
Induction of Apoptosis via Inactivating PI3K/AKT Pathway in Colorectal Cancer Cells Using Aged Chinese Hakka Stir-Fried Green Tea Extract

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Freeze-Dried Powder of Tea Extract
2.2. Analysis of Active Ingredients in Tea Extract
2.3. Cell Viability Assay
2.4. Cell Cycle Assay
2.5. Cell Apoptosis Assay
2.6. Western Blotting Assay
2.7. Data Statistics and Analysis
3. Results
3.1. Identification of Active Ingredients in the Extracts of Chinese Green Tea (HSGT) Aged for Different Years
3.2. Aged Chinese Green Tea (HSGT) Inhibits the Proliferation of Colon Cancer Cells
3.3. Aged Chinese Green Tea (HSGT) Promotes Cell Apoptosis
3.4. Aged Chinese Green Tea (HSGT) Slows Down Cell-Cycle Progression
3.5. Aged Chinese Green Tea (HSGT) Reduces the Expression of Cell-Cycle-Related Proteins in Colon Cancer Cells
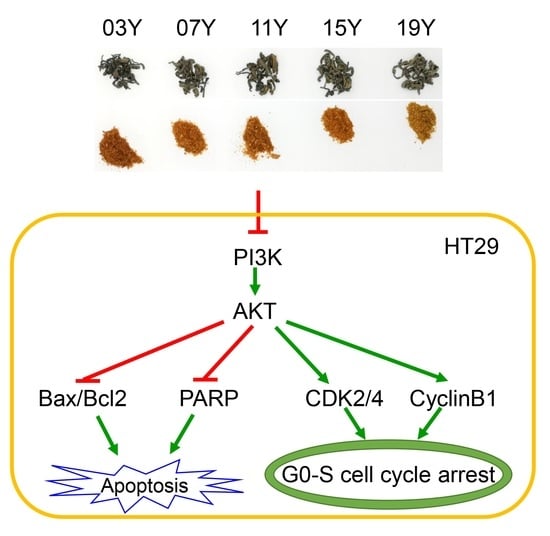
3.6. Aged Chinese Green Tea (HSGT) Inactivates PI3K/AKT Signalling and Enhances Apoptotic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kyu, H.H.; Bachman, V.F.; Alexander, L.T.; Mumford, J.E.; Afshin, A.; Estep, K.; Veerman, J.L.; Delwiche, K.; Iannarone, M.L.; Moyer, M.L.; et al. Physical activity and risk of breast cancer, colon cancer, diabetes, ischemic heart disease, and ischemic stroke events: Systematic review and dose-response meta-analysis for the Global Burden of Disease Study 2013. BMJ 2016, 354, i3857. [Google Scholar] [CrossRef] [PubMed]
- Zaborowski, A.M.; Abdile, A.; Adamina, M.; Aigner, F.; d’Allens, L.; Allmer, C.; Álvarez, A.; Anula, R.; Andric, M.; Atallah, S.; et al. Characteristics of early-onset vs late-onset colorectal cancer: A review. JAMA Surg. 2021, 156, 865–874. [Google Scholar] [CrossRef]
- Biller, L.H.; Schrag, D. Diagnosis and treatment of metastatic colorectal cancer: A review. JAMA 2021, 325, 669–685. [Google Scholar] [CrossRef]
- Bars-Cortina, D.; Sakhawat, A.; Piñol-Felis, C.; Motilva, M.J. Chemopreventive effects of anthocyanins on colorectal and breast cancer: A review. Semin. Cancer Biol. 2022, 81, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Lien, E.C.; Dibble, C.C.; Toker, A. PI3K signaling in cancer: Beyond AKT. Curr. Opin. Cell Biol. 2017, 45, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Sinnett-Smith, J.; Young, S.H.; Rozengurt, E. PKD1 mediates negative feedback of PI3K/Akt activation in response to G protein-coupled receptors. PLoS ONE 2013, 8, e73149. [Google Scholar] [CrossRef] [PubMed]
- Bishnupuri, K.S.; Alvarado, D.M.; Khouri, A.N.; Shabsovich, M.; Chen, B.; Dieckgraefe, B.K.; Ciorba, M.A. IDO1 and kynurenine pathway metabolites activate PI3K-Akt signaling in the neoplastic colon epithelium to promote cancer cell proliferation and inhibit apoptosis. Cancer Res. 2019, 79, 1138–1150. [Google Scholar] [CrossRef]
- Lian, G.; Chen, S.; Ouyang, M.; Li, F.; Chen, L.; Yang, J. Colon cancer cell secretes EGF to promote M2 polarization of TAM through EGFR/PI3K/AKT/mTOR pathway. Technol. Cancer Res. Treat. 2019, 18, 1533033819849068. [Google Scholar] [CrossRef]
- Song, G.; Xu, S.; Zhang, H.; Wang, Y.; Xiao, C.; Jiang, T.; Wu, L.; Zhang, T.; Sun, X.; Zhong, L.; et al. TIMP1 is a prognostic marker for the progression and metastasis of colon cancer through FAK-PI3K/AKT and MAPK pathway. J. Exp. Clin. Cancer Res. 2016, 35, 148. [Google Scholar] [CrossRef]
- Li, G.; Zhang, C.; Liang, W.; Zhang, Y.; Shen, Y.; Tian, X. Berberine regulates the Notch1/PTEN/PI3K/AKT/mTOR pathway and acts synergistically with 17-AAG and SAHA in SW480 colon cancer cells. Pharm. Biol. 2021, 59, 21–30. [Google Scholar] [CrossRef]
- Han, Y.; Peng, Y.; Fu, Y.; Cai, C.; Guo, C.; Liu, S.; Li, Y.; Chen, Y.; Shen, E.; Long, K.; et al. MLH1 deficiency induces cetuximab resistance in colon cancer via Her-2/PI3K/AKT signaling. Adv. Sci. 2020, 7, 2000112. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Mun, J.G.; Jeon, H.D.; Kee, J.Y.; Hong, S.H. Betulin inhibits lung metastasis by inducing cell cycle arrest, autophagy, and apoptosis of metastatic colorectal cancer cells. Nutrients 2019, 12, 66. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, K.-J.; Sung, J.H.; Nam, M.; Suh, K.J.; Kim, J.-W.; Kim, S.H.; Kim, J.W.; Kim, Y.J.; Lee, K.-W.; et al. PI3K p110α blockade enhances anti-tumor efficacy of abemaciclib in human colorectal cancer cells. Cancers 2020, 12, 2500. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, G.; Zhao, L.; Dai, S.; Han, J.; Hu, X.; Zhou, C.; Wang, F.; Ma, H.; Li, B.; et al. Periplocymarin induced colorectal cancer cells apoptosis via impairing PI3K/AKT pathway. Front. Oncol. 2021, 11, 753598. [Google Scholar] [CrossRef]
- Tan, X.; Gong, L.; Li, X.; Zhang, X.; Sun, J.; Luo, X.; Wang, Q.; Chen, J.; Xie, L.; Han, S. Promethazine inhibits proliferation and promotes apoptosis in colorectal cancer cells by suppressing the PI3K/AKT pathway. Biomed. Pharmacother. 2021, 143, 112174. [Google Scholar] [CrossRef]
- Shirakami, Y.; Shimizu, M. Possible mechanisms of green tea and its constituents against cancer. Molecules 2018, 23, 2284. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Li, X.; Ho, C.T.; Lin, X.; Zhang, Y.; Li, B.; Chen, Z. Cocoa tea (Camellia ptilophylla) induces mitochondria-dependent apoptosis in HCT116 cells via ROS generation and PI3K/Akt signaling pathway. Food Res. Int. 2020, 129, 108854. [Google Scholar] [CrossRef]
- Wu, W.; Tang, P.; Ding, B. Research on the culture and technology of Meizhou Hakka green stir-frying. Mod. Hortic. 2018, 5, 170–171. [Google Scholar] [CrossRef]
- Hu, D.; Chen, W.; Ma, C.; Fang, Z.; Huang, H.; Miao, A. Effect of High-temperature and Long-time Final Roasting on Aroma of Roasted Green Tea. Food Sci. 2022, 43, 255–261. [Google Scholar] [CrossRef]
- Li, Q.; Lai, X.; Sun, L.; Cao, J.; Ling, C.; Zhang, W.; Xiang, L.; Chen, R.; Li, D.; Sun, S. Antiobesity and anti-inflammation effects of Hakka stir-fried tea of different storage years on high-fat diet-induced obese mice model via activating the AMPK/ACC/CPT1 pathway. Food Nutr. Res. 2020, 64, 1861. [Google Scholar] [CrossRef]
- Bocchetti, M.; Ferraro, M.G.; Ricciardiello, F.; Ottaiano, A.; Luce, A.; Cossu, A.M.; Scrima, M.; Leung, W.-Y.; Abate, M.; Stiuso, P.; et al. The role of microRNAs in development of colitis-associated colorectal cancer. Int. J. Mol. Sci. 2021, 22, 3967. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shen, X. Long noncoding RNAs: Functions and mechanisms in colon cancer. Mol. Cancer 2020, 19, 167. [Google Scholar] [CrossRef] [PubMed]
- Gelibter, A.J.; Caponnetto, S.; Urbano, F.; Emiliani, A.; Scagnoli, S.; Sirgiovanni, G.; Napoli, V.M.; Cortesi, E. Adjuvant chemotherapy in resected colon cancer: When, how and how long? Surg. Oncol. 2019, 30, 100–107. [Google Scholar] [CrossRef]
- Schmitt, M.; Greten, F.R. The inflammatory pathogenesis of colorectal cancer. Nat. Rev. Immunol. 2021, 21, 653–667. [Google Scholar] [CrossRef]
- Wada, K.; Oba, S.; Tsuji, M.; Goto, Y.; Mizuta, F.; Koda, S.; Uji, T.; Hori, A.; Tanabashi, S.; Matsushita, S.; et al. Green tea intake and colorectal cancer risk in Japan: The Takayama study. Jpn. J. Clin. Oncol. 2019, 49, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.W.; Xia, J.; Cheng, B.H.; Gao, H.C.; Fu, L.W.; Luo, X.L. Tea polyphenol EGCG inhibited colorectal-cancer-cell proliferation and migration via downregulation of STAT3. Gastroenterol. Rep. 2021, 9, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Uesato, S.; Kitagawa, Y.; Kamishimoto, M.; Kumagai, A.; Hori, H.; Nagasawa, H. Inhibition of green tea catechins against the growth of cancerous human colon and hepatic epithelial cells. Cancer Lett. 2001, 170, 41–44. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; Zhang, J.; Zhu, J.; Liu, P.; Xu, L.; Wei, K.; Zhou, H.; Peng, L.; Zhang, J.; et al. Dynamic changes of metabolic profile and taste quality during the long-term aging of Qingzhuan Tea: The impact of storage age. Food Chem. 2021, 359, 129953. [Google Scholar] [CrossRef]
- Huang, A.; Jiang, Z.; Tao, M.; Wen, M.; Xiao, Z.; Zhang, L.; Zha, M.; Chen, J.; Liu, Z.; Zhang, L. Targeted and nontargeted metabolomics analysis for determining the effect of storage time on the metabolites and taste quality of keemun black tea. Food Chem. 2021, 359, 129950. [Google Scholar] [CrossRef]
- Li, F.; Lu, Q.; Li, M.; Yang, X.; Xiong, C.; Yang, B. Comparison and risk assessment for trace heavy metals in raw Pu-erh tea with different storage years. Biol. Trace Elem. Res. 2020, 195, 696–706. [Google Scholar] [CrossRef]
- Xu, P.; Chen, L.; Wang, Y. Effect of storage time on antioxidant activity and inhibition on α-Amylase and α-Glucosidase of white tea. Food Sci. Nutr. 2019, 7, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhou, Y.; Zhao, J.; Yao, S.; Wang, J. Effect of storage time on biochemical characteristics and antioxidant activity of hawk tea (Litsea coreana) processed by boiling water fixation. Food Sci. Nutr. 2020, 8, 6182–6191. [Google Scholar] [CrossRef] [PubMed]
- Jeng, K.C.; Chen, C.S.; Fang, Y.P.; Hou, R.C.; Chen, Y.S. Effect of microbial fermentation on content of statin, GABA, and polyphenols in Pu-Erh tea. J. Agric. Food Chem. 2007, 55, 8787–8792. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.P.; Jaganathan, S.K.; Mandal, M.; Supriyanto, E.; Muhamad, I.I. Gallic acid induced apoptotic events in HCT-15 colon cancer cells. World J. Gastroenterol. 2016, 22, 3952–3961. [Google Scholar] [CrossRef]
- Zhou, L.A.; Liu, T.B.; Lü, H.N. Geraniin inhibits proliferation and induces apoptosis through inhibition of phosphatidylinositol 3-kinase/Akt pathway in human colorectal cancer in vitro and in vivo. Anticancer Drugs 2020, 31, 575–582. [Google Scholar] [CrossRef]
- Tan, X.; Hu, D.; Li, S.; Han, Y.; Zhang, Y.; Zhou, D. Differences of four catechins in cell cycle arrest and induction of apoptosis in LoVo cells. Cancer Lett. 2000, 158, 1–6. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, L.; Li, C.; Chen, R.; Liu, C.; Chen, M. Lipophilized epigallocatechin gallate derivative exerts anti-proliferation efficacy through induction of cell cycle arrest and apoptosis on DU145 human prostate cancer cells. Nutrients 2019, 12, 92. [Google Scholar] [CrossRef]
- Du, B.X.; Lin, P.; Lin, J. EGCG and ECG induce apoptosis and decrease autophagy via the AMPK/mTOR and PI3K/AKT/mTOR pathway in human melanoma cells. Chin. J. Nat. Med. 2022, 20, 290–300. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | 03Y | 07Y | 11Y | 15Y | 19Y |
|---|---|---|---|---|---|
| Tea polyphenols/% | 39.26 ± 0.24 b | 32.39 ± 0.33 c | 41.65 ± 0.09 a | 39.91 ± 0.22 ab | 38.27 ± 0.01 b |
| Free amino acid/% | 5.35 ± 0.04 ab | 4.40 ± 0.00 bc | 4.19 ± 0.02 c | 5.01 ± 0.08 b | 5.51 ± 0.09 a |
| Ratio of TP to FAA | 7.44 ± 0.06 c | 7.38 ± 0.30 c | 9.93 ± 0.05 a | 8.14 ± 0.06 b | 7.04 ± 0.01 c |
| Soluble sugar/% | 26.84 ± 0.05 a | 23.69 ± 0.62 b | 24.34 ± 0.53 b | 23.64 ± 0.26 b | 26.65 ± 0.29 a |
| Flavonoids/% | 2.60 ± 0.04 d | 3.01 ± 0.03 b | 3.25 ± 0.05 a | 2.85 ± 0.03 c | 2.59 ± 0.01 d |
| Ingredients (mg/g) | 03Y | 07Y | 11Y | 15Y | 19Y |
|---|---|---|---|---|---|
| CG | 2.64 ± 0.05 a | 1.67 ± 0.04 ab | 1.65 ± 0.14 ab | 0.76 ± 0.04 b | 1.08 ± 0.08 ab |
| ECG | 29.73 ± 0.96 b | 33.74 ± 0.37 ab | 33.94 ± 0.81 a | 30.34 ± 1.18 b | 26.51 ± 0.21 b |
| GCG | 16.47 ± 0.39 a | 10.45 ± 0.48 b | 11.44 ± 0.21 b | 11.13 ± 0.51 b | 12.24 ± 0.24 b |
| EGCG | 137.55 ± 1.04 a | 141.79 ± 0.04 a | 146.77 ± 0.85 a | 131.21 ± 0.98 a | 131.20 ± 0.63 a |
| EC | 9.65 ± 0.11 b | 9.31 ±0.25 b | 12.41 ± 0.21 a | 13.52 ± 0.47 a | 14.37 ± 0.15 a |
| C | 6.17 ± 0.03 a | 4.48 ± 0.13 ab | 6.49 ± 0.02 a | 5.46 ± 0.14 ab | 4.35 ± 0.26 b |
| GC | 69.02 ± 0.00 b | 86.00 ± 1.28 a | 51.71 ± 0.85 c | 52.05 ± 1.87 c | 44.13 ± 0.73 c |
| EGC | 38.65 ± 1.25 c | 33.02 ± 0.77 c | 45.03 ± 0.09 b | 58.59 ± 1.82 a | 63.67 ± 0.36 a |
| GA | 10.88 ± 0.03 a | 9.79 ± 0.17 b | 6.40 ± 0.22 c | 5.54 ± 0.33 c | 3.45 ± 0.03 d |
| CAFF | 92.67 ± 0.25 a | 90.45 ± 1.47 a | 75.85 ± 0.10 b | 88.99 ± 1.06 ab | 71.96 ± 0.01 b |
| Ester catechins | 185.59 ± 4.96 a | 184.01 ± 2.66 a | 180.13 ± 7.42 a | 188.57 ± 7.20 a | 169.85 ± 2.02 a |
| Non-ester catechins | 120.91 ± 2.13 a | 132.37 ± 0.72 a | 115.26 ± 0.15 a | 129.62 ± 4.30 a | 128.21 ± 2.61 a |
| Total catechins | 306.50 ± 5.45 a | 328.23 ± 2.40 a | 308.18 ± 15.38 a | 303.29 ± 11.49 a | 298.06 ± 4.89 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Huang, H.; Sun, S.; Li, D.; Sun, L.; Li, Q.; Chen, R.; Lai, X.; Zhang, Z.; Zheng, X.; et al. Induction of Apoptosis via Inactivating PI3K/AKT Pathway in Colorectal Cancer Cells Using Aged Chinese Hakka Stir-Fried Green Tea Extract. Molecules 2022, 27, 8272. https://doi.org/10.3390/molecules27238272
Zhang X, Huang H, Sun S, Li D, Sun L, Li Q, Chen R, Lai X, Zhang Z, Zheng X, et al. Induction of Apoptosis via Inactivating PI3K/AKT Pathway in Colorectal Cancer Cells Using Aged Chinese Hakka Stir-Fried Green Tea Extract. Molecules. 2022; 27(23):8272. https://doi.org/10.3390/molecules27238272
Chicago/Turabian StyleZhang, Xinyue, Haiying Huang, Shili Sun, Dongli Li, Lingli Sun, Qiuhua Li, Ruohong Chen, Xingfei Lai, Zhenbiao Zhang, Xi Zheng, and et al. 2022. "Induction of Apoptosis via Inactivating PI3K/AKT Pathway in Colorectal Cancer Cells Using Aged Chinese Hakka Stir-Fried Green Tea Extract" Molecules 27, no. 23: 8272. https://doi.org/10.3390/molecules27238272
APA StyleZhang, X., Huang, H., Sun, S., Li, D., Sun, L., Li, Q., Chen, R., Lai, X., Zhang, Z., Zheng, X., Wong, W.-L., & Wen, S. (2022). Induction of Apoptosis via Inactivating PI3K/AKT Pathway in Colorectal Cancer Cells Using Aged Chinese Hakka Stir-Fried Green Tea Extract. Molecules, 27(23), 8272. https://doi.org/10.3390/molecules27238272