
Allelopathic Toxicity of Cyanamide Could Control Amaranth (Amaranthus retroflexus L.) in Alfalfa (Medicago sativa L.) Field

Abstract
1. Introduction
2. Results and Discussion
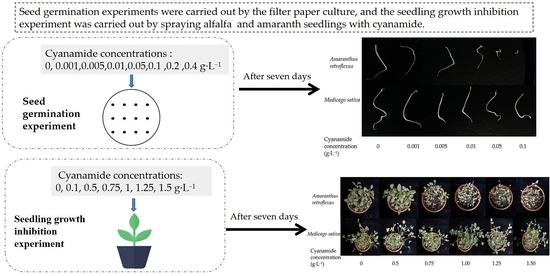
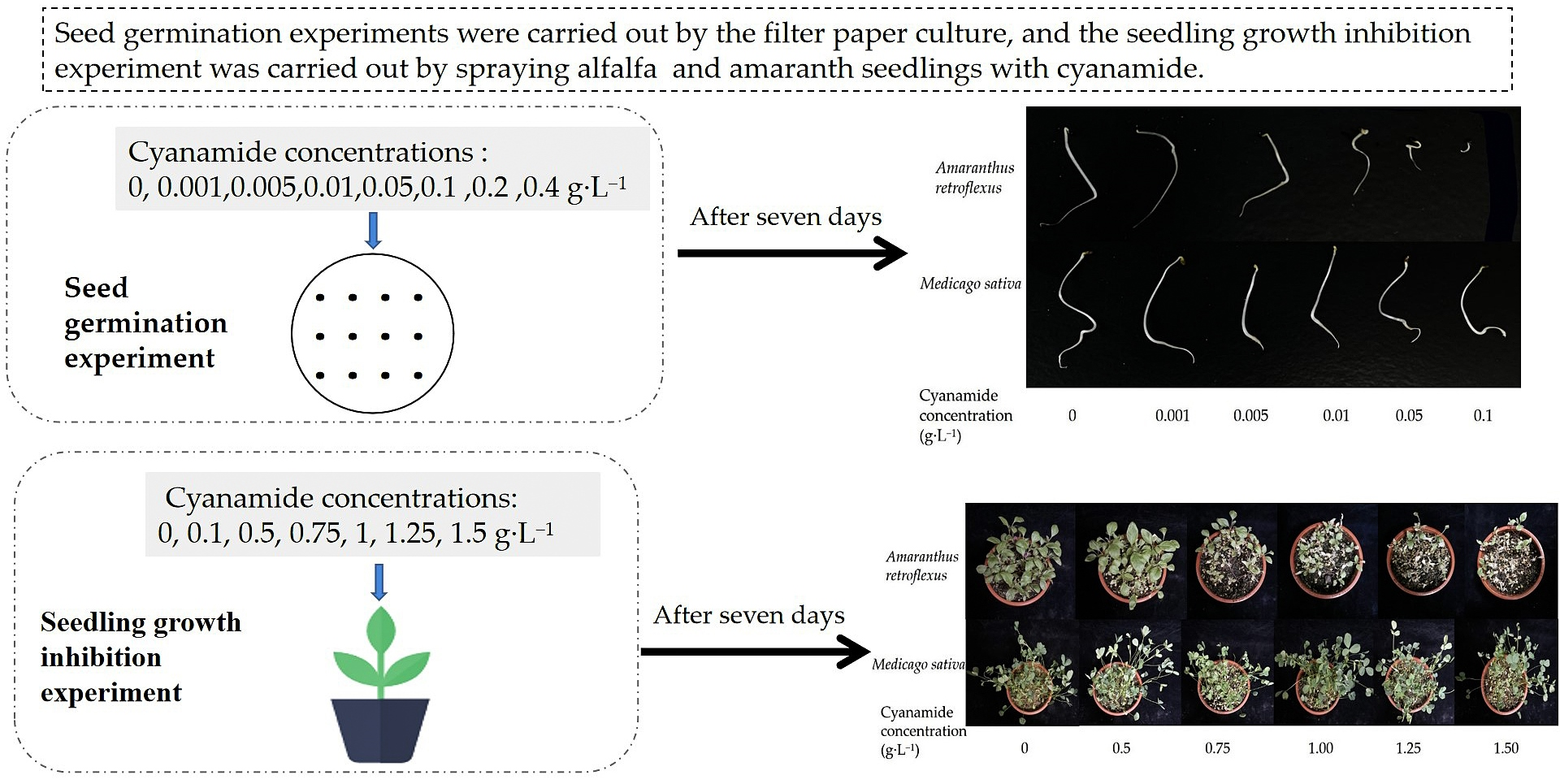
2.1. Effects of CA on Seed Germination
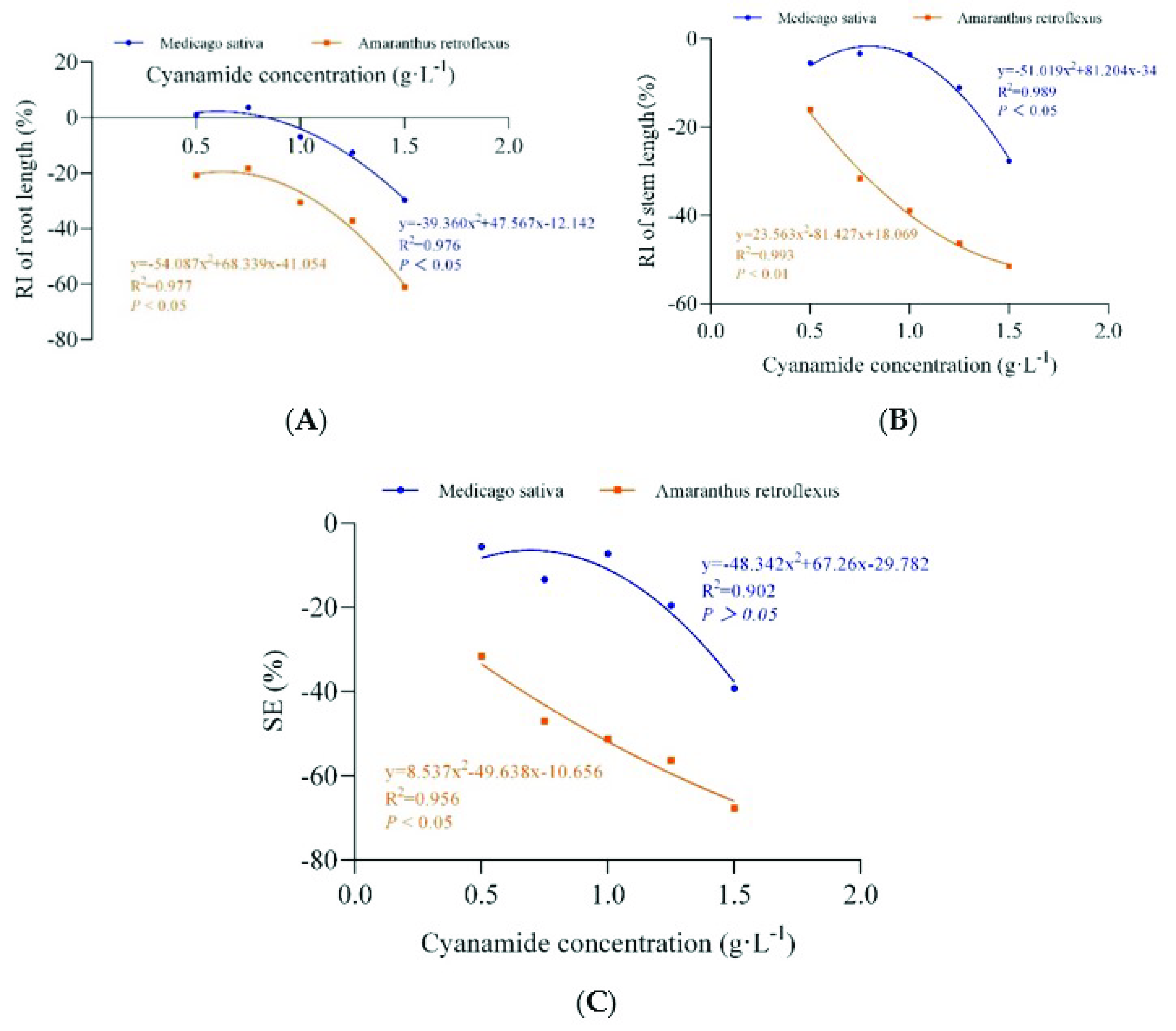
2.2. Effect of CA on Seedling Growth
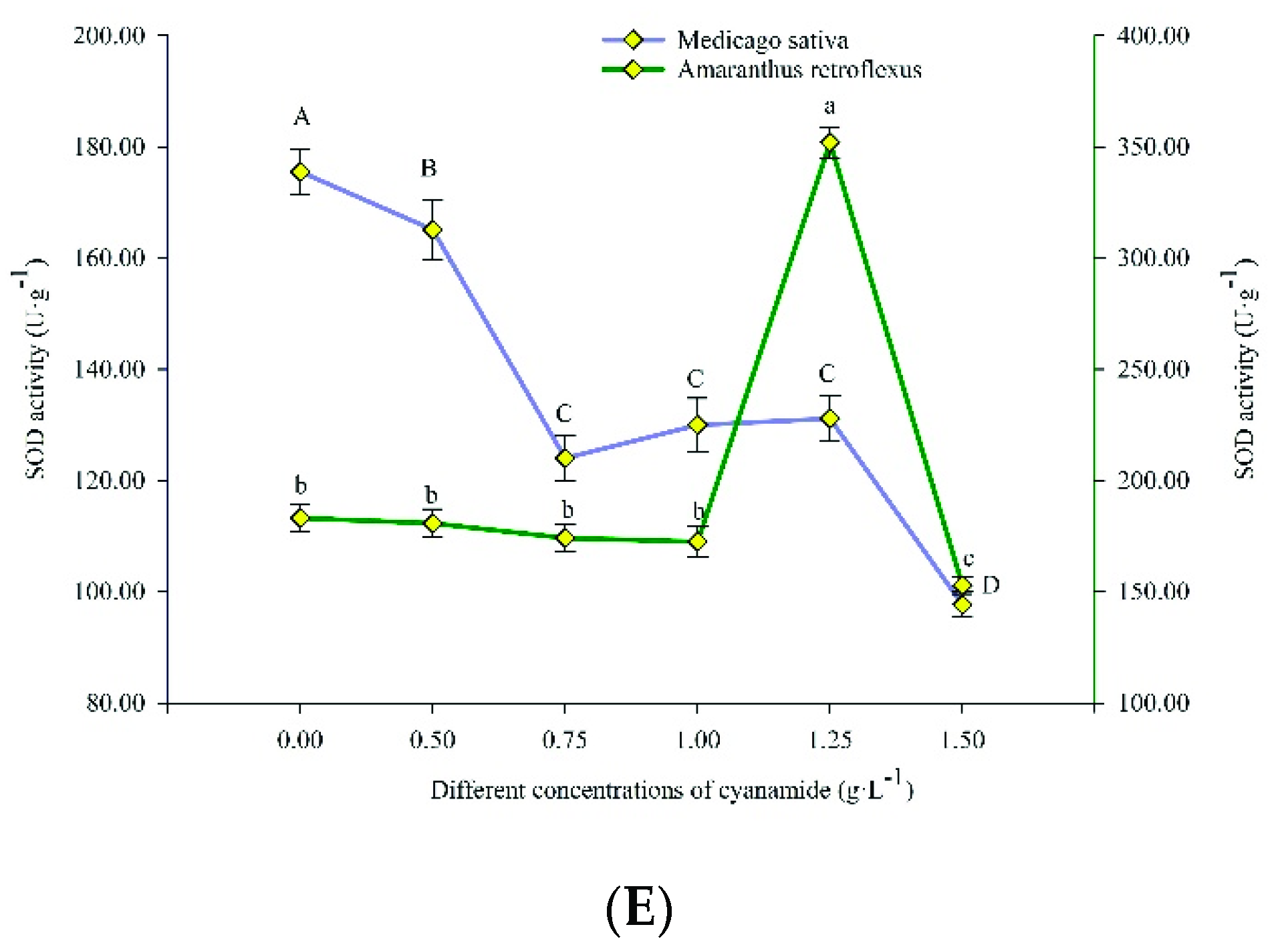
2.3. Effects of CA on Activation of Antioxidant Defensive Enzymes
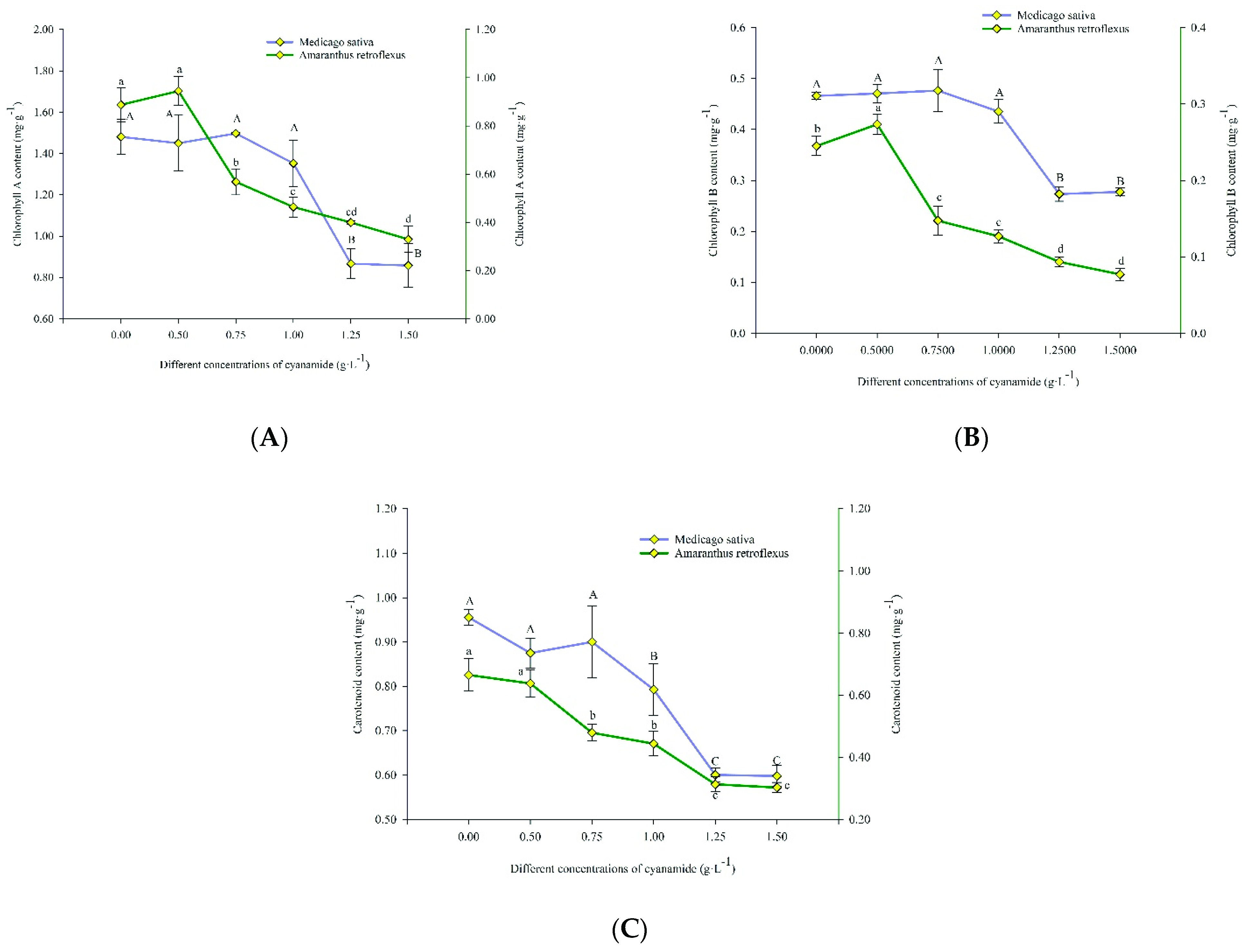
2.4. Effects on Photosynthetic Activities
3. Materials and Methods
3.1. Experimental Materials
3.2. Seeds Germination Assays
3.3. Pot Experiment
3.4. Antioxidant Enzyme Activities
3.5. Concentrations of Chl-a, Chl-b, Carotenoids
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Diatta, A.A.; Min, D.; Jagadish, K.S. Drought stress responses in non-transgenic and transgenic alfalfa—Current status and future research directions. Adv. Agron. 2021, 170, 2–66. [Google Scholar]
- Dillehay, B.L.; Curran, W.S. Comparison of herbicide programs for weed control in glyphosate-resistant alfalfa. Weed Technol. 2010, 24, 130–138. [Google Scholar] [CrossRef]
- Sheaffer, C.C.; Martinson, K.M.; Wyse, D.L.; Moncada, K.M. Companion crops for organic alfalfa establishment. Agron. J. 2014, 106, 309–314. [Google Scholar] [CrossRef]
- Mohammadi, G.; Norouzi, Y.; Nosratti, I. The response of three weed species to sorghum, alfalfa and tobacco plant tissues incorporated into the soil. Philipp. Agric. Sci. 2016, 99, 105–112. [Google Scholar]
- Evon, P.; Langalerie, G.D.; Ramaux, T.; Labonne, L.; Ouagne, P. Amaranth, a model for the future biorefinery of whole plants. Symbiose Flower Conf. Nantes 2019, 14, 1–2. [Google Scholar]
- Orłowski, G.; Czarnecka, J. Granivory of birds and seed dispersal: Viable seeds of Amaranthus retroflexus L. recovered from the droppings of the Grey Partridge Perdix perdix L. Pol. J. Ecol. 2009, 57, 191–196. [Google Scholar]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Neve, P. Modeling the simultaneous evolution of resistance to ALS- and ACCase-inhibiting herbicides in barnyardgrass (Echinochloa crus-galli) in clearfield rice. Weed Technol. 2014, 28, 89–103. [Google Scholar] [CrossRef]
- Li, N.; Lee, H.K. Sample preparation based on dynamic ion-exchange solid-phase extraction for GC/MS analysis of acidic herbicides in environmental waters. Anal. Chem. 2000, 72, 3077–3084. [Google Scholar] [CrossRef]
- Kughur, P.G. The effects of herbicides on crop production and environment in Makurdi Local Government area of benu State. Nigeria 2012, 14, 35–46. [Google Scholar]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Dawra, G.P. Potential of allelopathy and allelochemicals for weed management. In Handbook of Sustainable Weed Management; CAB Direct: Binghamton, NY, USA, 2006; Volume 32, pp. 136–148. [Google Scholar]
- Dokoozlian, N.K.; Williams, L.E.; Neja, R.A. Chilling exposure and hydrogen cyanamide interact in breaking dormancy of grape buds. Hortence Publ. Am. Soc. Hortic. Ence 1995, 30, 1244–1247. [Google Scholar] [CrossRef]
- Kamo, T.; Hiradate, S.; Fujii, Y. First isolation of natural cyanamide as a possible allelochemical from hairy vetch Vicia villosa. J. Chem. Ecol. 2003, 29, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Soltys, D.; Rudzińska-Langwald, A.; Kurek, W.; Szajko, K.; Sliwinska, E.; Bogatek, R.; Gniazdowska, A. Phytotoxic cyanamide affects maize (Zea mays) root growth and root tip function: From structure to gene expression. J. Plant Physiol. 2014, 171, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Steller, W.A.; Morgan, P.W. Determination of cyanamide residues on ginned cottonseed. J. Agric. Food Chem. 1965, 13, 329–330. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Ismail, B.S.; Tan, P.W.; Chuah, T.S. Assessment of the potential allelopathic effects of Pennisetum purpureum Schumach. on the germination and growth of Eleusine indica (L.) Gaertn. Sains Malays. 2015, 44, 269–274. [Google Scholar] [CrossRef]
- Wu, A.P.; Huang, Z.Y.; Miao, S.L.; Dong, M. Effects of Mikania micrantha extracts and their exposure time on seed vigor, seed germination and seedling growth of plants. Allelopath. J. 2010, 25, 502–512. [Google Scholar]
- Hussain, M.I.; Gonzlez, L.; Reigosa, M.J. Allelopathic potential of Acacia melanoxylon on the germination and root growth of native species. Weed Biol. Manag. 2011, 11, 18–28. [Google Scholar] [CrossRef]
- Maninang, J.S.; Okazaki, S.; Fujii, Y. Cyanamide phytotoxicity in soybean (Glycine max) seedlings involves aldehyde dehydrogenase inhibition and oxidative stress. Nat. Prod. Commun. 2015, 10, 743–746. [Google Scholar] [CrossRef]
- Soltys, D.; Rudzińska-Langwald, A.; Kurek, W.; Gniazdowska, A.; Sliwinska, E.; Bogatek, R. Cyanamide mode of action during inhibition of onion (Allium cepa L.) root growth involves disturbances in cell division and cytoskeleton formation. Planta 2011, 234, 609–621. [Google Scholar] [CrossRef]
- Soltys, D.; Rudzińska-Langwald, A.; Gniazdowska, A.; Wi Niewska, A.; Bogatek, R. Inhibition of tomato (Solanum lycopersicum L.) root growth by cyanamide is due to altered cell division, phytohormone balance and expansin gene expression. Planta 2012, 236, 1629–1638. [Google Scholar] [CrossRef]
- Burgos, N.R.; Talbert, R.E.; Kim, K.S.; Kuk, Y.I. Growth inhibition and root ultrastructure of cucumber seedlings exposed to allelochemicals from rye (Secale cereale). J. Chem. Ecol. 2004, 30, 671–689. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, A.R. Two distinct sources of elicited reactive oxygen species in tobacco epidermal cells. Plant Cell Online 1997, 9, 1559–1572. [Google Scholar]
- Pellizzon, C.H. The use of menthol in skin wound healing—Anti-inflammatory potential, antioxidant defense system stimulation and increased epithelialization. Pharmaceutics 2021, 13, 11–24. [Google Scholar]
- Jie, Z.; Min, Z.; Shengke, T.; Lingli, L.; Jis, M.; Xiaoe, Y. The activities of SOD, POD CAT and the content of H2O2 in the shoot and roots of wild-type and transgenic tobacco plants. PLoS ONE 2014, 17, 142–152. [Google Scholar]
- Yukinori, Y.; Takanori, M.; Kazuya, Y.; Takahiro, I.; Shigeru, S. Two distinct redox signaling pathways for cytosolic APX induction under photooxidative stress. Plant Cell Physiol. 2004, 45, 1586–1594. [Google Scholar]
- Pirinccioglu, A.G.Z.; Kalp, G.D.; Pirinccioglu, M.; Kizil, G.K.; Kizil, M. Malondialdehyde (MDA) and protein carbonyl (PCO) levels as biomarkers of oxidative stress in subjects with familial hypercholesterolemia. Clin. Biochem. 2010, 43, 1220–1224. [Google Scholar] [CrossRef]
- Devi, S.R.; Prasad, M. Ferulic acid mediated changes in oxidative enzymes of maize seedlings: Implications in growth. Biol. Plant. 1996, 38, 387–395. [Google Scholar] [CrossRef]
- Etinski, M.; Petkovic, M.; Ristic, M. A study of the low-lying singlet and triplet electronic states of chlorophyll a and b. J. Serb. Chem. Soc. 2013, 78, 1775–1787. [Google Scholar] [CrossRef]
- Elisante, F.; Tarimo, M.T.; Ndakidemi, P.A. Allelopathic effect of seed and leaf aqueous extracts of Datura stramonium on leaf chlorophyll content, shoot and root elongation of Cenchrus ciliaris and Neonotonia wightii. Am. J. Plant Sci. 2013, 4, 2332–2339. [Google Scholar] [CrossRef]
- Ashrafi, Z.Y.; Rahnavard, A.; Sadeghi, S. Allelopathic potential of sunflower (Helianthus annuus) against seed germination in wild mustard (Sinapis arvensis) and foxtail (Setaria viridis). Indian J. Weed Sci. 2010, 42, 82–87. [Google Scholar]
- Edwards, M.S. Using in situ substratum sterilization and fluorescence microscopy in studies of microscopic stages of marine macroalgae. Hydrobiologia 1999, 398, 253–259. [Google Scholar]
- Ladhari, A.; Andolfi, A.; Dellagreca, M. Physiological and oxidative stress responses of lettuce to cleomside A: A thiohydroximate, as a new allelochemical from Cleome arabica L. Molecules 2020, 25, 4461. [Google Scholar] [CrossRef]
- Ramachandran, S.; Asokkumar, K.; Maheswari, M.U.; Ravi, T.K.; Dharman, J. Investigation of antidiabetic, antihyperlipidemic, and in vivo antioxidant properties of Sphaeranthus indicus Linn. in type 1 diabetic rats: An identification of possible biomarkers. Evid. Based Complementray Altern. Med. 2021, 8, 1741–4271. [Google Scholar]
- Cakmak, I.; Marschner, H. Enhanced superoxide radical production in roots of zinc-deficient plants. J. Exp. Bot. 1988, 39, 1449–1460. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyanamide Concentration (g·L−1) | Germination Rate (%) | RI (%) | Radicle Length (cm) | RI (%) | Hypocotyl Length (cm) | RI (%) | Fresh Weight (g) | RI (%) | Dry Weight (g) | RI (%) | SE (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amaranthus retroflexus | 0 | 47.33 ± 4.16 a | 1.45 ± 0.14 a | 2.01 ± 0.36 a | 0.0033 ± 0.00098 a | 0.0004 ± 0.00015 a | ||||||
| 0.001 | 49.33 ± 9.02 a | 4.23 | 1.61 ± 0.17 a | 11.16 | 0.86 ± 0.08 b | −57.09 | 0.0027 ± 0.0002 ab | −18.18 | 0.0001 ± 0.00001 ab | −75.00 | −14.97 | |
| 0.005 | 54.00 ± 7.21 a | 14.08 | 1.50 ± 0.23 a | 3.80 | 0.65 ± 0.11c | −67.85 | 0.0021 ± 0.00006 bc | −36.36 | 0.0001 ± 0.00002 ab | −75.00 | −21.58 | |
| 0.01 | 53.33 ± 10.07 a | 12.68 | 0.96 ± 0.16 b | −33.72 | 0.56 ± 0.11 c | −72.25 | 0.0024 ± 0.0001 b | −27.27 | 0.0002 ± 0.00008 b | −50.00 | −30.14 | |
| 0.05 | 17.33 ± 6.11 b | −63.38 | 0.39 ± 0.08 c | −72.84 | 0.36 ± 0.05 d | −82.02 | 0.0014 ± 0.00046 c | −57.58 | 0.0001 ± 0.00001 c | −75.00 | −68.96 | |
| 0.1 | 0.00 ± 0.00 c | −100 | 0.1 ± 0.09 d | −93.10 | 0.00 ± 0.00 e | −100 | 0.00 ± 0.00 d | −100 | 0.00 ± 0.00 c | −100.00 | −98.27 | |
| Medicago sativa | 0 | 93.33 ± 1.15 A | 1.55 ± 0.12 A | 4.55 ± 0.18 A | 0.0245 ± 0.0015 A | 0.0014 ± 0.00010 A | ||||||
| 0.001 | 91.33 ± 3.06 A | −2.14 | 1.52 ± 0.13 AB | −1.94 | 4.32 ± 0.12 B | −5.06 | 0.0257 ± 0.0016 A | 4.90 | 0.0014 ± 0.00021 A | 0.00 | −0.85 | |
| 0.005 | 87.33 ± 4.16 A | −6.43 | 1.35 ± 0.11 BC | −12.73 | 3.51 ± 0.19 C | −19.09 | 0.0257 ± 0.0012 A | 4.90 | 0.0014 ± 0.00025 A | 0.00 | −6.67 | |
| 0.01 | 87.33 ± 3.06 A | −6.43 | 1.53 ± 0.21 AB | −0.76 | 4.22 ± 0.13 B | −7.29 | 0.0242 ± 0.0021 A | −1.22 | 0.0014 ± 0.00035 A | 0.00 | −3.14 | |
| 0.05 | 88.67 ± 2.31 A | −5.00 | 1.36 ± 0.10 ABC | −12.19 | 4.30 ± 0.19 B | −5.42 | 0.0240 ± 0.0018 A | −2.04 | 0.0013 ± 0.00015 A | −7.14 | −6.36 | |
| 0.1 | 87.33 ± 4.16 A | −6.43 | 1,32 ± 0.18 C | −14.56 | 3.72 ± 0.28 C | −18.28 | 0.02.7 ± 0.0014 B | −15.51 | 0.0013 ± 0.00026 A | −7.14 | −12.39 |
| Cyanamide Concentration (g·L−1) | Root Length (cm) | RI (%) | Stem Length (cm) | RI (%) | Fresh Weight (g) | RI (%) | Dry Weight (g) | RI (%) | SE (%) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Amaranthus retroflexus | 0 | 4.018 ± 0.518 a | 7.533 ± 0.345 a | 0197 ± 0.010 a | 0.067 ± 0.002 a | |||||
| 0.50 | 3.183 ± 0.249 b | −20.78 | 6.328 ± 0.201 b | −16.00 | 0.101 ± 0.007 b | −48.40 | 0.040 ± 0.006 b | −40.90 | −31.52 | |
| 0.75 | 3.285 ± 0.530 b | −18.25 | 5.155 ± 0.224 c | −31.57 | 0.051 ± 0.007 c | −73.89 | 0.024 ± 0.003 c | −63.88 | −46.90 | |
| 1.00 | 2.793 ± 0.630 bc | −30.49 | 4.605 ± 0.257 d | −38.87 | 0.058 ± 0.004 c | −70.48 | 0.023 ± 0.002 c | −65.07 | −51.23 | |
| 1.25 | 2.523 ± 0.303 c | −37.20 | 4.045 ± 0.240 e | −46.31 | 0.059 ± 0.009 c | −70.13 | 0.019 ± 0.002 d | −71.34 | −56.25 | |
| 1.50 | 1.563 ± 0.391 d | −61.10 | 3.653 ± 0.377 f | −51.50 | 0.034 ± 0.007 d | −82.95 | 0.017 ± 0.002 d | −74.63 | −67.55 | |
| Medicago sativa | 0 | 10.850 ± 0.718 A | 12.720 ± 0.760 A | 0.430 ± 0.036 A | 0.054 ± 0.003 A | |||||
| 0.50 | 10.973 ± 0.436 A | 1.14 | 12.028 ± 0.766 AB | −5.44 | 0.350 ± 0.024 A | −18.53 | 0.055 ± 0.005 A | 0.92 | −5.90 | |
| 0.75 | 11.252 ± 0.468 A | 3.70 | 12.298 ± 0.804 AB | −3.32 | 0.221 ± 0.018 A | −48.58 | 0.052 ± 0.004 A | −4.96 | −13.29 | |
| 1.00 | 10.105 ± 0.324 B | −6.87 | 12.270 ± 0.730 AB | −3.54 | 0.376 ± 0.033 A | −12.63 | 0.051 ± 0.003 A | −5.51 | −7.14 | |
| 1.25 | 9.485 ± 0.603 B | −12.58 | 11.313 ± 0.937 B | −11.06 | 0.311 ± 0.027 B | −27.70 | 0.040 ± 0.003 B | −26.47 | −19.45 | |
| 1.50 | 7.632 ± 0.539 C | −29.66 | 9.208 ± 0.779 C | −27.61 | 0.210 ± 0.010 C | −51.28 | 0.028 ± 0.000 C | −48.16 | −39.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Yang, C.; Shan, X.; An, M.; Wang, X. Allelopathic Toxicity of Cyanamide Could Control Amaranth (Amaranthus retroflexus L.) in Alfalfa (Medicago sativa L.) Field. Molecules 2022, 27, 7347. https://doi.org/10.3390/molecules27217347
Sun W, Yang C, Shan X, An M, Wang X. Allelopathic Toxicity of Cyanamide Could Control Amaranth (Amaranthus retroflexus L.) in Alfalfa (Medicago sativa L.) Field. Molecules. 2022; 27(21):7347. https://doi.org/10.3390/molecules27217347
Chicago/Turabian StyleSun, Weihong, Chaowei Yang, Xinhe Shan, Mingzhu An, and Xianguo Wang. 2022. "Allelopathic Toxicity of Cyanamide Could Control Amaranth (Amaranthus retroflexus L.) in Alfalfa (Medicago sativa L.) Field" Molecules 27, no. 21: 7347. https://doi.org/10.3390/molecules27217347
APA StyleSun, W., Yang, C., Shan, X., An, M., & Wang, X. (2022). Allelopathic Toxicity of Cyanamide Could Control Amaranth (Amaranthus retroflexus L.) in Alfalfa (Medicago sativa L.) Field. Molecules, 27(21), 7347. https://doi.org/10.3390/molecules27217347