Abstract
Kahweol and cafestol are two diterpenes extracted from Coffea arabica beans that have distinct biological activities. Recent research describes their potential activities, which include anti-inflammatory, anti-diabetic, and anti-cancer properties, among others. The two diterpenes have been shown to have anticancer effects in various in vitro and in vivo cancer models. This review aims to shed light on the recent developments regarding the potential effects of kahweol and cafestol on various cancers. A systematic literature search through Google Scholar and PubMed was performed between February and May 2022 to collect updates about the potential effects of cafestol and kahweol on different cancers in in vitro and in vivo models. The search terms “Kahweol and Cancer” and “Cafestol and Cancer” were used in this literature review as keywords; the findings demonstrated that kahweol and cafestol exhibit diverse effects on different cancers in in vitro and in vivo models, showing pro-apoptotic, cytotoxic, anti-proliferative, and anti-migratory properties. In conclusion, the diterpenes kahweol and cafestol display significant anticancer effects, while remarkably unaffecting normal cells. Our results show that both kahweol and cafestol exert their actions on various cancers via inducing apoptosis and inhibiting cell growth. Additionally, kahweol acts by inhibiting cell migration.
1. Introduction
Coffee is one of the most widely consumed beverages in the world. This may be attributed to its desirable stimulatory effect and distinctive taste. Fortunately, coffee’s constituents were found to have protective functions against many diseases, including type 2 diabetes mellitus, liver cirrhosis, and neurodegenerative disorders [1,2,3,4,5,6]. The physiologically active components of coffee that play this biological role include caffeine, diterpenes, chlorogenic acids, and melanoidins; their biological properties make them a hot topic to investigate.
The diterpenes kahweol and cafestol are fat-soluble compounds derived from Coffee arabica beans [7]. They are structural analogues, with the difference of an additional double bond in the kahweol molecule (Figure 1) [8]. Extraction of kahweol and cafestol can be performed through direct hot saponification (DHS), direct cold saponification (DCS), and Bligh and Dyer (BD) or Soxhlet (SO) extraction followed by saponification [9]. DHS was reported to be quicker, more efficient, and more economical than the other techniques for extracting the diterpenes from coffee. Purification can be performed using a flash chromatography column (FCC) packed with flash silica gel, with hexane/ethyl acetate as the mobile phase [10]. The collected fractions can be further identified by high-performance liquid chromatography with diode array and mass spectrometry detectors (HPLC–DAD–MS/MS) [9]. This method provides a simple characterization of kahweol and cafestol. In addition, it distinguishes the two molecules by their molecular mass.
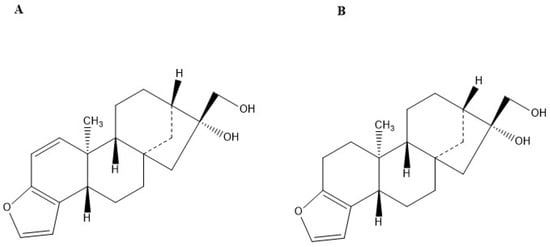
Figure 1.
Chemical structure of (A) Kahweol and (B) Cafestol, created by ChemDraw.
In unfiltered coffee, such as Turkish coffee or espresso, kahweol and cafestol are found in relatively higher concentrations in comparison to filtered and instant coffee. A study performed on healthy ileostomy volunteers showed that, upon coffee consumption, gastric juices break down nearly 30% of kahweol and cafestol, and 64–70% of cafestol and 70–73% of kahweol are absorbed in the small intestine, with minute amounts of glucuronidated or sulphated conjugate forms being excreted in the urine [11]. As only about 1% or less of conjugate cafestol and kahweol is excreted in the urine, the two diterpenes are likely metabolised further in the body, and there is evidence that cafestol circulates in the liver and gastrointestinal tract, exerting its effects [12].
Kahweol and cafestol’s role in raising serum lipids is well-documented. The long-term ingestion of unfiltered coffee has been linked to increased plasma levels of triacylglycerol and low-density lipoprotein (LDL) cholesterol in human subjects, and cafestol and kahweol are the primary coffee constituents responsible for this effect, with cafestol exerting a more potent effect than kahweol [13]. Some in vitro studies showed that cafestol suppressed LDL receptor activity by decreasing its ability to bind, uptake, and degrade LDL [14]. However, LDL levels were also found to be raised in mice with LDL receptor knockdown following cafestol administration [15]. Cafestol suppresses enzymes involved in bile acid synthesis (sterol 27-hydroxylase and cholesterol 7 alpha-hydroxylase), providing an alternate explanation for the rise in cholesterol levels [15].
Emerging evidence has shown that kahweol and cafestol target several pathways to prevent disease. Their anti-inflammatory effect is prominently displayed by their ability to regulate several inflammatory mediators, such as ICAM1, MCP1, and IL-8, which are involved in cardiovascular disease progression [16,17]. In addition, kahweol and cafestol inhibit osteoclast differentiation and bone resorption, while promoting osteoblast differentiation, thus ameliorating degenerative bone diseases [18,19]. Of the many roles the diterpenes play, their anti-carcinogenic effect is perhaps the most significant due to the poor prognosis of many cancers.
The chemopreventive function of diterpenes can be seen in their ability to induce phase 2 detoxifying enzymes and anti-oxidant proteins, which prevents early mutagenic events. Their pro-apoptotic characteristic also contributes to their anti-tumorigenic effects by downregulating anti-apoptotic proteins, such as Bcl-2, Bcl-xL, mcl1, cFLIP, decreasing the release of cytochrome c and activating caspase enzymes [20]. In addition, diterpenes are also inhibitors of angiogenesis, as their introduction to HMVECs (Human Microvascular Endothelial Cells) inhibits cell proliferation and migration [21].
This is the first review focusing on the various effects exerted by kahweol and cafestol on different in vitro and in vivo cancer models and the pathways involved in the eradication of such cancers. The current literature primarily focuses on the general bioactivity and pharmacological effects of these two diterpenes, including their anti-inflammatory, anti-diabetic and anti-osteoclastogenesis activity, with only a cursory description of their anticancer effects [8]. Thus, this review aims to discuss, in detail, the tumour-suppressive role both kahweol and cafestol play in various cancer, through apoptosis, inhibition of proliferation and migration, or induction of cytotoxicity. The biochemical pathways through which these compounds exert their effects on multiple cancers are outlined, and their potential function in cancer treatment is explored.
2. Kahweol and Cafestol Effects on Several Cancer Cell Lines
Kahweol and cafestol exert anticancer effects on several cancer cell lines. The pathways through which they exhibit their properties are discussed in this section. Table 1 and Table 2 summarise the findings.

Table 1.
In Vitro Impact of Kahweol and Cafestol on Cancer Cell Lines (↑ indicates induction, ↓ indicates suppression).
Table 2.
In Vivo Impact of Kahweol and Cafestol on Cancer models (↑ indicates induction, ↓ indicates suppression).
2.1. Lung Cancer
Kahweol and cafestol were tested in vitro against different lung cancer cell lines, and kahweol-induced apoptosis was observed in all cell lines. With regard to mesothelioma, MSTO-211H cells and H28 cells were examined [7]. The apoptosis of cancerous cells was activated by the upregulation of Bax, alongside the downregulation of Bcl-xL by kahweol and the cleavage of Bid, caspase-3, and PARP by cafestol. Kahweol’s effects were also tested on the lung adenocarcinoma cell line A549, demonstrating DNA fragmentation effects and apoptosis, a decrease in STAT3 expression, as well as an increase in caspase-3 cleavage [22].
Finally, non-small cell lung cancer was also tested, specifically, the cell lines NCI-H358 and NCI-H1299 [23]. Apoptosis induced by kahweol was achieved, overall, through a very similar molecular targeting that seems to be common between mesotheliomas, lung adenocarcinomas, and non-small cell lung cancers and that involves an increase in the cleavage of both PARP and caspase-3, eventually leading to apoptosis.
2.2. Oral Squamous Cancer
The oral squamous cancer cell lines HN22 and HSC4 were used to help assess the effects of kahweol in an in vitro environment [24]. Suppression of the transcription factor Sp1, which helps in cell differentiation, cell growth, apoptosis, response to DNA damage, and chromatin remodelling, helped achieve cancer cell apoptosis induced by kahweol.
2.3. Prostate Cancer
The in vitro impact of kahweol acetate and cafestol was assessed in the human prostate cancer cell lines PC-3, DU145, and LNCaP [25]. Kahweol acetate and cafestol significantly inhibited proliferation and migration in addition to enhancing apoptosis in the cell lines. Cleaved caspase-3 and its downstream target cleaved PARP, both pro-apoptotic proteins, were upregulated in all cell lines, while the anti-apoptotic proteins STAT3, Bcl-2, and Bcl-xL were diminished. Androgen receptor (AR), a strong driver of proliferation in prostate cancer, was downregulated following the treatment with kahweol acetate and cafestol. Moreover, the levels of CCL-2 and CCL-5, in addition to those of their receptors CCR-2 and CCR-5, were decreased.
Kahweol acetate and cafestol also exhibited anti-oncogenic activity in vivo. In a xenograft study where human cell lines DU-145 were injected in SCID mouse models, the oral intake of kahweol and cafestol significantly reduced tumour growth.
2.4. Breast Cancer
Kahweol has been shown to exert an anti-tumour effect on breast cancer cell lines. In vitro treatment of MDA-MB231 with kahweol resulted in the inhibition of cell proliferation along with the induction of apoptosis [26]. The levels of the pro-apoptotic proteins caspase-3/7 and 9, as well as of the haemeprotein cytochrome c, were increased by kahweol treatment. Kahweol also led to an increase in H2O2 cytotoxicity in a dose-dependent manner [26].
Kahweol’s effects on the MDA-MB231 cell line can be traced back to the increased levels of phosphorylated Akt (p-Akt) and extracellular-signal-regulated kinase (ERK), activating a signalling pathway that regulates multiple cellular processes such as proliferation and apoptosis [26]. The migratory ability of the cell line was also investigated following kahweol treatment, showing reduced levels of matrix metalloproteinase-9 (MMP-9) and urokinase-type plasminogen activator (uPA).
Kahweol treatment resulted in reduced proliferation and increased apoptosis in the HER2-overexpressing SKBR3 cell line [27]. It also led to the downregulation of HER2, a growth factor essential to proliferation. To evaluate kahweol’s effect on HER2, two molecular targets were assessed: PEA3, which is a suppressor of HER2 transcription, and AP-2, which upregulates HER2. Kahweol treatment increased the levels of PEA3 while reducing those of AP-2, confirming that the two pathways were targeted by kahweol. In addition, fatty acid synthase (FASN), normally elevated in HER2-overexpressing breast cancers, was decreased in concentration, which was attributed to kahweol’s modulation of sterol regulatory element-binding protein-1c (SREBP-1c) activity. Kahweol also led to a reduction in the levels of p-Akt and of its downstream targets mTOR and cyclin D1, which had an additional impact on the downregulation of FASN [27].
2.5. Colorectal Cancer
Colorectal cancer cell lines have also been shown to be susceptible to kahweol. Kahweol induced apoptosis in HCT116 cells in vitro through the overexpression of ATF3, which is mediated by CREB1, ERK1/2, and GSK3β [28]. In addition, kahweol suppressed the proliferation in vitro of HCT116 and SW480 cells, as evidenced by their reduced levels of cyclin D1, a cell cycle protein that promotes progression through the cell cycle [29]. Kahweol-mediated degradation of cyclin D1 was achieved through the phosphorylation of cyclin D1 at threonine-286 (Thr286). This effect was attenuated upon the inhibition of any of kahweol’s molecular targets ERK1/2, JNK, and GSK3β.
Kahweol’s anti-apoptotic characteristics were also observed in the human colon adenocarcinoma HT-29 cells [30,31]. Cell death was induced by kahweol treatment in a dose-dependent manner, which was further proven by the elevation of the pro-apoptotic markers cleaved caspase-3 and PARP, as well as the reduction of the anti-apoptotic markers Bcl-2 and p-AKT. Furthermore, the levels of heat shock proteins (HSP40, HSP70, and HSP90) were diminished following kahweol treatment. HSPs are a family of molecular chaperones that prevent cell death, and their downregulation is associated with increased cytotoxicity of tumour cells.
2.6. Renal Carcinoma
Kahweol’s anti-tumour properties were assessed in several renal carcinoma cell lines in vitro, with a predominant focus on Caki cells. TRAIL, a natural ligand for death receptors found on cancer cells, exerted its apoptotic function on Caki cells more effectively when it was combined with kahweol [33]. Co-treatment with TRAIL and kahweol strongly activated DEVDases and reduced the levels of c-FLIP and Bcl-2, which are anti-apoptotic proteins. The JNK and p38 MAPK pathways were found to mediate these effects. Similarly, the treatment of Caki cells with melatonin and kahweol exhibited comparable findings, which included the induction of apoptosis and the activation of DEVDases [32]. The levels of the pro-apoptotic Bcl-2 protein from the p53 Upregulated Modulator of Apoptosis (PUMA) protein family were also upregulated in response to the treatment. This upregulation was achieved through the C/EBP homologous protein (CHOP), a pro-apoptotic transcription factor. Furthermore, co-treatment with kahweol and sorafenib, a tyrosine kinase inhibitor, had analogous effects on renal carcinoma cells [34]. Caspase-dependent apoptosis and downregulation of c-FLIP and Mcl-1 were induced by those compounds when tested on Caki cells as well as on other renal carcinoma cell lines, including A498 and ACHN cells.
Cafestol was also shown to exhibit its anti-carcinogenic effect in renal Caki cells. Cafestol-induced apoptosis was found to be mediated by the activation of caspase-2 and 3, the upregulation of the pro-apoptotic proteins Bim and Bax, and the downregulation of anti-apoptotic proteins, including c-FLIP, Bcl-2, Mcl-1, and Bcl-xL [31]. The apoptotic effect was also achieved by the inhibition of both STAT3 activation and the PI3K/Akt pathway.
The combined anti-tumour activity of kahweol acetate and cafestol was documented in renal Caki and ACHN cell lines [35]. In these cells, the induction of apoptosis and epithelial–mesenchymal transition upon administration of the diterpenes inhibited proliferation and migration. The inhibition of STAT3 activation and the downregulation of Bcl-2- and Bcl-xL-mediated the apoptosis was exerted by the diterpenes, alongside the upregulation of Bax. Moreover, the diterpenes inhibited Akt and ERK phosphorylation, which are both known to accelerate metastasis and tumour growth.
Finally, cafestol was tested on the Caki cells both in vitro and in vivo using BALB/c-nude mice as the cancer cells recipients [36]. Anti-cancer effects were induced via the synergistic impacts of cafestol and ABT-737, a Bcl-2 family inhibitor. The inactivation of Bcl-2 proteins (Bcl-2, Bcl-xL, and Bcl-w) helped induce apoptosis. PARP cleavage was also increased by the combined actions of cafestol and ABT-737. Mcl-1 protein was also downregulated by this combination in the in vivo setting, leading overall to a pro-apoptotic effect on Caki cells.
2.7. Leukaemia
The U937 leukaemia cell line was tested on in vitro to evaluate kahweol’s apoptotic effect, which was found to be mediated by the activation of caspase-3 and the downregulation of Bcl-2 [38]. The AKT and JNK pathways were also found to be associated with this effect.
Other cell lines such as NB4, K562, HL60, and KG1 were also tested in vitro using cafestol and Ara-c, which is an anti-leukemic agent that was used as a positive control [39]. Increased caspase-3 cleavage was observed in HL60 cells, which helped induce apoptosis via cafestol. Finally, cafestol also helped increase the expression of CD15 and CD11b and decrease the formation of ROS.
Kahweol and cafestol were also shown to enhance the activity of the NK cell line KHYG-1 and its cytolytic effect on the NK-sensitive leukaemia cell line K562. The two diterpenes increased granzyme B expression in NK cells, likely through the phosphorylation of ATF-2, c-Jun, and CREB, transcription factors involved in the regulation of granzyme B. The net result was the enhancement of the cytolytic activity of NK cells in leukaemia cell lines [40].
2.8. Fibrosarcoma
Kahweol acetate was found to attenuate cancer formation, proliferation, and migration by the fibrosarcoma HT-1080 cell line by inhibiting MMP-9, which is usually upregulated by PMA, 12-phorbol 13-myristate acetate, a synthetic compound known for its oncogenic effect [30]. Kahweol acetate was found to act by inhibiting MMP-9, which is usually upregulated by PMA. This inhibition was achieved by the suppression of PMA-induced NF-κB activity, alongside the suppression of the Akt, p38 MAPK, and JNK1/2 signalling pathways, which were also found to activate MMP-9.
2.9. Hepatocellular Carcinoma
In vitro, kahweol exerted apoptotic effects as well as inhibited cell proliferation in the hepatocellular carcinoma cell lines Hep3B, SNU182, and SNU42 [37]. The Src signalling pathway, which is activated upon the phosphorylation of Src, is highly functional in HCC. The Src pathway was blocked by kahweol by inhibiting the expression of p-Akt, p-mTOR, p-p70S6K, and p-4EBP1 in Hep3B and SNU182 cells. This had a direct apoptotic and anti-proliferative effect on the cancer cells. mTOR, which has a proliferative function, was also inhibited by kahweol. Finally, STAT3 was also blocked by kahweol, which induced a pro-apoptotic effect in the HCC cell lines.
2.10. Head and Neck Squamous Cell Carcinoma
Upon the addition of cafestol in vitro, the cell lines of SCC25, CAL27 and FaDu were found to undergo apoptosis in a dose-dependent manner. When combined with cisplatin, cafestol displayed an antagonistic effect in all cell lines; moreover, its addition to radiation therapy showed an additive effect in the cell lines SCC25 and CAL27 [41]. Very limited studies have been performed on head and neck squamous cell carcinoma, which calls for more extensive research on the topic.
3. Conclusions and Future Perspectives
Diterpenes are fat-soluble compounds that can be derived from Coffea arabica beans and are known for their disease-preventing characteristics. Kahweol and cafestol, two diterpenes, were found to have a striking role in preventing the progression of several types of cancer, as discussed above. This effect is achieved mainly by inducing apoptosis, alongside cytotoxicity and mitochondrial damage, which are mediated by targeting certain downstream molecules and pathways, as summarised in Figure 2.
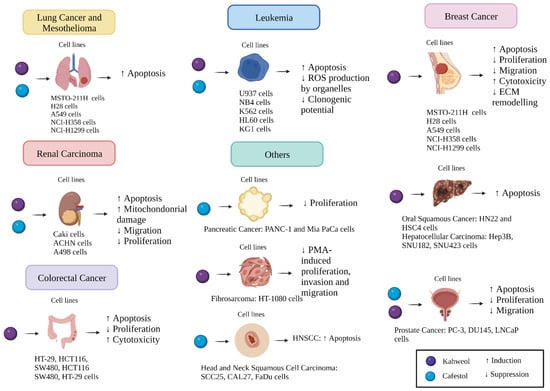
Figure 2.
A diagram summarising the potential anti-cancer mechanisms of kahweol and cafestol in different cancer cell lines. Created by BioRender.com.
Figure 3 portrays the most common molecular targets of kahweol and cafestol, through which they exhibit their tumour-suppressive function. The two diterpenes, as depicted in the figure, either upregulate anti-cancer pathways, such as c-PARP and c-caspase, leading to the apoptosis of cancer cells, or downregulate molecules, including Bcl-2 and Bcl-xL, which also achieves an apoptotic effect.
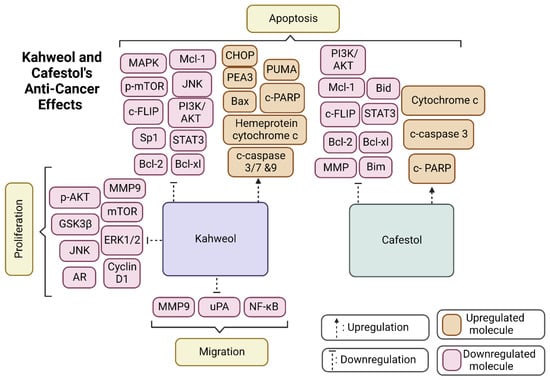
Figure 3.
The diagram summarises kahweol and cafestol’s effects on anticancer pathways, including migration, proliferation, and apoptosis. Created by BioRender.com.
Anticancer properties can also be exerted by other diterpenes and their derivatives. Taxanes, a well-known group of chemotherapeutic agents, are a class of diterpenes extracted from the plant genus Taxus, indicated for the treatment of breast, ovarian, and prostate cancers, among others [42]. Another diterpene, triptolide, has long been held as a promising anticancer agent [43]. It possesses anti-proliferative and pro-apoptotic properties, inducing cell apoptosis through the upregulation of cleaved caspase-3, cleaved caspase-9, cleaved PARP, and Bax and the downregulation of Bcl-2, in a similar way as kahweol and cafestol [44].
The chemotherapeutic role of both diterpenes is mainly exerted in cancer cell lines, with a possible minimal effect on normal body cells. The anticancer effects of these diterpenes are dose-dependent and time-dependent. According to the current literature, the doses of kahweol and cafestol required to exert their anticancer properties are, dependent on the type of cancer and cell line. For example, kahweol suppressed proliferation and promoted apoptosis in lung adenocarcinoma A549 cells at a dose of 10–40 μM when administered for 24 to 48 h [22]. Similarly, an in vitro study reported that 20–80 μM cafestol inhibited human umbilical vein endothelial cells (HUVEC) proliferation in a dose-dependent manner, ultimately achieving cafestol’s anti-angiogenic effects [8]. The precise impact of cafestol and kahweol on normal body cells has not been thoroughly studied and thus warrants further investigation in order to accurately determine the toxicity of these compounds.
Compared to the anticancer doses of kahweol and cafestol, their anti-inflammatory doses are lower. Both diterpenes can inhibit the production of PGE2 and NO, two inflammatory modulators, in lipopolysaccharide (LPS)-activated macrophages in a dose-dependent manner [8]. For instance, at a dose of 0.5–10 μM, kahweol and cafestol can inhibit the activation of IκB kinase (IKK), which is the main activator of the inflammatory transcription factor NF-κB, in LPS-activated macrophages [45]. Furthermore, at the small dose of 0.5 μM, kahweol can potently inhibit cyclooxygenase-2 (COX-2), which is an enzyme that increases the levels of the inflammatory modulators PGE2 and NO [45].
Alongside their potent anti-inflammatory roles, kahweol and cafestol’s anti-oxidant effects are also prominent. The diterpenes exhibit the ability to induce the Keap1/Nrf2/ARE signalling pathway, which is primarily responsible for protection against reactive oxygen species within the liver cells [8]. This activity, like its anti-inflammatory function, is dose-dependent. Lee et al. highlighted the diterpenes’ anti-oxidant ability by using them to treat tetrachloride-induced liver damage in mice [46]. By scavenging free radicals and limiting lipid peroxidation, the diterpenes were able to suppress liver damage, starting from a dose of 20 μM. At higher doses of 100 μM and 200 μM, liver injury was further decreased. Unfortunately, this study was conducted in 2007 [46], and no recent literature highlights any advancements regarding the diterpenes’ anti-oxidant effects, making it an interesting topic to revisit.
Diterpenes have also been utilised in the treatment of other diseases. Vitamin A, a lipid-soluble diterpene, is a dietary supplement that is essential for human health, as it has functions in vision, early growth and development, as well as higher brain functions such as learning and memory [47]. Moreover, Vitamin A is used to produce Tretinoin, an FDA-approved drug that is indicated for the treatment of acne vulgaris, psoriasis, and cutaneous warts [48]. Interestingly, it has also been shown to induce remission in patients with acute promyelocytic leukaemia. Phytol is a diterpene alcohol that, along with its derivatives, displays a multitude of biological functions, such as anxiolytic, antimicrobial, anti-inflammatory, and immune-modulating effects [49]. Vitamin K1 can be extracted from phytol and used in the treatment of clotting factor disorders [50,51].
In conclusion, this review provides sufficient evidence portraying the anticancer role of kahweol and cafestol in vitro and in vivo. Inhibition of proliferation and the induction of apoptosis are the primary mechanisms by which the two diterpenes exert their tumour-suppressive effects. The literature surrounding in vitro studies is impressive, especially in regard to renal cell carcinoma, breast cancer, leukaemia, and colorectal cancer, as the tumour-suppressive effects of kahweol and cafestol were verified in numerous studies. Other cancer cell lines, including mesothelioma, lung adenocarcinoma, non-small cell lung cancer, oral squamous cancer, prostate cancer, hepatocellular carcinoma, fibrosarcoma, and head and neck squamous cell carcinoma, were tested using the two diterpenes in no more than two in vitro studies. Prostate cancer and renal cell carcinoma cells are the only cell lines used in in vivo studies, which confirmed their potential response to kahweol and cafestol, calling for a more extensive clinical investigation. As such, the chemotherapeutic function of these diterpenes should be assessed alongside their pharmacokinetics, including their absorption, bioavailability, metabolism, and elimination, as well as their safety profile via toxicological testing. The two diterpenes have a promising potential to revolutionise the field of cancer treatment.
4. Methods
A systematic literature search was conducted in the databases PubMed and Google Scholar between February 2022 and May 2022. The search terms were “Kahweol and Cancer” and “Cafestol and Cancer”. Each term was searched separately on both databases, and the articles whose title or abstract was found to be relevant were filtered based on the inclusion criteria. The researchers independently selected the articles that discussed the effect of kahweol and cafestol on human cancer cell lines in vivo and in vitro and discarded the articles that did not discuss the anticancer effects of the two diterpenes. Figure 4 summarises the data collection process.
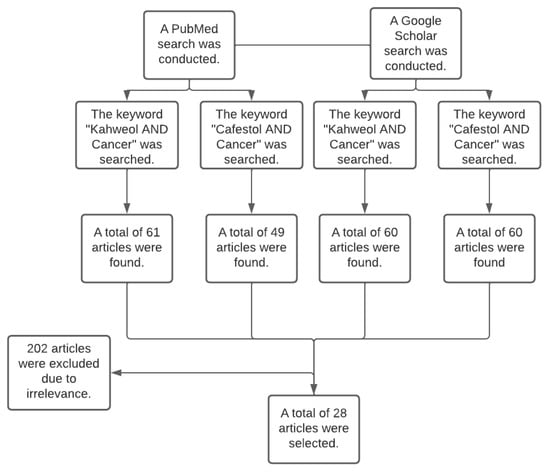
Figure 4.
The flowchart represents the data collection process, including the used databases (PubMed and Google Scholar), the keywords, and the number of included and excluded articles.
Author Contributions
H.B., E.A.-G. and W.E.-H. contributed to conceptualisation and methodology, S.E., R.Q. and R.A.H. contributed to the initial writing of the review. Editing was performed by H.B., Y.B., E.A.-G. and W.E.-H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the College of Research and Graduate Studies, University of Sharjah (Grant number [2001090271] and Grant No. [2001090185].
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AP-2 | Activator Protein 2 |
| AR | Androgen Receptor |
| ATF3 | Cyclic AMP-dependent transcription factor |
| BALB/c-nude mice | Bagg and Albino/c-nude mice |
| Bax | BCL2-Associated X Protein |
| Bcl-2 | B-cell leukemia/lymphoma 2 protein. |
| Bcl-xL | B-cell lymphoma-extra large |
| Bid | BH3 interacting-domain death agonist |
| Bim | Bcl-2-like protein 11 |
| BTF3 | Basic transcription factor 3 |
| CCL2-CCR2 | Chemokine (C-C motif) ligand 2-CC chemokine receptor 2 |
| CCL5-CCR5 | Chemokine (C-C motif) ligand 5-CC chemokine receptor 5 |
| CCR6 | CC chemokine receptor 6 |
| CD11b | Cluster of differentiation 11b |
| CD15 | Cluster of differentiation 15 |
| cFLIP | Cellular FLICE (FADD-like IL-1β-converting enzyme)-inhibitory protein |
| CHOP | C/EBP Homologous Protein |
| DEVDase | Caspase-3-like proteases |
| ECM | Extra-cellular Matrix |
| ER | Endoplasmic Reticulum |
| ERK signaling | Extracellular signal-regulated kinase |
| FASN | Fatty Acid Synthase |
| FDA | Food and Drug administration |
| GSK-3β | Glycogen synthase kinase-3 beta |
| HER2 | Human epidermal growth factor receptor 2 |
| HMVECs | Human microvascular endothelial cell-1 |
| HSP40/70/90 | Human Heat shock protein 40/70/90 |
| ICAM1 | Intercellular adhesion molecule 1 |
| IL-8 | Interleukin-8 |
| JNK | Jun N-terminal kinase |
| LDL | Low-density Lipoprotein |
| MAPK | Mitogen-activated protein kinase |
| mcl1 | Myeloid cell leukemia-1 |
| MCP1 | Monocyte chemoattractant protein-1 |
| MMP-9 | Matrix metallopeptidase 9 |
| mTOR | Mammalian target of rapamycin |
| NK-kB | nNuclear factor kappa light chain enhancer of activated B cells |
| p-4EBP1 | Phosphorylated eukaryotic translation initiation factor |
| p-AKT | Phosphorylated Protein kinase B |
| PARP | Poly-ADP ribose polymerase |
| PD-L1 | Programmed death-ligand 1 |
| PEA3 | Polyoma enhancer activator 3 |
| PI3K/Akt pathway | Phosphatidylinositol 3-kinase/Protein kinase B |
| PMA | Phorbol 12-myristate 13-acetate |
| p-mTOR | Phosphorylated mammalian target of rapamycin |
| p-p70S6K | Phosphorylated Ribosomal protein S6 kinase beta-1 |
| p-Src | Phosphorilated sarcoma |
| p-STAT3 | Phosphorylated signal transducer and activator of transcription 3 |
| PUMA | p53-upregulated modulator of apoptosis |
| ROS | Reactive oxygen species |
| SCID mice | Severe combined immunodeficiency mice |
| Sp1 | Specificity protein 1 |
| SREBP-1c | Sterol regulatory element-binding protein-1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| Thr286 | Threonine 286 |
| TRAIL-Mediated apoptosis | Tumor necrosis factor-related apoptosis-inducing ligand |
| uPA | Urokinase-type plasminogen activator (uPA) |
| XIAP | X-linked inhibitor of apoptosis protein |
References
- Hong, C.T.; Chan, L.; Bai, C.H. The Effect of Caffeine on the Risk and Progression of Parkinson’s Disease: A Meta-Analysis. Nutrients 2020, 12, 1860. [Google Scholar] [CrossRef]
- Gokcen, B.B.; Sanlier, N. Coffee consumption and disease correlations. Crit. Rev. Food Sci. Nutr. 2019, 59, 336–348. [Google Scholar] [CrossRef]
- Feld, J.J.; Lavoie, E.G.; Fausther, M.; Dranoff, J.A. I drink for my liver, Doc: Emerging evidence that coffee prevents cirrhosis. F1000Research 2015, 4, 95. [Google Scholar] [CrossRef]
- Cano-Marquina, A.; Tarin, J.J.; Cano, A. The impact of coffee on health. Maturitas 2013, 75, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Camandola, S.; Plick, N.; Mattson, M.P. Impact of Coffee and Cacao Purine Metabolites on Neuroplasticity and Neurodegenerative Disease. Neurochem. Res. 2019, 44, 214–227. [Google Scholar] [CrossRef] [PubMed]
- El-Huneidi, W.; Anjum, S.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic beta-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules 2021, 26, 5167. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Chae, J.I.; Shim, J.H. Natural diterpenes from coffee, cafestol and kahweol induce apoptosis through regulation of specificity protein 1 expression in human malignant pleural mesothelioma. J. Biomed. Sci. 2012, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef]
- Dias, R.C.; Faria, A.F.D.; Mercadante, A.Z.; Bragagnolo, N.; Benassi, M.D.T. Comparison of extraction methods for kahweol and cafestol analysis in roasted coffee. J. Braz. Chem. Soc. 2013, 24, 492–499. [Google Scholar] [CrossRef]
- Novaes, F.J.M.; Junior, I.I.; Sutili, F.K.; Marriott, P.J.; Bizzo, H.R.; de Aquino Neto, F.R.; de Souza, R.O.M.A.; Rezende, C.M. Lipase-catalysed esters synthesis of cafestol and kahweol. Food Chem. 2018, 259, 226–233. [Google Scholar] [CrossRef] [PubMed]
- De Roos, B.; Meyboom, S.; Kosmeijer-Schuil, T.G.; Katan, M.B. Absorption and urinary excretion of the coffee diterpenes cafestol and kahweol in healthy ileostomy volunteers. J. Intern. Med. 1998, 244, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Cruchten, S.T.; de Waart, D.R.; Kunne, C.; Hooiveld, G.J.; Boekschoten, M.V.; Katan, M.B.; Elferink, R.P.; Witkamp, R.F. Absorption, distribution, and biliary excretion of cafestol, a potent cholesterol-elevating compound in unfiltered coffees, in mice. Drug Metab. Dispos. 2010, 38, 635–640. [Google Scholar] [CrossRef]
- De Roos, B.; Van Tol, A.; Urgert, R.; Scheek, L.M.; Van Gent, T.; Buytenhek, R.; Princen, H.M.; Katan, M.B. Consumption of French-press coffee raises cholesteryl ester transfer protein activity levels before LDL cholesterol in normolipidaemic subjects. J. Intern. Med. 2000, 248, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, B.; Ranheim, T.; Nenseter, M.S.; Huggett, A.C.; Drevon, C.A. Effect of a coffee lipid (cafestol) on cholesterol metabolism in human skin fibroblasts. J. Lipid Res. 1998, 39, 901–912. [Google Scholar] [CrossRef]
- Post, S.M.; de Wit, E.C.; Princen, H.M. Cafestol, the cholesterol-raising factor in boiled coffee, suppresses bile acid synthesis by downregulation of cholesterol 7 alpha-hydroxylase and sterol 27-hydroxylase in rat hepatocytes. Arter. Thromb. Vasc. Biol. 1997, 17, 3064–3070. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Hong, H.J.; Liu, J.C.; Chen, J.J. Cafestol Activates Nuclear Factor Erythroid-2 Related Factor 2 and Inhibits Urotensin II-Induced Cardiomyocyte Hypertrophy. Am. J. Chin. Med. 2019, 47, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Chen, P.Y.; Cheng, T.H.; Chao, H.H.; Liu, J.C.; Chen, J.J. Corrigendum to “Cafestol Inhibits Cyclic-Strain-Induced Interleukin-8, Intercellular Adhesion Molecule-1, and Monocyte Chemoattractant Protein-1 Production in Vascular Endothelial Cells”. Oxid. Med. Cell. Longev. 2019, 2019, 1536397. [Google Scholar] [CrossRef] [PubMed]
- Fumimoto, R.; Sakai, E.; Yamaguchi, Y.; Sakamoto, H.; Fukuma, Y.; Nishishita, K.; Okamoto, K.; Tsukuba, T. The coffee diterpene kahweol prevents osteoclastogenesis via impairment of NFATc1 expression and blocking of Erk phosphorylation. J. Pharmacol. Sci. 2012, 118, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Fukuma, Y.; Sakai, E.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Cafestol has a weaker inhibitory effect on osteoclastogenesis than kahweol and promotes osteoblast differentiation. Biofactors 2015, 41, 222–231. [Google Scholar] [CrossRef]
- Choi, M.J.; Park, E.J.; Oh, J.H.; Min, K.J.; Yang, E.S.; Kim, Y.H.; Lee, T.J.; Kim, S.H.; Choi, Y.H.; Park, J.W.; et al. Cafestol, a coffee-specific diterpene, induces apoptosis in renal carcinoma Caki cells through down-regulation of anti-apoptotic proteins and Akt phosphorylation. Chem. Biol. Interact. 2011, 190, 102–108. [Google Scholar] [CrossRef]
- Moeenfard, M.; Cortez, A.; Machado, V.; Costa, R.; Luis, C.; Coelho, P.; Soares, R.; Alves, A.; Borges, N.; Santos, A. Anti-Angiogenic Properties of Cafestol and Kahweol Palmitate Diterpene Esters. J. Cell. Biochem. 2016, 117, 2748–2756. [Google Scholar] [CrossRef]
- Kim, H.G.; Hwang, Y.P.; Jeong, H.G. Kahweol blocks STAT3 phosphorylation and induces apoptosis in human lung adenocarcinoma A549 cells. Toxicol. Lett. 2009, 187, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Bang, W.; Cho, J.H.; Lee, R.H.; Kim, S.H.; Kim, M.S.; Park, S.M.; Shin, J.C.; Chung, H.J.; Oh, K.B.; et al. Kahweol induces apoptosis by suppressing BTF3 expression through the ERK signaling pathway in non-small cell lung cancer cells. Int. J. Oncol. 2016, 49, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.I.; Jeon, Y.J.; Shim, J.H. Anti-proliferative properties of kahweol in oral squamous cancer through the regulation specificity protein 1. Phytother. Res. 2014, 28, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, H.; Izumi, K.; Natsagdorj, A.; Naito, R.; Makino, T.; Kadomoto, S.; Hiratsuka, K.; Shigehara, K.; Kadono, Y.; Narimoto, K.; et al. Coffee diterpenes kahweol acetate and cafestol synergistically inhibit the proliferation and migration of prostate cancer cells. Prostate 2019, 79, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.; Quesada, A.R.; Medina, M.A. Insights on the antitumor effects of kahweol on human breast cancer: Decreased survival and increased production of reactive oxygen species and cytotoxicity. Biochem. Biophys. Res. Commun. 2014, 447, 452–458. [Google Scholar] [CrossRef]
- Oh, S.H.; Hwang, Y.P.; Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Chung, Y.H.; Chung, Y.C.; Jeong, H.G. Kahweol inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. Food Chem. Toxicol. 2018, 121, 326–335. [Google Scholar] [CrossRef]
- Park, G.H.; Song, H.M.; Jeong, J.B. Kahweol from Coffee Induces Apoptosis by Upregulating Activating Transcription Factor 3 in Human Colorectal Cancer Cells. Biomol. Ther. 2017, 25, 337–343. [Google Scholar] [CrossRef]
- Park, G.H.; Song, H.M.; Jeong, J.B. The coffee diterpene kahweol suppresses the cell proliferation by inducing cyclin D1 proteasomal degradation via ERK1/2, JNK and GKS3beta-dependent threonine-286 phosphorylation in human colorectal cancer cells. Food Chem. Toxicol. 2016, 95, 142–148. [Google Scholar] [CrossRef]
- Choi, J.H.; Hwang, Y.P.; Jin, S.W.; Lee, G.H.; Kim, H.G.; Han, E.H.; Kim, S.K.; Kang, K.W.; Chung, Y.C.; Jeong, H.G. Suppression of PMA-induced human fibrosarcoma HT-1080 invasion and metastasis by kahweol via inhibiting Akt/JNK1/2/p38 MAPK signal pathway and NF-kappaB dependent transcriptional activities. Food Chem. Toxicol. 2019, 125, 1–9. [Google Scholar] [CrossRef]
- Choi, D.W.; Lim, M.S.; Lee, J.W.; Chun, W.; Lee, S.H.; Nam, Y.H.; Park, J.M.; Choi, D.H.; Kang, C.D.; Lee, S.J.; et al. The Cytotoxicity of Kahweol in HT-29 Human Colorectal Cancer Cells Is Mediated by Apoptosis and Suppression of Heat Shock Protein 70 Expression. Biomol. Ther. 2015, 23, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Um, H.J.; Oh, J.H.; Kim, Y.N.; Choi, Y.H.; Kim, S.H.; Park, J.W.; Kwon, T.K. The coffee diterpene kahweol sensitizes TRAIL-induced apoptosis in renal carcinoma Caki cells through down-regulation of Bcl-2 and c-FLIP. Chem. Biol. Interact. 2010, 186, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Um, H.J.; Park, J.W.; Kwon, T.K. Melatonin sensitizes Caki renal cancer cells to kahweol-induced apoptosis through CHOP-mediated up-regulation of PUMA. J. Pineal. Res. 2011, 50, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Um, H.J.; Kim, J.I.; Kwon, T.K. The coffee diterpene kahweol enhances sensitivity to sorafenib in human renal carcinoma Caki cells through down-regulation of Mcl-1 and c-FLIP expression. Oncotarget 2017, 8, 83195–83206. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Izumi, K.; Hiratsuka, K.; Kano, H.; Shimada, T.; Nakano, T.; Kadomoto, S.; Naito, R.; Iwamoto, H.; Yaegashi, H.; et al. Anti-proliferative and anti-migratory properties of coffee diterpenes kahweol acetate and cafestol in human renal cancer cells. Sci. Rep. 2021, 11, 675. [Google Scholar] [CrossRef]
- Woo, S.M.; Min, K.J.; Seo, B.R.; Nam, J.O.; Choi, K.S.; Yoo, Y.H.; Kwon, T.K. Cafestol overcomes ABT-737 resistance in Mcl-1-overexpressed renal carcinoma Caki cells through downregulation of Mcl-1 expression and upregulation of Bim expression. Cell Death Dis. 2014, 5, e1514. [Google Scholar] [CrossRef]
- Seo, H.Y.; Lee, S.H.; Lee, J.H.; Lee, J.H.; Jang, B.K.; Kim, M.K. Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 10509. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Lee, J.T.; Yang, E.S.; Chang, J.S.; Lee, D.S.; Kim, S.H.; Choi, Y.H.; Park, J.W.; Kwon, T.K. The coffee diterpene kahweol induces apoptosis in human leukemia U937 cells through down-regulation of Akt phosphorylation and activation of JNK. Apoptosis 2009, 14, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.S.; Spindola, D.G.; Bechara, A.; Garcia, D.M.; Palmeira-Dos-Santos, C.; Peixoto-da-Silva, J.; Erustes, A.G.; Michelin, L.F.G.; Pereira, G.J.S.; Smaili, S.S.; et al. Cafestol, a diterpene molecule found in coffee, induces leukemia cell death. Biomed. Pharmacother. 2017, 92, 1045–1054. [Google Scholar] [CrossRef]
- Saito, T.; Abe, D.; Nogata, Y.J.F.S. Coffee diterpenes potentiate the cytolytic activity of KHYG-1 NK Leukemia cells. Food Sci. Technol. Res. 2015, 21, 281–284. [Google Scholar] [CrossRef]
- Kotowski, U.; Heiduschka, G.; Seemann, R.; Eckl-Dorna, J.; Schmid, R.; Kranebitter, V.; Stanisz, I.; Brunner, M.; Lill, C.; Thurnher, D. Effect of the coffee ingredient cafestol on head and neck squamous cell carcinoma cell lines. Strahlenther. Onkol. 2015, 191, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Noel, P.; Von Hoff, D.D.; Saluja, A.K.; Velagapudi, M.; Borazanci, E.; Han, H. Triptolide and Its Derivatives as Cancer Therapies. Trends Pharmacol. Sci. 2019, 40, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.P.; Tan, Z.J.; Wu, X.S.; Liu, T.Y.; Jiang, L.; Bao, R.F.; Shu, Y.J.; Li, M.L.; Weng, H.; Ding, Q.; et al. Triptolide induces s phase arrest and apoptosis in gallbladder cancer cells. Molecules 2014, 19, 2612–2628. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jung, K.S.; Jeong, H.G. Suppressive effects of the kahweol and cafestol on cyclooxygenase-2 expression in macrophages. FEBS Lett. 2004, 569, 321–326. [Google Scholar] [CrossRef]
- Lee, K.J.; Choi, J.H.; Jeong, H.G. Hepatoprotective and antioxidant effects of the coffee diterpenes kahweol and cafestol on carbon tetrachloride-induced liver damage in mice. Food Chem. Toxicol. 2007, 45, 2118–2125. [Google Scholar] [CrossRef]
- Parker, G.L.; Smith, L.K.; Baxendale, I.R. Development of the industrial synthesis of vitamin A. Tetrahedron 2016, 72, 1645–1652. [Google Scholar] [CrossRef]
- Yoham, A.L.; Casadesus, D. Tretinoin. In StatPearls [Internet]; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra Shill, M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K. Vitamins K1 and K2: The Emerging Group of Vitamins Required for Human Health. J. Nutr. Metab. 2017, 2017, 6254836. [Google Scholar] [CrossRef]
- Daines, A.M.; Payne, R.J.; Humphries, M.E.; Abell, A.D. The synthesis of naturally occurring vitamin K and vitamin K analogues. Curr. Org. Chem. 2003, 7, 1625–1634. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).