. [Google Scholar]
- Rios-Ruiz, C.A.; Bojórquez, V.G.; Cárdenas, R.A.; Mariscal, C.J.; Justo, L.A.C.; Nito, C.M.A.N. Chitosan as a fungicide from shrimp exoskeleton. Rev. Cienc. Tecnol. 2020, 3, 57–62. [Google Scholar]
- Castañeda-Ramírez, J.; Laurel-Ángeles, V.; Espinoza-Zamora, J.; Salcedo-Hernández, R.; López-Ramírez, M.; De la Fuente-Salcido, N. Efecto del quitosano para el biocontrol de hongos fitopatógenos identificados molecularmente de frutas y hortalizas en Guanajuato. Investig. Desarro. Cienc. Tecnol. Aliment. 2016, 1, 207–213. [Google Scholar]
- Hernández-Téllez, C.; Cortez-Rocha, M.; Burgos-Hernández, A.; Rosas-Burgos, E.; Lizardi-Mendoza, J.; Torres-Arreola, W.; Burboa-Zazueta, M.; Plascencia-Jatomea, M. Chitosan/carrageenan/lysozyme particles: Synthesis, characterization and antifungal activity against Aspergillus parasiticus. Rev. Mex. Ing. Quim. 2018, 17, 897–912. [Google Scholar] [CrossRef]
- Rodríguez-Romero, V.M.; Villanueva-Arce, R.; Trejo-Raya, A.B.; Silvía, B.-B. Chitosan and Pseudomonas fluorescens extracts for Alternaria alternata control in tomato (Solanum lycopersicum). Mex. J. Phytopathol. 2019, 37, 202–219. [Google Scholar]
- Berumen, G.V.; Coronado, L.D.P.; Ochoa, V.A.J.; Chacón, M.A.L.; Gutiérrez, P.M. Effect of chitosan on the induction of disease resistance against Colletotrichum sp. in mango (Mangifera indica L.) cv. Tommy Atkins. Investig. Cienc. 2015, 23, 16–21. [Google Scholar]
- Bautista-Baños, S.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.; Corona-Rangel, M.L. Chitosan: A versatile antimicrobial polysaccharide for fruit and vegetables in postharvest–a review. Rev. Chapingo Ser. Hortic. 2017, 23, 103–121. [Google Scholar] [CrossRef]
- SIAP (Servicio de Información Agroalimentaria y Pesquera). Panorama Agroalimentario 2020; Secretaria de Agricultura y Desarrrollo Social: Gobierno de México, México, 2020. Available online: https://nube.siap.gob.mx/gobmx_publicaciones_siap/ (accessed on 11 August 2021).
- Mendoza-Herrera, Y.M.; Rios-Velasco, C.; Cambero-Campos, J.; De dios-Ávila, N.; Pérez-Corral, D.A.; Rodríguez-Guerra, R.; Estrada-Virgen, M.O. Antagonistic actinomycetes to Colletotrichum sp. Penz in the mango cultivation in Nayarit, Mexico. Ecosistemas Y Recur. Agropecu. 2020, 7, 1–9. [Google Scholar]
- Valdés, L.A.; Calero Consuegra, D.; Carballo, M.E.; Capote, M.; González, I.; Álvarez, J.M. Characterization morphological, cultural and pathogenic of isolated Colletotrichum sp. anthracnose producing in mango (Mangifera indica L.). La Granja Rev. De Cienc. De La Vida 2017, 26, 38–51. [Google Scholar] [CrossRef] [Green Version]
- Lima, O.P.D.; de Oliveira, K.Á.R.; Dos Santos Vieira, W.A.; Câmara, M.P.S.; de Souza, E.L. Control of anthracnose caused by Colletotrichum species in guava, mango and papaya using synergistic combinations of chitosan and Cymbopogon citratus (D.C. ex Nees) Stapf. essential oil. Int. J. Food Microbiol. 2018, 266, 87–94. [Google Scholar] [CrossRef]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on fruit crops: A “complex” challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef]
- Tovar-Pedraza, J.M.; Mora-Aguilera, J.A.; Nava-Díaz, C.; Lima, N.B.; Michereff, S.J.; Sandoval-Islas, J.S.; Camara, M.P.S.; Téliz-Ortiz, D.; Leyva-Mir, S.G. Distribution and pathogenicity of Colletotrichum species associated with mango anthracnose in Mexico. Plant Dis. 2020, 104, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Bu, J.; Shu, J.; Yu, Z.; Tang, L.; Huang, L.; Guo, T.; Mo, J.; Luo, S.; Solangi, G.S.; et al. Colletotrichum species associated with mango in southern China. Sci. Rep. 2019, 9, 18891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaria, L. Diversity of Colletotrichum species associated with anthracnose disease in tropical fruit crops. A review. Agriculture 2021, 11, 297. [Google Scholar] [CrossRef]
- Arauz, L.F. Mango anthracnose: Economic impact and current options for integrated management. Plant Dis. 2000, 84, 600–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, N.H.; Mohd, M.; Nor, N.M.I.M.; Zakaria, L. Characterization and pathogenicity of Fusarium species associated with leaf spot of mango (Mangifera indica L.). Microb. Pathog. 2018, 114, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Rakesh, A.; Mishra, J.P.; Prasad, R.; Sekhar, J.C.; Gupta, D.; Reddy, V.P.; Kumar, S. Isolation and in vitro evaluation of fungicides against Pestalotiopsis mangiferae causing grey blight of mango. J. Pharmacogn. Phytochem. 2020, 9, 1368–1370. [Google Scholar]
- Niren, M.; Chandra, M.N. Evaluation of Media for growth and sporulation characteristics of postharvest pathogens Curvularia lunata and Pestalotiopsis mangiferae. Ann. Plant Sci. 2019, 27, 89–94. [Google Scholar]
- Lim, L.; Mohd, M.H.; Zakaria, L. Identification and pathogenicity of Diaporthe species associated with stem-end rot of mango (Mangifera indica L.). Eur. J. Plant Pathol. 2019, 155, 687–696. [Google Scholar] [CrossRef]
- Garg, N.; Singh, B.; Vaish, S.; Kumar, S.; Arora, S. Exploring microbial community diversity of mango leaf compost. Curr. Hortic. 2021, 9, 27–35. [Google Scholar] [CrossRef]
- Verlee, A.; Mincke, S.; Stevens, C.V. Recent developments in antibacterial and antifungal chitosan and its derivatives. Carbohydr. Polym. 2017, 164, 268–283. [Google Scholar] [CrossRef]
- Grande-Tovar, C.D.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan coatings enriched with essential oils: Effects on fungi involved in fruit decay and mechanisms of action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, L.; Zeng, K. Efficacy of Pichia membranaefaciens combined with chitosan against Colletotrichum gloeosporioides in citrus fruits and possible modes of action. Biol. Control. 2016, 96, 39–47. [Google Scholar] [CrossRef]
- Algam, S.A.; Elwagia, M.E. Evaluation of chitosan efficacy on tomato growth and control of early blight disease. Jordan J. Agric. Sci. 2015, 11, 27–36. [Google Scholar]
- Dibona-Villanueva, L.; Fuentealba, D. Novel Chitosan-Riboflavin Conjugate with Visible Light-Enhanced Antifungal Properties against Penicillium digitatum. J. Agric. Food Chem. 2021, 69, 945–954. [Google Scholar] [CrossRef]
- Peña, A.; Sánchez, N.S.; Calahorra, M. Effects of Chitosan on Candida albicans: Conditions for Its Antifungal Activity. BioMed Res. Int. 2013, 527549, 15. [Google Scholar]
- Ai, H.; Wang, F.; Xia, Y.; Chen, X.; Lei, C. Antioxidant, antifungal and antiviral activities of chitosan from the larvae of housefly, Musca domestica L. Food Chem. 2012, 132, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, X.; Liu, J.; Zhang, W.; Tang, X. Molecular weight-dependent antifungal activity and action mode of chitosan against Fulvia fulva (cooke). J. Appl. Polym. Sci. 2010, 119, 3127–3135. [Google Scholar] [CrossRef]
- Jiangtao, W.; Hedong, W. Preparation of soluble p-aminobenzoyl chitosan ester by Schiff’s base and antibacterial activity of the derivatives. Int. J. Biol. Macromol. 2011, 48, 523–529. [Google Scholar]
- Hosseinnejad, M.; Jafari, S.M. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Jinasena, D.; Pathirathna, P.; Wickramarachchi, S.; Marasinghe, E. Use of chitosan to control anthracnose on “Embul” banana. In Proceedings of the International Conference on Asia Agriculture and Animal, IPCBEE, Hong Kong, China, 2–3 July 2011; IACSIT Press: Singapore, 2011; Volume 13, pp. 56–60. [Google Scholar]
- Nascimento, J.I.G.; Stamford, T.C.M.; Melo, N.F.C.B.; Nunes, I.d.S.; Lima, M.A.B.; Pintado, M.M.E.; Stamford, T.L.M. Chitosan-citric acid edible coating to control Colletotrichum gloeosporioides and maintain quality parameters of fresh-cut guava. Int. J. Biol. Macromol. 2020, 163, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.L.R.; Stamford, T.C.M.; de Oliveira, K.Á.R.; Pessoa, A.D.M.P.; de Lima, M.A.B.; Pintado, M.M.E.; Câmara, M.P.S.; de Oliveira Franco, L.; Magnani, M.; de Souza, E.L. Chitosan produced from Mucorales fungi using agroindustrial by-products and its efficacy to inhibit Colletotrichum species. Int. J. Biol. Macromol. 2018, 108, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Obregón, E.A.; Plascencia-Jatomea, M.; López-Cervantes, J.; Cira-Chávez, L.A.; Sánchez-Machado, D.I.; Cortez-Rocha, M.O. Antifungal activity of chitosan in Cladosporium cladosporioides isolated from safflower. Rev. Mex. Mic. 2011, 34, 93–96. [Google Scholar]
- Walker, G.M.; White, N.A. Introduction to Fungal Physiology. In Fungi: Biology and Applications, 3rd ed.; Kavanagh, K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–35. [Google Scholar]
- Hu, M.-J.; Grabke, A.; Schnabel, G. Investigation of the Colletotrichum gloeosporioides species complex causing peach anthracnose in South Carolina. Plant Dis. 2015, 99, 797–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.K. Anthracnose diseases of some common medicinally important fruit plants. J. Med. Plants 2016, 4, 233–236. [Google Scholar]
- Jiang, J.; Zhai, H.; Li, H.; Wang, Z.; Chen, Y.; Hong, N.; Wang, G. Chofong, G.N.; Xu, W. Identification and characterization of Colletotrichum fructicola causing black spots on young fruits related to bitter rot of pear (Pyrus bretschneideri Rehd.) in China. Crop Prot. 2014, 58, 41–48. [Google Scholar] [CrossRef]
- Villafana, R.T.; Rampersad, S.N. Diversity, structure, and synteny of cutinase gene of Colletotrichum species. Ecol. Evol. 2020, 10, 1425–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes-Aragón, D.; Guarnaccia, V.; Rebollar-Alviter, A.; Juárez-Vázquez, S.B.; Aguirre-Rayo, F.; Silva-Rojas, H.V. Multilocus identification and thiophanate-methyl sensitivity of Colletotrichum gloeosporioides species complex associated with fruit with symptoms and symptomless leaves of mango. Plant Pathol. 2020, 69, 1125–1138. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000; pp. 147–163. [Google Scholar]
- Khan, T.A.; Peh, K.K.; Ch’ng, H.S. Reporting degree of deacetylation values of chitosan: The influence of analytical methods. J. Pharm. Pharm. Sc. 2002, 5, 205–212. [Google Scholar]
- Brugnerotto, J.; Lizardi, J.; Goycoolea, F.M.; Argüelles-Monal, W.; Desbrieres, J.; Rinaudo, M. An infrared investigation in relation with chitin and chitosan characterization. Polymer 2001, 42, 3569–3580. [Google Scholar] [CrossRef]
- Seo, S.; King, J.M.; Prinyawiwatkul, W. Simultaneous depolymerization and decolorization of chitosan by ozone treatment. Food Sci. 2007, 72, C552–C556. [Google Scholar] [CrossRef] [PubMed]
- Badawy, M.E.; Rabea, E.I. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. Int. J. Carbohydr. Chem. 2011, 460381, 1–30. [Google Scholar] [CrossRef]
- Solomon, O.F.; Ciutǎ, I.Z. Détermination de la viscosité intrinsèque de solutions de polymères par une simple détermination de la viscosité. J. Appl. Polym. Sci. 1962, 6, 683–686. [Google Scholar] [CrossRef]
- Seuvre, A.M.; Mathlouthi, M. Solutions properties and solute–solvent interactions in ternary sugar–salt–water solutions. Food Chem. 2010, 122, 455–461. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Meletiadis, J.; Meis, J.F.G.M.; Mouton, J.W.; Verweij, P.E. Analysis of Growth Characteristics of Filamentous Fungi in Different Nutrient Media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
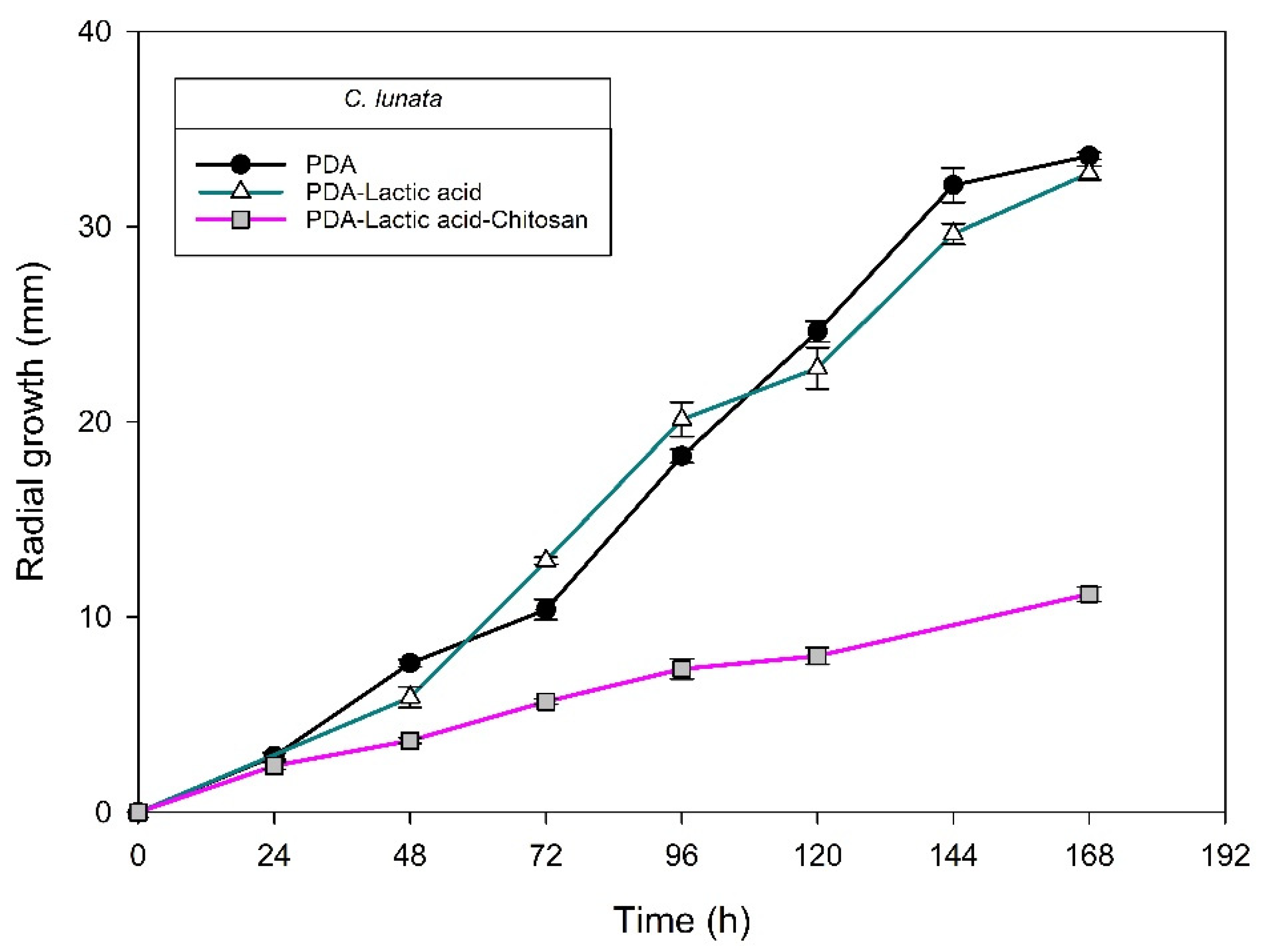
| Isolate/Species | Rate of Growth in Log Phase μm h−1 | Inhibition of Radial Growth 1 (%) by Chitosan at 120 h | ||
|---|---|---|---|---|
| PDA | PDA-Lactic Acid | PDA-Lactic Acid-Chitosan | ||
| H4-1/C. fructicola | 212 ± 3 CDa2 | 208 ± 3 Da | 202 ± 4 ABa | 2.84 ± 2.30 D |
| H1-3/C. fructicola | 248 ± 2 Bb | 268 ± 3 Ba | 217 ± 2 Ac | 13.38 ± 1.28 C |
| 003/C. fructicola | 30 ± 4 Fb2 | 117 ± 5 Fa | 126 ± 0 Da | 0.00 ± 0.00 D |
| H6-1/C. siamense | 175 ± 4 Ea | 173 ± 3 Ea | 135 ± 4 CDb | 29.95 ± 0.62 A |
| H1-2/C. chrysophilum | 228 ± 0 Cb | 242 ± 0 Ca | 149 ± 2 Cc | 12.95 ± 1.46 C |
| H1-1/C. chrysophilum | 207 ± 6 Db | 307 ± 1 Aa | 187 ± 7 Bc | 22.43 ± 1.20 B |
| H5-5/C. musae | 341 ± 5 Aa | 236 ± 5 Cb | 198 ± 4 Bc | 18.44 ± 1.37 B |
| Isolate/Species | Radial Growth Inhibition1 (%) by Chitosan (g L−1) at 120 h | |||
|---|---|---|---|---|
| 0.1 | 0.5 | 0.75 | 1 | |
| H4-1/C. fructicola | 13.85 ± 0.93 2 | 12.27 ± 1.08 | 0.00 ± 0.00 | 2.84 ± 2.30 |
| H1-3/C. fructicola | 0.00 ± 0.00 | 10.09 ± 0.70 | 12.84 ± 1.29 | 13.38 ± 1.28 |
| 003/C. fructicola | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| H6-1/C. siamense | 6.00 ± 0.54 | 12.56 ± 0.28 | 23.44 ± 2.95 | 29.95 ± 0.62 |
| H1-2/C. chrysophilum | 40.89 ± 2.16 | 26.40 ± 1.23 | 42.99 ± 0.62 | 12.95 ± 1.46 |
| H1-1/C. chrysophilum | 0.00 ± 0.00 | 20.08 ± 0.36 | 19.22 ± 0.371 | 22.43 ± 1.20 |
| H5-5/C.musae | 0.00 ± 0.00 | 13.16 ± 0.41 | 11.69 ± 1.13 | 18.44 ± 1.37 |
| 004/Alternaria sp. | 0.00 ± 0.00 | 18.67 ± 2.84 | 19.31 ± 0.97 | 20.28 ± 2.40 |
| 009-1/Alternaria sp. | 0.00 ± 0.00 | 37.08 ± 1.94 | 33.37 ± 1.74 | 24.07 ± 2.62 |
| H6-1b/Alternaria tenuissima | 0.00 ± 0.00 | 68.83 ± 0.00 | 68.18 ± 2.75 | 72.72 ± 1.84 |
| H2-2/Fusarium sp. | 0.00 ± 0.00 | 53.46 ± 0.54 | 39.61 ± 0.54 | 71.54 ± 0.00 |
| H2-3/Fusarium sp. | 38.39 ± 0.92 | 77.25 ± 0.15 | 81.51 ± 0.54 | 87.62 ± 0.00 |
| H2-7/Pestalotiopsis sp. | 0.00 ± 0.00 | 54.68 ± 0.44 | 67.81 ± 1.32 | 67.18 ± 0.44 |
| H2-9/Pestalotiopsis sp. | 0.00 ± 0.00 | 62.23 ± 2.54 | 59.71 ± 2.03 | 74.87 ± 0.00 |
| H5-3/Pestalotiopsis sp. | 16.28 ± 0.53 | 49.63 ± 0.62 | 54.49 ± 0.07 | 43.33 ± 1.57 |
| H3-8/Curvularia lunata | 0.00 ± 0.00 | 50.53 ± 0.75 | 59.33 ± 0.34 | 65.22 ± 1.88 |
| 008/Diaporthe pseudomangiferae | 0.00 ± 0.00 | 45.66 ± 0.30 | 60.72 ± 2.30 | 53.75 ± 0.0 |
| H6-2/Epicoccum nigrum | 10.26 ± 0.37 | 44.42 ± 0.80 | 59.82 ± 0.24 | 69.22 ± 1.71 |
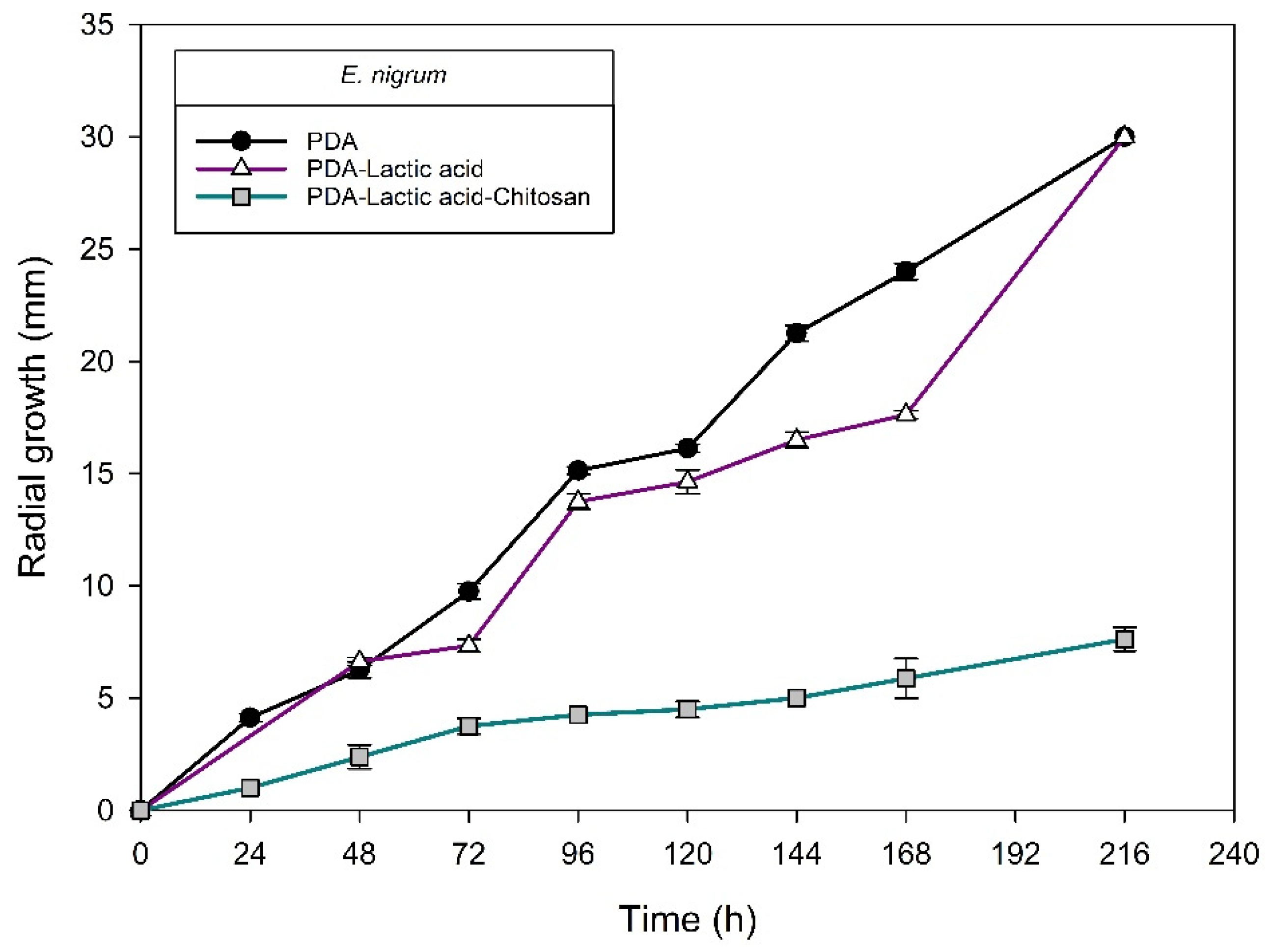
| Isolate/Specie | Radial Growth Rate: Log Phase (μm h−1) | Inhibition1 of Radial Growth (%) by Chitosan at 120 h | ||
|---|---|---|---|---|
| PDA | PDA-Lactic Acid | PDA-Lactic Acid Chitosan | ||
| 004/Alternaria sp. | 116 ± 2 a2 | 60 ± 1 b | 35 ± 1 c | 20.28 ± 2.40 G |
| 009-1/Alternaria sp. | 192 ± 3 b | 294 ± 3 a | 17 ± 4 c | 24.07 ± 2.62 G |
| H6-1b/Alternaria tenuissima | 76 ± 3 b | 155 ± 3 a | 24 ± 3 c | 72.72 ± 1.84 BC |
| H2-2/Fusarium sp. | 206 ± 1 b | 255 ± 2 a | 105 ± 7c | 71.54 ± 0.00 BC |
| H2-3/Fusarium sp. | 277 ± 1 a | 228 ± 3 b | 23 ± 4 c | 87.62 ± 0.00 A |
| H2-7/Pestalotiopsis sp. | 578 ± 7 a | 497± 4 b | 159 ± 4 c | 67.18 ± 0.44 CD |
| H2-9/Pestalotiopsis sp. | 297 ± 1 a | 294 ±1 b | 87 ± 1 c | 74.87 ± 0.00 B |
| H5-3/Pestalotiopsis sp. | 40 ± 0 c | 158 ± 1 a | 59 ± 1 b | 43.33 ± 1.57 F |
| H3-8/Curvularia lunata | 223 ± 1 a | 186 ± 8 b | 55 ± 1 c | 65.22 ± 1.88 D |
| 008/Diaporthe pseudomangiferae | 293 ± 6 a | 153± 7 b | 106 ± 8 c | 53.75 ± 0.0 E |
| H6-2/Epicoccum nigrum | 133 ± 2 a | 56 ± 3 b | 12 ± 1 c | 69.22 ± 1.71 BCD |
| Genetic Marker | Forward (5′-3′) | Reverse (5′-3′) | Reference |
|---|---|---|---|
| Internal transcribed spacers (ITS) | ITS1 (TCCGTAGGTGAACCTGCGG) | ITS4 (TCCTCCGCTTATTGATATGC) | [48,49] |
| β-tubulin 2 (β-Tub2) | Bt2a (GGTAACCAAATCGGTGCTGCTTTC) | Bt2b (ACCCTCAGTGTAGTGACCCTTGGC) | [48] |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | GDF1 (GCCGTCAACGACCCCTTCATTGA) | GDR1 (GGGTGGAGTCGTACTTGAGCATGT) | [48] |
| Actin (Act) | ACT-512F (ATGTGCAAGGCCGGTTTCGC) | ACT-783R (TACGAGTCCTTCTGGCCCAT) | [48,49] |
| Calmodulin (CaM) | CL1C (GAATTCAAGGAGGCCTTCTC) | CL2C (CTTCTGCATCATGAGCT GAC) | [48] |
| Chitin synthase (CHS-1) | CHS1-79F (TGGGGCAAGGATGCTTGGAAGAAG) | CHS-1-354R (TGGAAGAACCATCTGTGAGAGTTG) | [48,49] |
| Apn2-Mat1-2 intergenic spacer (ApMat) | AMF1 (TCATTCTACGTATGTGCCCG) | AMR1 (CCAGAAATACACCGAACTTGC) | [50] |
| Number or Name of Equation | Equation |
|---|---|
| 1 (DA: acetylation degree) | |
| 2 (DD: deacetylation degree) | |
| Relative viscosity | ηrel = tflux solution of chitosan/tflux of solvent |
| Specific viscosity | ηsp = ηrel – 1 |
| Huggins | [η] = ηsp/C |
| Kramer | [η] = (ln ηrel)/C |
| Mark–Houwink–Sakurada | [ƞ] = k (Mv)α |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela-Ortiz, G.; Gaxiola-Camacho, S.M.; San-Martín-Hernández, C.; Martínez-Téllez, M.Á.; Aispuro-Hernández, E.; Lizardi-Mendoza, J.; Quintana-Obregón, E.A. Chitosan Sensitivity of Fungi Isolated from Mango (Mangifera indica L.) with Anthracnose. Molecules 2022, 27, 1244. https://doi.org/10.3390/molecules27041244
Valenzuela-Ortiz G, Gaxiola-Camacho SM, San-Martín-Hernández C, Martínez-Téllez MÁ, Aispuro-Hernández E, Lizardi-Mendoza J, Quintana-Obregón EA. Chitosan Sensitivity of Fungi Isolated from Mango (Mangifera indica L.) with Anthracnose. Molecules. 2022; 27(4):1244. https://doi.org/10.3390/molecules27041244
Chicago/Turabian StyleValenzuela-Ortiz, Griselda, Soila Maribel Gaxiola-Camacho, Cesar San-Martín-Hernández, Miguel Ángel Martínez-Téllez, Emmanuel Aispuro-Hernández, Jaime Lizardi-Mendoza, and Eber Addí Quintana-Obregón. 2022. "Chitosan Sensitivity of Fungi Isolated from Mango (Mangifera indica L.) with Anthracnose" Molecules 27, no. 4: 1244. https://doi.org/10.3390/molecules27041244
APA StyleValenzuela-Ortiz, G., Gaxiola-Camacho, S. M., San-Martín-Hernández, C., Martínez-Téllez, M. Á., Aispuro-Hernández, E., Lizardi-Mendoza, J., & Quintana-Obregón, E. A. (2022). Chitosan Sensitivity of Fungi Isolated from Mango (Mangifera indica L.) with Anthracnose. Molecules, 27(4), 1244. https://doi.org/10.3390/molecules27041244