
Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF-κB

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
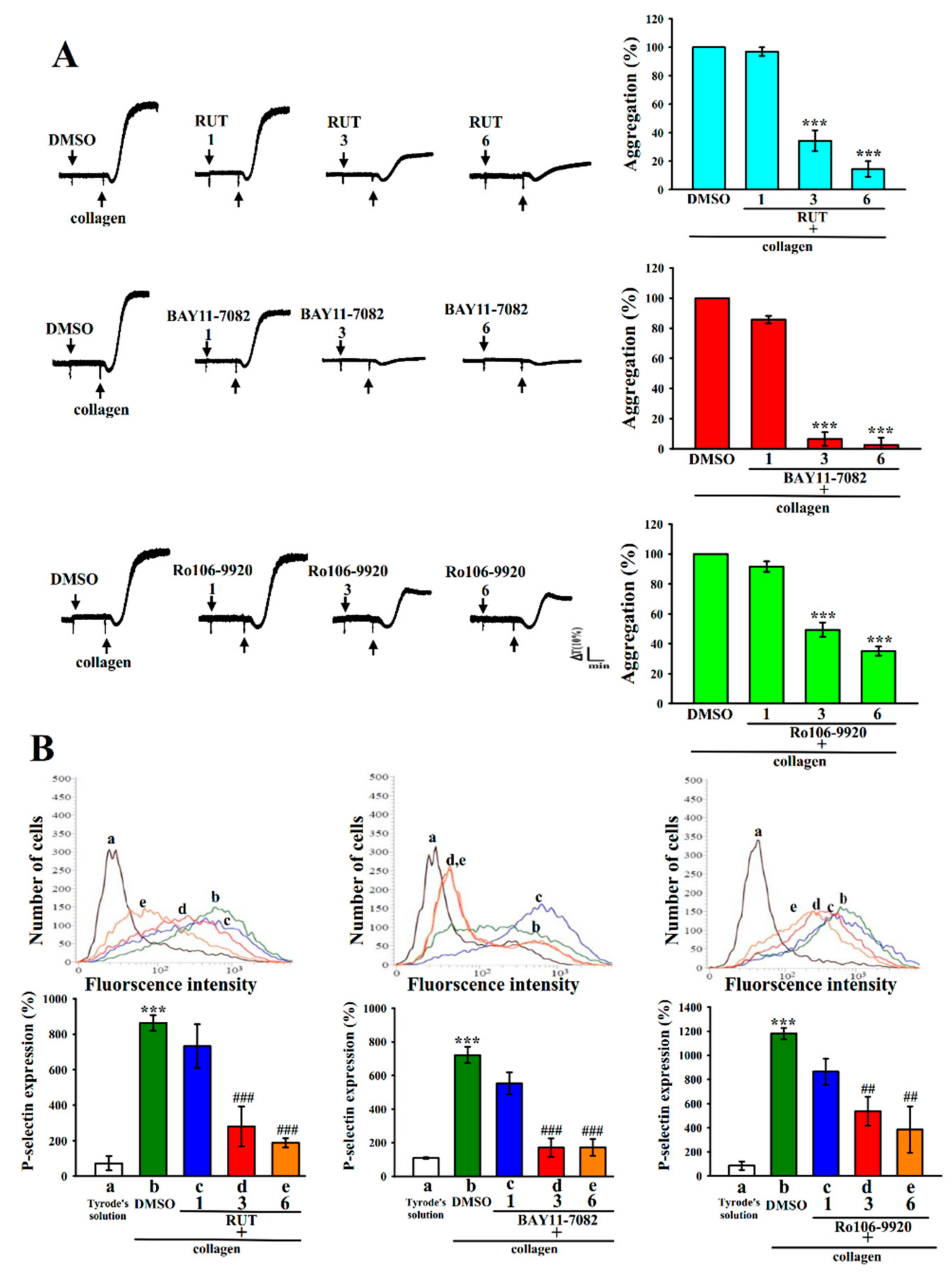
2.1. Relative Activities of RUT with NF-κB Inhibitors in Human Platelet Aggregation and P-Selectin Expression
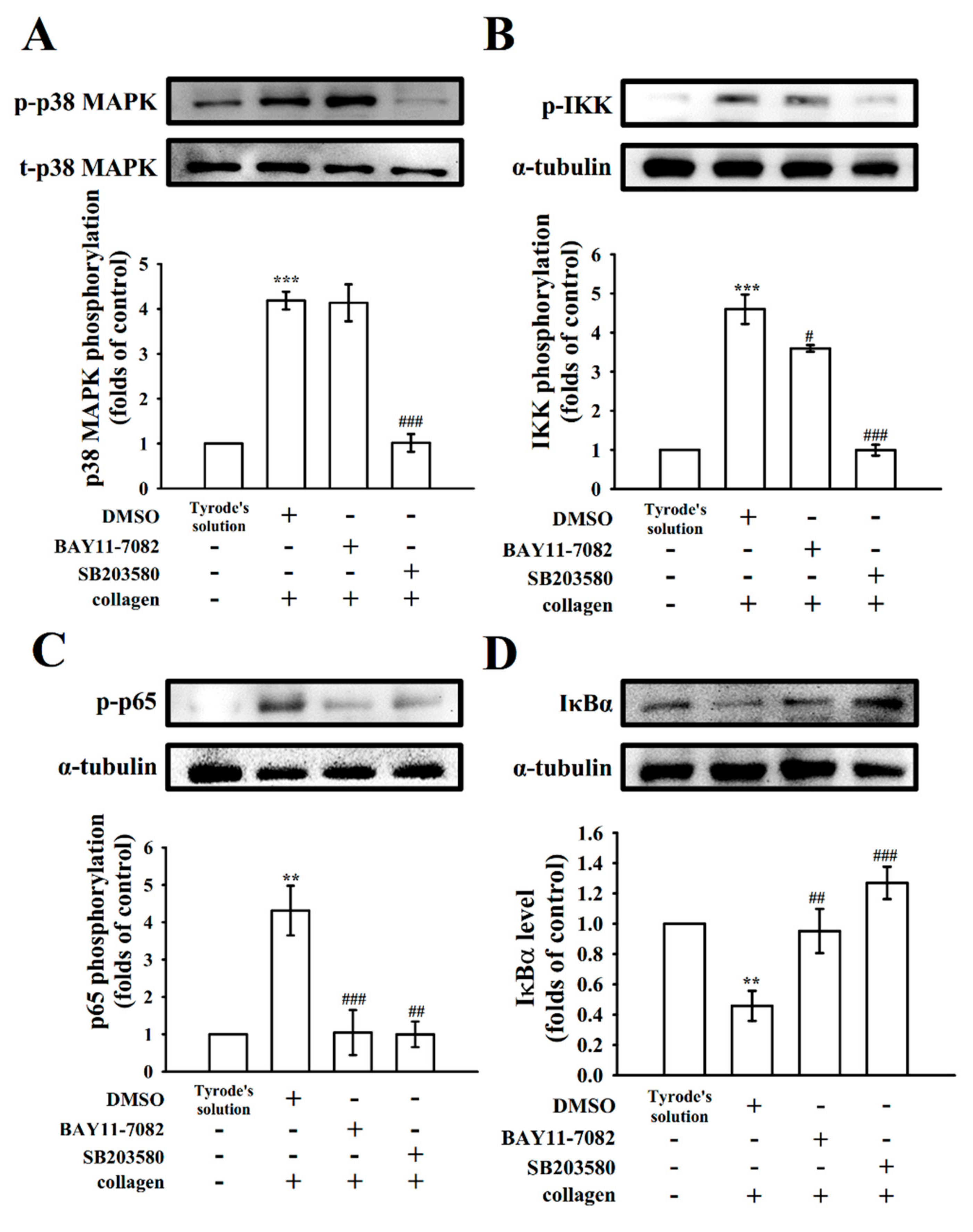
2.2. Regulatory Characteristics of RUT in NF-κB Signals
2.3. Verification of the Signal Connection between p38 and NF-κB
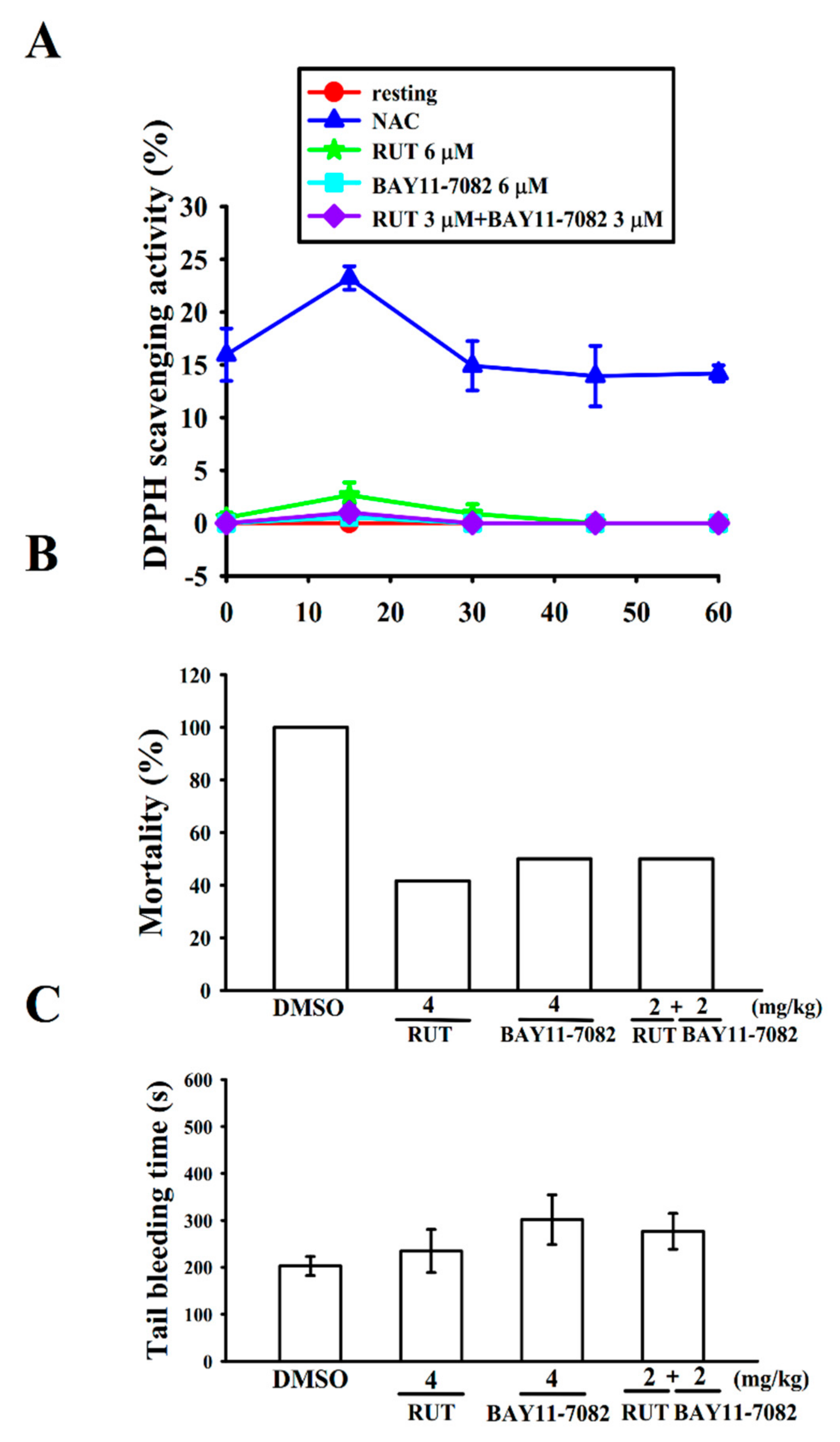
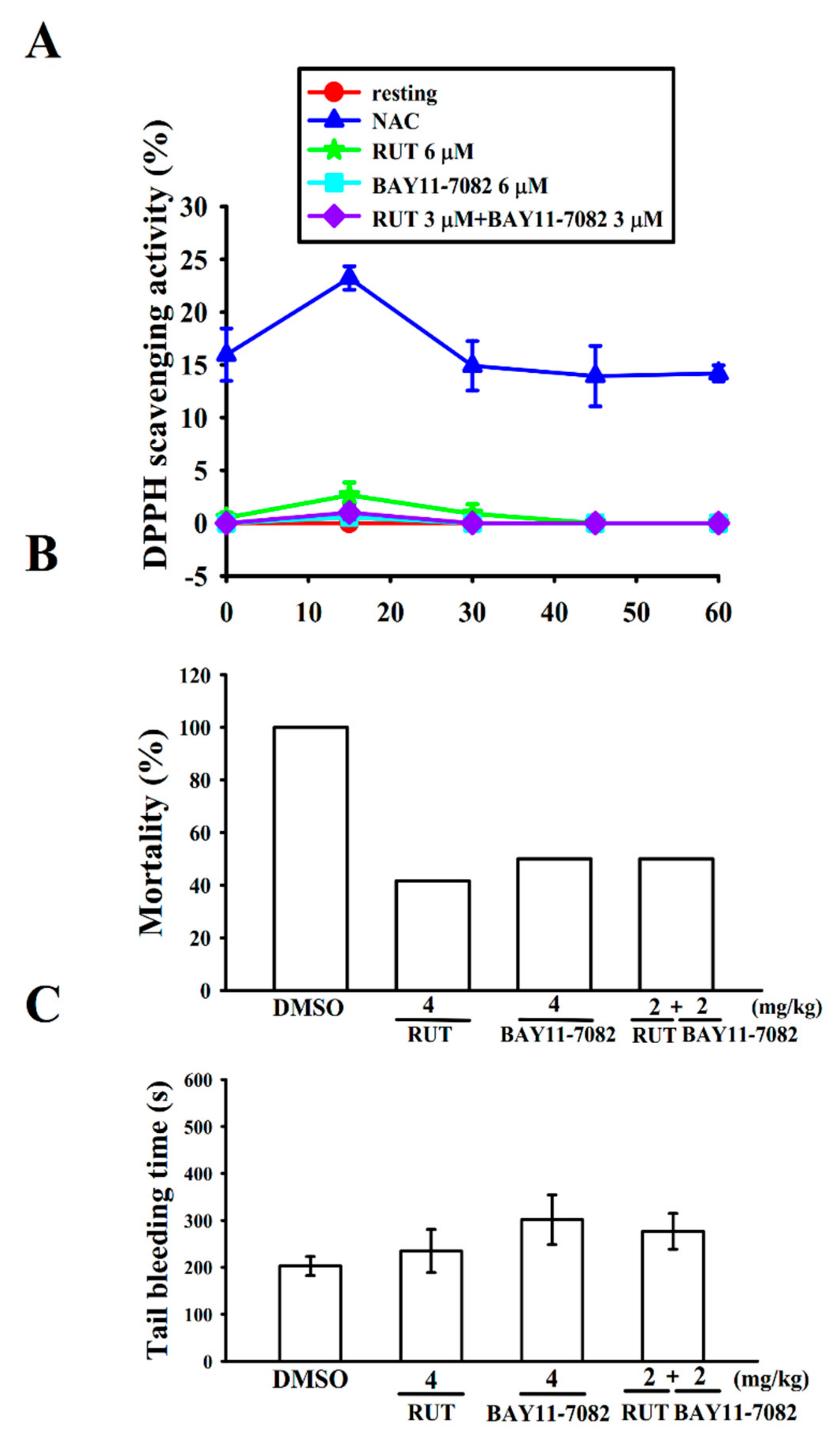
2.4. Interaction with Stable Free Radical-Scavenging Action of RUT and BAY11-7082 in a Cell-Free System
2.5. Relative Potency of RUT and BAY11-7082 in Acute Pulmonary Thrombosis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Platelet Aggregation and Surface P-Selectin Expression
4.3. Immunoblotting
4.4. Confocal Laser Fluorescence Microscopy
4.5. Measurement of Stable Free Radical Scavenging Activity
4.6. ADP-Induced Pulmonary Thrombosis in Mice
4.7. Tail Bleeding Time
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sheu, J.R. Pharmacological effects of rutaecarpine, an alkaloid isolated from evodia eutaecarpa. Cardiovasc. Drug Rev. 1999, 17, 237–245. [Google Scholar]
- Qin, X.P.; Zeng, S.Y.; Li, D.; Chen, Q.Q.; Luo, D.; Zhang, Z.; Hu, G.Y.; Deng, H.W.; Li, Y.J. Calcitonin gene-related peptide-mediated depressor effect and inhibiting vascular hypertrophy of rutaecarpine in renovascular hypertensive rats. J. Cardiovasc. Pharmacol. 2007, 50, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Shum, A.Y.; Liao, J.F.; Chen, C.F. Studies of the cellular mechanisms underlying the vasorelaxant effects of rutaecarpine, a bioactive component extracted from an herbal drug. J. Cardiovasc. Pharmacol. 1997, 29, 490–498. [Google Scholar] [CrossRef]
- Wu, X.B.; Luo, X.Q.; Gu, S.Y.; Xu, J.H. The effects of polygonum cuspidatum extract on wound healing in rats. J. Ethnopharmacol. 2012, 141, 934–937. [Google Scholar] [CrossRef]
- Yamahara, J.; Yamada, T.; Kitani, T.; Naitoh, Y.; Fujimura, H. Antianoxic action and active constituents of Evodiae Fructus. Chem. Pharma. Bull. 1989, 37, 1820–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawaz, M.; Langer, H.; May, A.E. May platelets in inflammation and atherogenesis. J. Clin. Investig. 2005, 115, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Sheu, J.R.; Yen, M.H.; Hung, W.C.; Lee, Y.M.; Su, C.H.; Huang, T.F. Triflavin inhibits platelet-induced vasoconstriction in de-endothelialized aorta. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 3461–3468. [Google Scholar] [CrossRef]
- Tedgui, A.; Mallat, Z. Cytokines in atherosclerosis: Pathogenic and regulatory pathways. Physiol. Rev. 2006, 86, 515–581. [Google Scholar] [CrossRef] [Green Version]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Monaco, C.; Andreakos, E.; Kiriakidis, S.; Mauri, C.; Bicknell, C.; Foxwell, B.; Cheshire, N.; Paleolog, E.; Feldmann, M. Canonical pathway of nuclear factor kappa B activation selectively regulates proinflammatory and prothrombotic responses in human atherosclerosis. Proc. Natl. Acad. Sci. USA 2004, 101, 5634–5639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu, J.R.; Kan, Y.C.; Hung, W.C.; Su, C.H.; Lin, C.H.; Lee, Y.M.; Yen, M.H. The antiplatelet activity of rutaecarpine, an alkaloid isolated from Evodia rutaecarpa, is mediated through inhibition of phospholipase C. Thromb. Res. 1998, 92, 53–64. [Google Scholar] [CrossRef]
- Hsia, C.W.; Huang, W.C.; Yang, C.H.; Hsia, C.H.; Jayakumar, T.; Bhavan, P.S.; Sheu, J.R.; Chiou, K.R. Comparison of the potency of pterostilbene with NF-κB inhibitors in platelet activation: Mutual activation by Akt-NF-κB signaling in human platelets. Appl. Sci. 2021, 11, 6149. [Google Scholar] [CrossRef]
- Sheu, J.R.; Hung, W.C.; Wu, C.H.; Lee, Y.M.; Yen, M.H. Antithrombotic effect of rutaecarpine, an alkaloid isolated from Evodia rutaecarpa, on platelet plug formation in in vivo experiments. Br. J. Haematol. 2000, 110, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Cosemans, J.M.; Iserbyt, B.F.; Deckmyn, H.; Heemskerk, J.W. Multiple ways to switch platelet integrins on and off. J. Thromb. Haemost. 2008, 6, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Hung, W.C.; Lee, Y.M.; Yen, M.H. Mechanism of inhibition of platelet aggregation by rutaecarpine, an alkaloid isolated from Evodia rutaecarpa. Eur. J. Pharmacol. 1996, 318, 469–475. [Google Scholar] [CrossRef]
- Huang, C.J.; Huang, W.C.; Lin, W.T.; Shu, L.H.; Sheu, J.R.; Tran, O.T.; Hsia, C.W.; Jayakumar, T.; Bhavan, P.S.; Hsieh, C.Y.; et al. Rutaecarpine, an alkaloid from Evodia rutaecarpa, can prevent platelet activation in humans and reduce microvascular thrombosis in mice: Crucial role of the PI3K/Akt/GSK3β signal axis through a cyclic nucleotides/VASP—independent mechanism. Int. J. Mol. Sci. 2021, 22, 11109. [Google Scholar] [CrossRef] [PubMed]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, C.; Shi, P.; Gao, W.; Gu, J.; Geng, Y.; Yang, W.; Wu, N.; Wang, Y.; Xu, Y.; et al. Platelet MEKK3 regulates arterial thrombosis and myocardial infarct expansion in mice. Blood Adv. 2018, 2, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.E.; Renshaw, M.W.; Pfaff, M.; Forsyth, J.; Keivens, V.M.; Schwartz, M.A.; Ginsberg, M.H. Suppression of integrin activation: A novel function of a Ras/Raf-initiated MAP kinase pathway. Cell 1997, 88, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Adam, F.; Kauskot, A.; Rosa, J.P.; Bryckaert, M. Mitogen-activated protein kinases in hemostasis and thrombosis. J. Thromb. Haemost. 2008, 6, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Stoclet, J.C.; Chataigneau, T.; Ndiaye, M.; Oak, M.H.; El Bedoui, J.; Chataigneau, M.; Schini-Kerth, V.B. Vascular protection by dietary polyphenols. Eur. J. Pharmacol. 2004, 500, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.S.; Park, B.J.; Kim, D.H.; Yoo, B.K.; Woo, J.S.; Bhamdari, K.; Jahng, Y.; Choi, H.G.; Lee, M.H. Short communication: In vivo evaluation of microemulsion system for oral and parenteral delivery of rutaecarpine to rats. Drug Dev. Ind. Pharm. 2007, 33, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.C.; Tsai, T.H.; Chou, C.J.; Hsu, S.Y.; Li, S.Y.; Chen, C.F. High-performance liquid chromatographic determination of rutaecarpine in rat plasma: Application to a pharmacokinetic study. J. Chromatogr. B Biomed. Appl. 1994, 655, 27–31. [Google Scholar] [CrossRef]
- Ghosh, S.; Hayden, M.S. New regulators of NF-kappaB in inflammation. Nat. Rev. Immunol. 2008, 8, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Kojok, K.; El-Kadiry, A.E.; Merhi, Y. Role of NF-κB in platelet function. Int. J. Mol. Sci. 2019, 20, 4185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, F.Y.; Davidson, S.J.; Moraes, L.A.; Traves, S.L.; Paul-Clark, M.; Bishop-Bailey, D.; Warner, T.D.; Mitchell, J.A. Role of nuclear receptor signaling in platelets: Antithrombotic effects of PPARbeta. FASEB J. 2006, 20, 326–328. [Google Scholar] [CrossRef]
- Moraes, L.A.; Paul-Clark, M.J.; Rickman, A.; Flower, R.J.; Goulding, N.J.; Perretti, M. Ligand-specific glucocorticoid receptor activation in human platelets. Blood 2005, 106, 4167–4175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaver, E.; Romaniuk, M.A.; D’Atri, L.P.; Pozner, R.G.; Negrotto, S.; Benzadon, R.; Schattner, M. NF-kappaB inhibitors impair platelet activation responses. J. Thromb. Haemost. 2009, 7, 1333–1343. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Naik, U.P. Platelet MAPKs-a 20+ year history: What do we really know? J. Thromb. Haemost. 2020, 18, 2087–2102. [Google Scholar] [CrossRef] [PubMed]
- Mazharian, A.; Roger, S.; Maurice, P.; Berrou, E.; Popoff, M.R.; Hoylaerts, M.F.; Fauvel-Lafeve, F.; Bonnefoy, A.; Bryckaert, M. Differential involvement of ERK2 and p38 in platelet adhesion to collagen. J. Biol. Chem. 2005, 280, 26002–26010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, P.; Zhang, L.; Zhang, M.; Yang, W.; Wang, K.; Zhang, J.; Otsu, K.; Huang, G.; Fan, X.; Liu, J. Platelet-specific p38alpha deficiency improved cardiac function after myocardial infarction in mice. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, M.U.; Patel, P.; Derstine, R.; Turaga, R.; Chen, X.; Golla, K.; Neeves, K.B.; Ichijo, H.; Naik, U.P. Ask1 regulates murine platelet granule secretion, thromboxane A2 generation, and thrombus formation. Blood 2017, 129, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Qiao, J.; Arthur, J.F.; Gardiner, E.E.; Andrews, R.K.; Zeng, L.; Xu, K. Regulation of platelet activation and thrombus formation by reactive oxygen species. Redox Biol. 2018, 14, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Lee, J.J.; Chang, C.C.; Lin, K.H.; Wang, S.H.; Sheu, J.R. Platelet protease-activated receptor (PAR)4, but not PAR1, associated with neutral sphingomyelinase responsible for thrombin-stimulated ceramide-NF-κB signaling in human platelets. Haematologica 2013, 98, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Ragab, A.; Severin, S.; Gratacap, M.P.; Aguado, E.; Malissen, M.; Jandrot-Perrus, M.; Malissen, B.; Ragab-Thomas, J.; Payrastre, B. Roles of the C-terminal tyrosine residues of LAT in GPVI-induced platelet activation: Insights into the mechanism of PLC gamma 2 activation. Blood 2007, 110, 2466–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.-C.; Hou, S.-M.; Wu, M.-P.; Hsia, C.-W.; Jayakumar, T.; Hsia, C.-H.; Bhavan, P.S.; Chung, C.-L.; Sheu, J.-R. Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF-κB. Molecules 2022, 27, 476. https://doi.org/10.3390/molecules27020476
Huang W-C, Hou S-M, Wu M-P, Hsia C-W, Jayakumar T, Hsia C-H, Bhavan PS, Chung C-L, Sheu J-R. Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF-κB. Molecules. 2022; 27(2):476. https://doi.org/10.3390/molecules27020476
Chicago/Turabian StyleHuang, Wei-Chieh, Shaw-Min Hou, Ming-Ping Wu, Chih-Wei Hsia, Thanasekaran Jayakumar, Chih-Hsuan Hsia, Periyakali Saravana Bhavan, Chi-Li Chung, and Joen-Rong Sheu. 2022. "Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF-κB" Molecules 27, no. 2: 476. https://doi.org/10.3390/molecules27020476
APA StyleHuang, W.-C., Hou, S.-M., Wu, M.-P., Hsia, C.-W., Jayakumar, T., Hsia, C.-H., Bhavan, P. S., Chung, C.-L., & Sheu, J.-R. (2022). Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF-κB. Molecules, 27(2), 476. https://doi.org/10.3390/molecules27020476