
Anticancer, Cardio-Protective and Anti-Inflammatory Potential of Natural-Sources-Derived Phenolic Acids

 , ,
, ,  ,
,
Abstract
1. Introduction
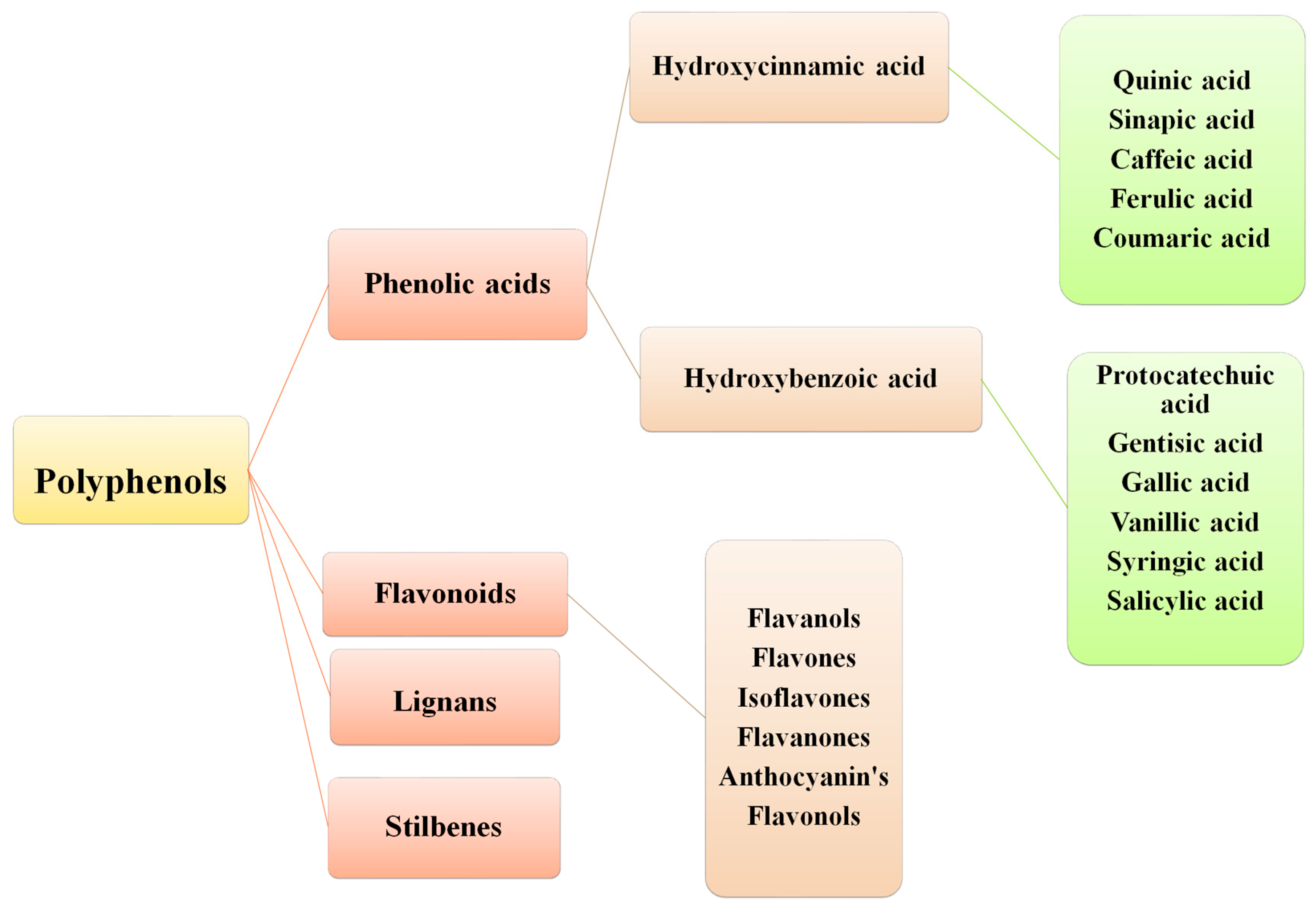
2. Sources and Classification of PAs
3. Hydroxybenzoic Acid Derivatives and Their Sources
4. Derivatives of Hydroxycinnamic Acids and Their Sources
5. Biosynthesis of PAs
6. Structure Activity Relationship of PAs
7. Pharmacological Activities of PAs
7.1. Anti-Cancer Actions of PAs
7.1.1. Anticancer Activity of EA
7.1.2. Anticancer Activity of FA and PCA
7.1.3. Anticancer Activity of CA and Derivatives
7.2. Anti-Inflammatory and Analgesic Actions of PAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acids | Cell Line/Animal Model | Dose | Mechanism | Type of Cancer | References |
|---|---|---|---|---|---|
| Caffeic acid, coumaric acid, ferulic acid | HT-29-D4 A549 | 50,100, 150 and 200 µM for 1 day | (-) cell proliferation (-) ROS production (-) cell adhesion | Lung carcinoma and colon adenocarcinoma | [53] |
| Ferulic acid, p-coumaric acid | Caco-2 | 15, 150 and 1500 µM for 3 days | ↓ cell growth ↓ cell proportion in G1 phase | Colon cancer | [52] |
| Caffeic acid | HCT15 | 500, 1000 and 2000 µM | (-) cell viability (-) colony development ↑cell accumulation at G1 ↑ scavenging activity √ apoptosis | Colon cancer | [59] |
| 3,4-Dihydroxyphenylacetic acid | HCT116 | 600 mg for >1.5 days | ↓ proliferative activity | Prostate and colon cancer | [60] |
| Cycloartenyl ferulate | SW480 SW620 Colo-201 | 40, 80 and 160 mM for 3 days | √ apoptosis in metastatic cancer cells √ tumor regression (-) cell growth | Colorectal adenocarcinoma | [61] |
| Male nude mice | 1.6 and 32 mg/kg for 10 days | ||||
| Dicaffeoylquinic acid | DLD-1 | 10, 100, 500 and 1000 µM for > 3 days | (-) cell growth and mutation √ apoptosis | Colon cancer | [42,62] |
| CAPE and CAPPE | BALB/C mice | 50 nmol/kg/day for 42 days | ↓ colorectal tumor (-) PCNA, FASN and MMP ↑Cell cycle arrest (-) cell growth by controlling P13K/AkT and mTOR cascades | Colon cancer | [55] |
| HCT-116SW-480 | 0, 5, 10, 20, 50 and 100 µM IC50s in CRC HCT-116 for CAPE 44.2 mM and CAPPE 32.7 mM In SW-480 | ||||
| Syringic acid | SW-480 | (2000 μM) | (-) cell proliferation ↑apoptosis Down-regulated NF-κB Modulate oxidative stress | Colorectal cancer | [63] |
| Rats | 50 mg/kg for 210 days | ||||
| Ellagic acid | MCF-7 cells | 10−7 to 10−9 M | ↑Estrogenic activity of ER-α, β | Breast cancer | [64] |
| Ellagic acid | SiHa, C33A and HeLa cells | 30 μM | (-) cell propagation Arrest G1 √Apoptosis (-) phosphorylation of JAK2 signaling and STAT3 expression | Cervical carcinoma | [65] |
| Ellagic acid | ES-2 PA-1 MRC-5 | 10–100 μM | G1 arrest ↑ P53 and p21 ↓ cyclin D1 and E. | Ovarian cancer | [51] |
| Ellagic acid | AIH (PC-3) and PLS10 | 80–200 μM | (-) invasive potential through action on the activity of proteases, such as collagenase/gelatinase and collagenase IV | Prostate cancer | [66] |
| 3-O-Methylgallic acid and gallic acid | Caco-2 | 10–100 mM | ↓ cell viability, S-phase √ apoptosis (-) G0/G1 phase, NF-κβ, AP-1, STAT-1, OCT-1 | Colon cancer | [50] |
| Gallic acid | 22Rv1 and DU145, PWR-1E | 0–100 µM | (-) tumor cell proliferation (√) apoptosis ↓ micro vessel density in tumor | Prostate cancer | [67] |
| Athymic male nude mice | CG or 0.3 and 1% (w/v) dose of GA for 5 days/week | ||||
| Vanillic acid | male Balb nude mice | 30 mg/kg | (-) HIF-1α by suppression of mTOR/p70S6K/4E-BP1 Raf kinase/MEK pathways and angiogenesis | Prostate cancer | [68] |
| HCT116, Hep3B A549 HUVEC cells | 0–30 µM | ||||
| Sinapic acid | PC-3 LNCaP | 250–4000 µM | (-) cell proliferation ↓ caspase-3 activity (-) cell invasion ↑ expression of BAX ↓ MMP-9 in PC-3 cell expression In LNCaP cells, ↑ BAX, CASP3, CASP7 and CYCS ↓ CDH2, MMP-2 and MMP-9 cells expression | Prostate cancer | [69] |
| Phenolic Acid/Source | Animal Model/Cell Line | Dose | Mechanism of Action | Result | References |
|---|---|---|---|---|---|
| Brazilian green propolis extract (EPP-AF) | LPS-induced macrophages in Swiss mice | 30, 100 and 300 μg/mL for 18 h | ↓ IL-1β, casp-1↓ (-) inflammosome activation | Regulates inflammasome path and prevents inflammatory activity | [70] |
| Shikimic acid | Dopamine inducing mechanical hyperalgesia in mice | 100 and 200 mg/kg for 30–180 min | ↓ cell viability ↓ TNF-α, nitrite production, IL-1β | Prevents inflammation and mild to moderate pain | [71] |
| Ferulic acid | Middle cerebral artery occlusion (MCAo) in male rats | (100 mg/kg i.v.) for 1 day | (-) ICAM-1 ↓ NF-κB | Reduces cerebral infarction and possess anti-inflammatory effect | [56] |
| Caffeic acid Butyl and octyl esters | Carrageenan-induced paw edema in mice | 30 mg/kg for 5 days | ↓ IL-1β levels ↓ MPO activity ↑ iNOS | Anti-inflammatory action | [72] |
| p-coumaric acid | CFA induces arthritis in rats | 100 mg/kg | (-) NF-κB ↓ macrophage phagocytic index ↑the serum immunoglobulin | Immunosuppressant and anti-inflammatory agent in arthritis | [73] |
| Sinapic acid | TPA and AA induced ear edema in mice | 0, 819, 1024, 1280, 1600, 2000, 2500 mg/kg for two weeks | C inhibit the MPO ↓ TNF-α ↓ IL-6 | Inhibits acute and chronic inflammation | [58] |
| Shikimic acid | Macrophage (RAW 264.7) cell DMEM | 10 mM | (-) cell viability and nitrite accumulation ↓ TNF-α, IL-1β | Inhibited LPS-induced cellular pro-inflammatory cytokines | [71] |
| Caffeic acid derivatives | LPS stimulated RAW 264.7, DMEM | 0.5 mg/ml | ↓ Nitrite accumulation (-) iNOS expression | In vitro anti-inflammatory action | [72] |
| Protocatechuic acid | RAW 264.7 cells, RPMI 1640 | 1, 2, 5, and 25 μM | ↓ TNF-α, IL-1β, NO and PGE2 | ↓ TNF-α and IL-1β ↓ NO and PGE2 (-) iNOS and COX-2 deprivation (-) phosphorylated NF-κβ, p38, ERK, and JNK | [57] |
| Protocatechuic acid | MAEC, RPMI 1640 | 0.05, 0.5, 5.0, 10, 20, and 40 μmol/L | (-) adhesion of HL-60 cells to MAEC’s (-) VCAM-1 and ICAM-1 mRNA expression↓ NF-κβ initiation ↓ TNF-α-induced cellular damage and monocyte adhesion | Anti-inflammatory | [74] |
7.3. Cardio-Protective Actions of PAs
7.3.1. Atherosclerosis
7.3.2. Myocardial Infarction and Stroke
7.3.3. Hypertension
| Phenolic Acids | Animal Model and Dose | Method | Biomarkers | Result | References |
|---|---|---|---|---|---|
| Gentisic acid | 100 mg/kg/day for 3 weeks in mice | Cardiac hypertrophy and cardiac fibrosis induced by transverse aortic restriction | Down-regulation of Sp1 and ERK1 or 2 pathways | Anti-hypertrophic and anti-fibrotic effects | [83] |
| Gentisic acid | 5, 10, 20 and 40 mg/kg/day for 28 days in Balb mice | Cardiac damage with doxorubicin | ↓ cardiac myofibrillar necrosis, ↓ hyalinization necrosis ↓ cardiac toxicity | Prevented cardiotoxic effects of doxorubicin and treated cardiac damage | [84] |
| Caffeic acid phenyl ester (CAPE) | CAPE 1 h before (3 and 15 mg/kg) or 30 min after (15 mg/kg) the onset of ischemia in rabbits | Acute myocardial ischemia reperfusion injury | (-) p38 MPAK phosphorylation and caspase activation ↓ LDH, CK, CK-MB induced by I/R injury, ↓ IL-1β and TNF-α | Cardioprotective effects against I/R injury | [89] |
| Ferulic acid and ascorbic acid | FA = 20 mg/kg and AsA = 80 mg/kg for 6 days in rats | Isoproterenol-induced myocardial infarction in rats | Improved CAT, SOD↓ CPK and LDH levels | Synergistically reduced myocardial infarction and showed cardioprotective effects | [85] |
| Ferulic acid | 20 mg and 40 mg/kg in Wistar rats | Doxorubicin-induced myocardial toxicity | (-) expression of ANP and BNP ↑myocardial GSH and Na+/K+ ATPase | Reduced cardiotoxicity | [90] |
| Syringic acid | H9c2 cardiomyocytes | hypoxia/reoxygenation injury | (-) apoptosis of cardiomyocytes ↓ p38 MAPK and JNK activation pathways | Inhibited apoptosis of cardiomyocytes, and prevented myocardial infarction | [91] |
| Syringic acid, revasterol | 50 mg/kg for 30 days in rats | Isoproterenol-induced cardiotoxicity | ↓ cardiac biomarkers, ↓ antioxidant enzymes ↓ docking with NF-κB and inflammatory markers | Cardioprotective | [92] |
| p-coumaric acid | 8 mg/kg for 7 days in male rats | MI induced by isoproterenol | (-)cardiac hypertrophy and variations in lipoproteins (-)HMG-CoA reductase | Treated myocardial infarction and normalized the ECG | [93] |
| Protocatechuic acid (PCA) | 250–500 mg/kg in male rats | MI/R injury | ↓ Myocardial infarction, ↓ serum TNF-α ↓ platelets count | Up-regulation of phosphorylated Akt expression in cardiomyocytes and cardioprotective effects in rats | [94] |
| Neonatal rat cardiomyocytes | Hypoxia, reoxygenation induced in cardiomyocytes | (-) apoptotic rate (-) cleaved caspase- | |||
| Protocatechuic acid (PCA) | 50 and 100 mg/kg for 84 days in male rats | T1DM induced by a streptozotocin | √cardiac function √ANS balance, (-) cardiac mitochondrial damage ↑ anti-apoptotic protein | Cardioprotective in type 1 diabetic rats | [95] |
8. Toxicity of Phenolic Acids
9. Conclusions and Future Aspects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From Chemistry to Biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Goleniowski, M.; Bonfill, M.; Cusido, R. Phenolic Acids 63, in Phenolic Acids. In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Amawi, H.; Ashby, J.C.R.; Samuel, T.; Peraman, R.; Tiwari, A.K. Polyphenolic Nutrients in Cancer Chemoprevention and Metastasis: Role of the Epithelial-to-Mesenchymal (EMT) Pathway. Nutrients 2017, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic potential of plant phenolic acids in the treatment of cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- Van Nies, J.A.; De Jong, Z.; van der Helm-van Mil, A.H.; Knevel, R.; Le Cessie, S.; Huizinga, T.W. Improved treatment strategies reduce the increased mortality risk in early RA patients. Rheumatology 2010, 49, 2210–2216. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef] [PubMed]
- Macheix, J.-J.; Fleuriet, A.; Billot, J. Fruit Phenolics; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Sikorska, M.; Matławska, I.; Głowniak, K.; Zgórka, G. Qualitative and quantitative analysis of phenolic acids in Asclepias syriaca L. Acta Pol. Pharm. 2000, 57, 69–72. [Google Scholar]
- Vuolo, M.M.; Lima, V.S.; Junior, M.R.M. Phenolic compounds: Structure, classification, and antioxidant power. In Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–50. [Google Scholar]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.; Hellström, J.; Törrönen, R. Phenolic Acids in Berries, Fruits, and Beverages. J. Agric. Food Chem. 2006, 54, 7193–7199. [Google Scholar] [CrossRef] [PubMed]
- Mohdaly, A.A.A.; Sarhan, M.A.; Mahmoud, A.; Ramadan, M.F.; Smetanska, I. Antioxidant efficacy of potato peels and sugar beet pulp extracts in vegetable oils protection. Food Chem. 2010, 123, 1019–1026. [Google Scholar] [CrossRef]
- Dos Santos Ferreira, C.I.; Pereyra, A.; Patriarca, A.R.; Mazzobre, M.F.; Polak, T.; Abram, V.; Buera, M.d.P.; Poklar Ulrih, N. Phenolic compounds in extracts from Eucalyptus globulus leaves and Calendula officinalis flowers. J. Nat. Prod. Resour. 2016, 2, 53–57. [Google Scholar]
- El Gharras, H. Polyphenols: Food sources, properties and applications—A review. Int. J. Food Sci. Technol. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Alsulami, A.L.; Gull, M. Screening of antimicrobial potential and bioactive components of selected medicinal plants against infectious bacterial isolates from leukemia patients. J. Exp. Biol. Agric. Sci. 2018, 6, 836–849. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef]
- Soto-Hernández, M.; García-Mateos, R.; Palma-Tenango, M. Plant Physiological Aspects of Phenolic Compounds; BoD—Books on Demand: Nordstedt, Germany, 2019. [Google Scholar]
- Marchiosi, R.; Dos Santos, W.D.; Constantin, R.P.; De Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.D.P.; Abrahão, J. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic Acids in Foods: An Overview of Analytical Methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, R.; Prakash, D.; Sarma, B.; Singh, H. Antioxidant and anti-quorum sensing activities of green pod of Acacia nilotica L. Food Chem. Toxicol. 2009, 47, 778–786. [Google Scholar] [CrossRef]
- Subhan, N.; Burrows, G.E.; Kerr, P.G.; Obied, H.K. Phytochemistry, ethnomedicine, and pharmacology of Acacia. Stud. Nat. Prod. Chem. 2018, 57, 247–326. [Google Scholar]
- Landete, J.M. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, functions and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Heinonen, M. Berry Phenolics and Their Antioxidant Activity. J. Agric. Food Chem. 2001, 49, 4076–4082. [Google Scholar] [CrossRef] [PubMed]
- Muthukumaran, S.; Tranchant, C.; Shi, J.; Ye, X.; Xue, S.J. Ellagic acid in strawberry (Fragaria spp.): Biological, technological, stability, and human health aspects. Food Qual. Saf. 2017, 1, 227–252. [Google Scholar] [CrossRef]
- Pinto, M.D.S.; Lajolo, F.M.; Genovese, M.I. Bioactive compounds and quantification of total ellagic acid in strawberries (Fragaria x ananassa Duch.). Food Chem. 2008, 107, 1629–1635. [Google Scholar] [CrossRef]
- Vazirian, M.; Khanavi, M.; Amanzadeh, Y.; Hajimehdipoor, H. Quantification of Gallic Acidin Fruits of Three Medicinal Plants. Iran. J. Pharm. Res. IJPR 2011, 10, 233–236. [Google Scholar]
- Mirunalini, S.; Krishnaveni, M. Therapeutic potential of Phyllanthus emblica (amla): The ayurvedic wonder. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 93–105. [Google Scholar] [CrossRef]
- Khan, A.K.; Rashid, R.; Fatima, N.; Mahmood, S.; Mir, S.; Khan, S.; Jabeen, N.; Murtaza, G. Pharmacological activities of protocatechuic acid. Acta Pol. Pharm. 2015, 72, 643–650. [Google Scholar]
- Kim, J.K.; Park, S.U. A recent overview on the biological and pharmacological activities of ferulic acid. EXCLI J. 2019, 18, 132–138. [Google Scholar]
- Manjunatha, B.K. Antibacterial activity of Pterocarpus santalinus. Indian J. Pharm. Sci. 2006, 68, 115. [Google Scholar] [CrossRef]
- Manuja, R.; Sachdeva, S.; Jain, A.; Chaudhary, J. A comprehensive review on biological activities of p-hydroxy benzoic acid and its derivatives. Int. J. Pharm. Sci. Rev. Res. 2013, 22, 109–115. [Google Scholar]
- Chong, K.; Zuraini, Z.; Sasidharan, S.; Devi, P.K.; Latha, L.Y.; Ramanathan, S. Antimicrobial activity of Elaeis guineensis leaf. Pharmacologyonline 2008, 3, 379–386. [Google Scholar]
- Heinonen, M.I. Carotenoids and provitamin A activity of carrot (Daucus carota L.) cultivars. J. Agric. Food Chem. 1990, 38, 609–612. [Google Scholar] [CrossRef]
- Abedi, F.; Razavi, B.M.; Hosseinzadeh, H. A review on gentisic acid as a plant derived phenolic acid and metabolite of aspirin: Comprehensive pharmacology, toxicology, and some pharmaceutical aspects. Phytother. Res. 2019, 34, 729–741. [Google Scholar] [CrossRef]
- Holobiuc, I.; Blîndu, R. In vitro culture of the protected rare species Gentiana lutea L. for conservative purpose. Contrib. Bot. 2008, 42, 125–134. [Google Scholar]
- Momčilović, I.; Grubiršić, D.; Nešković, M. Transgenic Gentiana species (Gentian). In Transgenic Crops III; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 123–138. [Google Scholar] [CrossRef]
- Luís, Â.; Neiva, D.; Pereira, H.; Gominho, J.; Domingues, F.; Duarte, A.P. Stumps of Eucalyptus globulus as a Source of Antioxidant and Antimicrobial Polyphenols. Molecules 2014, 19, 16428–16446. [Google Scholar] [CrossRef] [PubMed]
- Rababah, T.M.; Ereifej, K.I.; Esoh, R.B.; Al-u'datt, M.H.; Alrababah, M.A.; Yang, W. Antioxidant activities, total phenolics and HPLC analyses of the phenolic compounds of extracts from common Mediterranean plants. Nat. Prod. Res. 2011, 25, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Kurata, R.; Adachi, M.; Yamakawa, O.; Yoshimoto, M. Growth Suppression of Human Cancer Cells by Polyphenolics from Sweetpotato (Ipomoea batatas L.) Leaves. J. Agric. Food Chem. 2006, 55, 185–190. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hashemi, F.; Moghadam, E.R.; Hashemi, F.; Entezari, M.; Najafi, M. Curcumin in cancer therapy: A novel adjunct for combination chemotherapy with paclitaxel and alleviation of its adverse effects. Life Sci. 2020, 256, 117984. [Google Scholar] [CrossRef]
- Vogt, P.K.; Kang, S.; Elsliger, M.-A.; Gymnopoulos, M. Cancer-specific mutations in phosphatidylinositol 3-kinase. Trends Biochem. Sci. 2007, 32, 342–349. [Google Scholar] [CrossRef]
- Halilovic, E.; She, Q.-B.; Ye, Q.; Pagliarini, R.; Sellers, W.R.; Solit, D.B.; Rosen, N. PIK3CA Mutation Uncouples Tumor Growth and Cyclin D1 Regulation from MEK/ERK and Mutant KRAS Signaling. Cancer Res. 2010, 70, 6804–6814. [Google Scholar] [CrossRef]
- Rosa, L.d.S.; Silva, N.J.A.; Soares, N.C.P.; Monteiro, M.C.; Teodoro, A.J. Anticancer properties of phenolic acids in colon cancer—A review. J. Nutr. Food Sci. 2016, 6, 1000468. [Google Scholar]
- Chander, M. Anticancer efficacy of some plant Phenolics—A recent scenario. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1746–1768. [Google Scholar] [CrossRef]
- Choi, J.; Jiang, X.; Jeong, J.B.; Lee, S.-H. Anticancer Activity of Protocatechualdehyde in Human Breast Cancer Cells. J. Med. Food 2014, 17, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, B.A.; Geoffroy, O.; Willingham, M.C.; Re, G.G.; Nixon, D.W. p53/p21 (WAF1/CIP1) expression and its possible role in G1 arrest and apoptosis in ellagic acid treated cancer cells. Cancer Lett. 1999, 136, 215–221. [Google Scholar] [CrossRef]
- Forester, S.C.; Choy, Y.Y.; Waterhouse, A.L.; Oteiza, P.I. The anthocyanin metabolites gallic acid, 3-O-methylgallic acid, and 2,4,6-trihydroxybenzaldehyde decrease human colon cancer cell viability by regulating pro-oncogenic signals. Mol. Carcinog. 2012, 53, 432–439. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Lu, L.-C.; Tsai, M.-H.; Chen, Y.-J.; Chen, Y.-Y.; Yao, S.-P.; Hsu, C.-P. The Inhibitory Effect of Ellagic Acid on Cell Growth of Ovarian Carcinoma Cells. Evid.-Based Complement. Altern. Med. 2013, 2013, 306705. [Google Scholar] [CrossRef]
- Janicke, B.; Önning, G.; Oredsson, S.M. Differential Effects of Ferulic Acid and p-Coumaric Acid on S Phase Distribution and Length of S Phase in the Human Colonic Cell Line Caco-2. J. Agric. Food Chem. 2005, 53, 6658–6665. [Google Scholar] [CrossRef]
- Bouzaiene, N.N.; Jaziri, S.K.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef]
- Lin, H.P.; Jiang, S.S.; Chuu, C.P. Caffeic acid phenethyl ester causes p21Cip1 induction, Akt signaling reduction, and growth inhibition in PC-3 human prostate cancer cells. PLoS ONE 2012, 7, e31286. [Google Scholar] [CrossRef]
- Chiang, E.; Tsai, S.-Y.; Kuo, Y.-H.; Pai, M.-H.; Chiu, H.-L.; Rodriguez, R.L.; Tang, F.-Y. Caffeic Acid Derivatives Inhibit the Growth of Colon Cancer: Involvement of the PI3-K/Akt and AMPK Signaling Pathways. PLoS ONE 2014, 9, e99631. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Ho, T.-Y.; Lee, E.-J.; Su, S.-Y.; Tang, N.-Y.; Hsieh, C.-L. Ferulic Acid Reduces Cerebral Infarct Through Its Antioxidative and Anti-Inflammatory Effects Following Transient Focal Cerebral Ischemia in Rats. Am. J. Chin. Med. 2008, 36, 1105–1119. [Google Scholar] [CrossRef]
- Min, S.-W.; Ryu, S.-N.; Kim, D.-H. Anti-inflammatory effects of black rice, cyanidin-3-O-β-d-glycoside, and its metabolites, cyanidin and protocatechuic acid. Int. Immunopharmacol. 2010, 10, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.-F.; Hu, Z.; Ip, S.-P.; Chen, J.-N.; Su, Z.-R.; Lai, X.-P.; Lin, Z.-X. Comparison of the anti-inflammatory effects of Sinapis alba and Brassica juncea in mouse models of inflammation. Phytomedicine 2018, 50, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K. Growth Inhibition by Caffeic Acid, One of the Phenolic Constituents of Honey, in HCT 15 Colon Cancer Cells. Sci. World J. 2012, 2012, 372345. [Google Scholar] [CrossRef]
- Gao, K.; Xu, A.; Krul, C.; Venema, K.; Liu, Y.; Niu, Y.; Lu, J.; Bensoussan, L.; Seeram, N.P.; Heber, D. Of the Major Phenolic Acids Formed during Human Microbial Fermentation of Tea, Citrus, and Soy Flavonoid Supplements, Only 3,4-Dihydroxyphenylacetic Acid Has Antiproliferative Activity. J. Nutr. 2006, 136, 52–57. [Google Scholar] [CrossRef]
- Kong, C.K.; Lam, W.; Chiu, L.C.; Ooi, V.E.; Sun, S.S.; Wong, Y.-S. A rice bran polyphenol, cycloartenyl ferulate, elicits apoptosis in human colorectal adenocarcinoma SW480 and sensitizes metastatic SW620 cells to TRAIL-induced apoptosis. Biochem. Pharmacol. 2009, 77, 1487–1496. [Google Scholar] [CrossRef]
- Lin, L.Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 120, 319–326. [Google Scholar] [CrossRef]
- Mihanfar, A.; Darband, S.G.; Sadighparvar, S.; Kaviani, M.; Mirza-Aghazadeh-Attari, M.; Yousefi, B.; Majidinia, M. In vitro and in vivo anticancer effects of syringic acid on colorectal cancer: Possible mechanistic view. Chem. Interact. 2021, 337, 109337. [Google Scholar] [CrossRef] [PubMed]
- Papoutsi, Z.; Kassi, E.; Tsiapara, A.; Fokialakis, N.; Chrousos, G.P.; Moutsatsou, P. Evaluation of Estrogenic/Antiestrogenic Activity of Ellagic Acid via the Estrogen Receptor Subtypes ERα and ERβ. J. Agric. Food Chem. 2005, 53, 7715–7720. [Google Scholar] [CrossRef] [PubMed]
- Kalezic, A.; Macanovic, B.; Garalejic, E.; Korac, A.; Otasevic, V.; Korac, B. Level of NO/nitrite and 3-nitrotyrosine in seminal plasma of infertile men: Correlation with sperm number, motility and morphology. Chem. Interact. 2018, 291, 264–270. [Google Scholar] [CrossRef]
- Pitchakarn, P.; Chewonarin, T.; Ogawa, K.; Suzuki, S.; Asamoto, M.; Takahashi, S.; Shirai, T.; Limtrakul, P. Ellagic Acid Inhibits Migration and Invasion by Prostate Cancer Cell Lines. Asian Pac. J. Cancer Prev. 2013, 14, 2859–2863. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Velmurugan, B.; Rajamanickam, S.; Agarwal, R.; Agarwal, C. Gallic Acid, an Active Constituent of Grape Seed Extract, Exhibits Anti-proliferative, Pro-apoptotic and Anti-tumorigenic Effects Against Prostate Carcinoma Xenograft Growth in Nude Mice. Pharm. Res. 2009, 26, 2133–2140. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhou, S.; Yang, S. Vanillic Acid Suppresses HIF-1α Expression via Inhibition of mTOR/p70S6K/4E-BP1 and Raf/MEK/ERK Pathways in Human Colon Cancer HCT116 Cells. Int. J. Mol. Sci. 2019, 20, 465. [Google Scholar] [CrossRef]
- Eroğlu, C.; Avcı, E.; Vural, H.; Kurar, E. Anticancer mechanism of Sinapic acid in PC-3 and LNCaP human prostate cancer cell lines. Gene 2018, 671, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hori, J.I.; Zamboni, D.S.; Carrão, D.B.; Goldman, G.H.; Berretta, A.A. The Inhibition of Inflammasome by Brazilian Propolis (EPP-AF). Evid.-Based Complement. Altern. Med. 2013, 2013, 418508. [Google Scholar] [CrossRef]
- Rabelo, T.K.; Guimarães, A.G.; Oliveira, M.A.; Gasparotto, J.; Serafini, M.R.; de Souza Araújo, A.A.; Quintans-Júnior, L.J.; Moreira, J.C.F.; Gelain, D.P. Shikimic acid inhibits LPS-induced cellular pro-inflammatory cytokines and attenuates mechanical hyperalgesia in mice. Int. Immunopharmacol. 2016, 39, 97–105. [Google Scholar] [CrossRef]
- Da Cunha, F.M.; Duma, D.; Assreuy, J.; Buzzi, F.C.; Niero, R.; Campos, M.M.; Calixto, J.B. Caffeic Acid Derivatives: In Vitro and In Vivo Anti-inflammatory Properties. Free Radic. Res. 2004, 38, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Pragasam, S.J.; Venkatesan, V.; Rasool, M. Immunomodulatory and Anti-inflammatory Effect of p-Coumaric Acid, a Common Dietary Polyphenol on Experimental Inflammation in Rats. Inflammation 2012, 36, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wei, X.; Yan, X.; Jin, T.; Ling, W. Protocatechuic Acid, a Metabolite of Anthocyanins, Inhibits Monocyte Adhesion and Reduces Atherosclerosis in Apolipoprotein E-Deficient Mice. J. Agric. Food Chem. 2010, 58, 12722–12728. [Google Scholar] [CrossRef]
- Leifert, W.R.; Abeywardena, M.Y. Cardioprotective actions of grape polyphenols. Nutr. Res. 2008, 28, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Faoro, F. Bioactivity of Grape Chemicals for Human Health. Nat. Prod. Commun. 2009, 4, 611–634. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Volkova, N.; Coleman, R.; Aviram, M. Grape Powder Polyphenols Attenuate Atherosclerosis Development in Apolipoprotein E Deficient (E0) Mice and Reduce Macrophage Atherogenicity. J. Nutr. 2005, 135, 722–728. [Google Scholar] [CrossRef]
- Frederiksen, H.; Mortensen, A.; Schrøder, M.; Frandsen, H.; Bysted, A.; Knuthsen, P.; Rasmussen, S.E. Effects of red grape skin and seed extract supplementation on atherosclerosis in Watanabe heritable hyperlipidemic rabbits. Mol. Nutr. Food Res. 2007, 51, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Saibabu, V.; Fatima, Z.; Khan, L.A.; Hameed, S. Therapeutic Potential of Dietary Phenolic Acids. Adv. Pharmacol. Sci. 2015, 2015, 823539. [Google Scholar] [CrossRef] [PubMed]
- Mouhamed, D.; Ezzaher, A.; Gaha, L.; Douki, W.; Najjar, M.F. In vitro effects of salicylic acid on plasma paraoxonase 1 activity. J. Drug Metab. Toxicol. 2013, 4, 1–3. [Google Scholar] [CrossRef]
- Zhou, L.; Yu, X.; Meng, Q.; Li, H.; Niu, C.; Jiang, Y.; Cai, Y.; Li, M.; Li, Q.; An, C. Resistin reduces mitochondria and induces hepatic steatosis in mice by the protein kinase C/protein kinase G/p65/PPAR gamma coactivator 1 alpha pathway. Hepatology 2013, 57, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Kee, H.J.; Jin, L.; Ryu, Y.; Choi, S.Y.; Kim, G.R.; Jeong, M.H. Gentisic acid attenuates pressure overload-induced cardiac hypertrophy and fibrosis in mice through inhibition of the ERK 1/2 pathway. J. Cell. Mol. Med. 2018, 22, 5964–5977. [Google Scholar] [CrossRef] [PubMed]
- Altinoz, M.A.; Elmaci, I.; Cengiz, S.; Emekli-Alturfan, E.; Ozpinar, A. From epidemiology to treatment: Aspirin’s prevention of brain and breast-cancer and cardioprotection may associate with its metabolite gentisic acid. Chem.-Biol. Interact. 2018, 291, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Yogeeta, S.K.; Gnanapragasam, A.; Kumar, S.S.; Subhashini, R.; Sathivel, A.; Devaki, T. Synergistic interactions of Ferulic acid with Ascorbic acid: Its cardioprotective role during isoproterenol induced myocardial infarction in rats. Mol. Cell. Biochem. 2006, 283, 139–146. [Google Scholar] [CrossRef]
- Sarr, M.; Chataigneau, M.; Martins, S.; Schott, C.; El Bedoui, J.; Oak, M.-H.; Muller, B.; Chataigneau, T.; Schini-Kerth, V.B. Red wine polyphenols prevent angiotensin II-induced hypertension and endothelial dysfunction in rats: Role of NADPH oxidase. Cardiovasc. Res. 2006, 71, 794–802. [Google Scholar] [CrossRef]
- Lecour, S.; Lamont, K.T. Natural polyphenols and cardioprotection. Mini Rev. Med. Chem. 2011, 11, 1191–1199. [Google Scholar]
- Opie, L.H.; Commerford, P.J.; Gersh, B.J.; Pfeffer, M.A. Controversies in ventricular remodelling. Lancet 2006, 367, 356–367. [Google Scholar] [CrossRef]
- Tan, J.; Ma, Z.; Han, L.; Du, R.; Zhao, L.; Wei, X.; Hou, D.; Johnstone, B.H.; Farlow, M.R.; Du, Y. Caffeic acid phenethyl ester possesses potent cardioprotective effects in a rabbit model of acute myocardial ischemia-reperfusion injury. Am. J. Physiol. Circ. Physiol. 2005, 289, H2265–H2271. [Google Scholar] [CrossRef]
- Aswar, U.; Mahajan, U.; Kandhare, A.; Aswar, M. Ferulic acid ameliorates doxorubicin-induced cardiac toxicity in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 659–668. [Google Scholar] [CrossRef]
- Ding, S.-K.; Wang, L.-X.; Guo, L.-S.; Luo, P.; Du, J.-J.; Zhao, Z.-L.; Wang, G.-G. Syringic acid inhibits apoptosis pathways via downregulation of p38MAPK and JNK signaling pathways in H9c2 cardiomyocytes following hypoxia/reoxygenation injury. Mol. Med. Rep. 2017, 16, 2290–2294. [Google Scholar] [CrossRef]
- Shaik, A.H.; Al Omar, S.Y.; Mohammad, A.; Kodidhela, L.D. Combined cardio-protective ability of syringic acid and resveratrol against isoproterenol induced cardio-toxicity in rats via attenuating NF-kB and TNF-α pathways. Sci. Rep. 2020, 10, 3426. [Google Scholar]
- Roy, A.J.; Prince, P.S.M. Preventive effects of p-coumaric acid on cardiac hypertrophy and alterations in electrocardiogram, lipids, and lipoproteins in experimentally induced myocardial infarcted rats. Food Chem. Toxicol. 2013, 60, 348–354. [Google Scholar] [CrossRef]
- Tang, X.-L.; Liu, J.-X.; Dong, W.; Li, P.; Li, L.; Lin, C.-R.; Zheng, Y.-Q.; Cong, W.-H.; Hou, J.-C. The Cardioprotective Effect of Protocatechuic Acid on Myocardial Ischemia/Reperfusion Injury. J. Pharmacol. Sci. 2014, 125, 176–183. [Google Scholar] [CrossRef]
- Semaming, Y.; Kumfu, S.; Pannangpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Protocatechuic acid exerts a cardioprotective effect in type 1 diabetic rats. J. Endocrinol. 2014, 223, 13–23. [Google Scholar] [CrossRef]










| Phenolic Acid Class | Compound | Rich Plant/Fruit Source | Family | Part Used | Therapeutic Uses | Reference |
|---|---|---|---|---|---|---|
| Di-gallic acid | Acacia nilotica (L.) P.J.H. Hurter and Mabb. | Fabaceae | Pods | Anti-inflammatory, antioxidant | [23,24] | |
| Ellagic acid | Fragaria x ananassa (Duchesne ex Westson) Duchesne ex Rozier | Rosaceae | Fruits | Anti-cancer, chronic pancreatitis, fibrosis | [8,25,26,27,28] | |
| Gallic acid | Cynomorium coccineum L. Phyllanthus embelica L. | Euphorbiaceae | Fruits | Anti-oxidant, anti-inflammatory, antiulcer, anti-diabetic | [21,29,30] | |
| Protocatechuic acid | Prunus domestica L. Vitis vinifera L. | Rosaceae Vitaceae | Fruit | Anti-oxidant, anti-inflammatory, antibacterial | [31] | |
| Hydroxy cinnamic acids | Ferulic acid | Avena sativa L. Oryaz sativa L. | Gramineae | Seed | Anti-oxidant, anti-inflammatory, anti-viral, anti-microbial, anti-thrombotic, anti-carcinogenic | [32] |
| p-HBA | Elaeis guineensis Jacq. Pterocarpus santalinus L. | Palmae Fabaceae | Dried leaves, stem, and bark | Antimicrobial, anti-mutagenic, anti-estrogenic, hypoglycemic, anti-inflammatory, nematicidal, antiviral | [33,34,35,36] | |
| Gentisic acid | Gentiana acaulis L. | Gentianaceae | Roots | Hepatoprotective, anti-cancer, anti-inflammatory, anti-oxidant, cardio-protective | [37,38,39] | |
| Caffeic acid | Eucalyptus globulus Labill. | Mitraceae | Wood, bark | Antioxidant, antimicrobial, anti-diabetic | [40,41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afnan; Saleem, A.; Akhtar, M.F.; Sharif, A.; Akhtar, B.; Siddique, R.; Ashraf, G.M.; Alghamdi, B.S.; Alharthy, S.A. Anticancer, Cardio-Protective and Anti-Inflammatory Potential of Natural-Sources-Derived Phenolic Acids. Molecules 2022, 27, 7286. https://doi.org/10.3390/molecules27217286
Afnan, Saleem A, Akhtar MF, Sharif A, Akhtar B, Siddique R, Ashraf GM, Alghamdi BS, Alharthy SA. Anticancer, Cardio-Protective and Anti-Inflammatory Potential of Natural-Sources-Derived Phenolic Acids. Molecules. 2022; 27(21):7286. https://doi.org/10.3390/molecules27217286
Chicago/Turabian StyleAfnan, Ammara Saleem, Muhammad Furqan Akhtar, Ali Sharif, Bushra Akhtar, Rida Siddique, Ghulam Md Ashraf, Badrah S. Alghamdi, and Saif A. Alharthy. 2022. "Anticancer, Cardio-Protective and Anti-Inflammatory Potential of Natural-Sources-Derived Phenolic Acids" Molecules 27, no. 21: 7286. https://doi.org/10.3390/molecules27217286
APA StyleAfnan, Saleem, A., Akhtar, M. F., Sharif, A., Akhtar, B., Siddique, R., Ashraf, G. M., Alghamdi, B. S., & Alharthy, S. A. (2022). Anticancer, Cardio-Protective and Anti-Inflammatory Potential of Natural-Sources-Derived Phenolic Acids. Molecules, 27(21), 7286. https://doi.org/10.3390/molecules27217286