
Aspergillus ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential

 ,
,  and
and 
Abstract
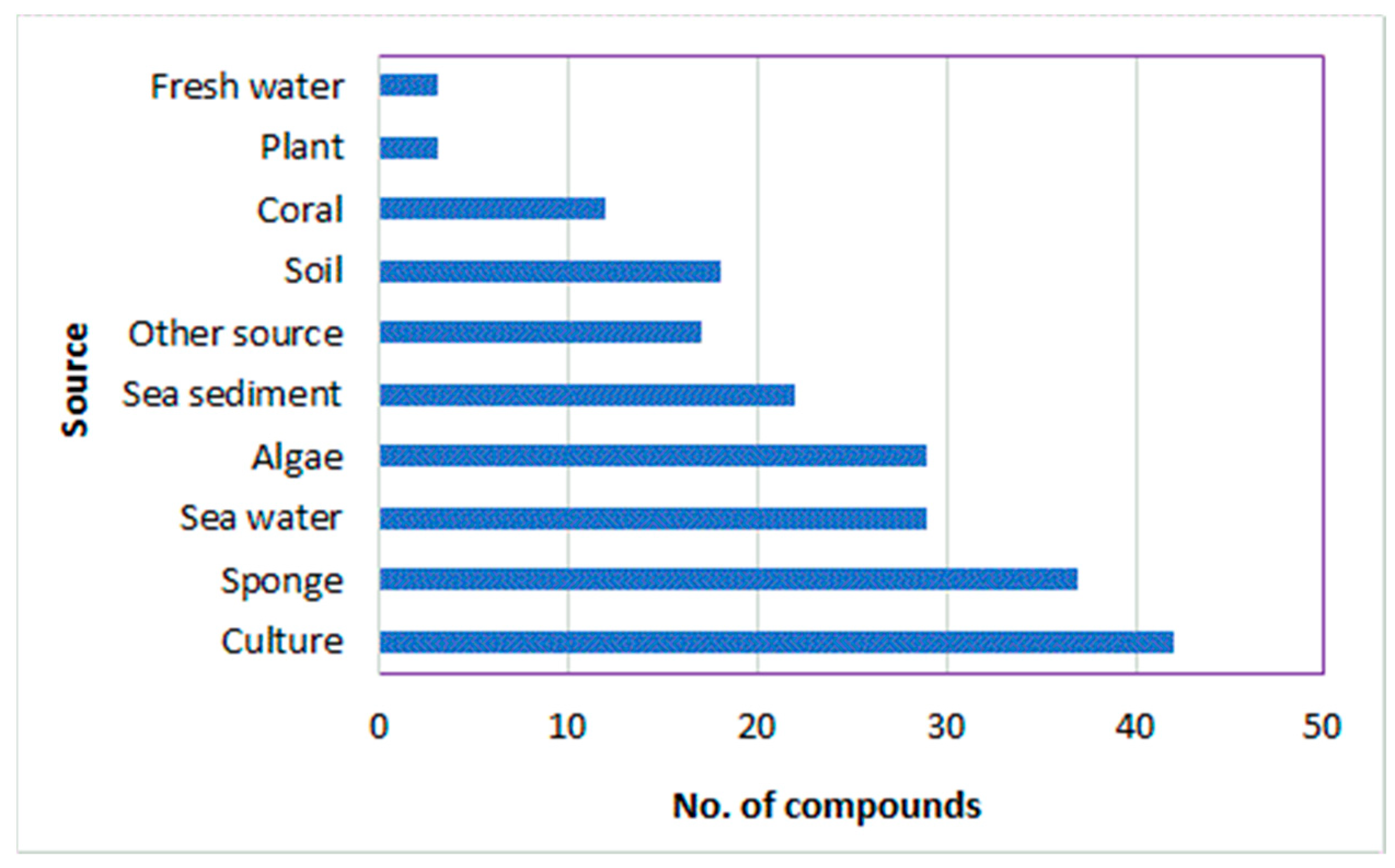
1. Introduction
2. Enzymes of A. ochraceus and Their Applications
2.1. Hydrolases
2.1.1. Glycoside Hydrolases
2.1.2. Proteolytic Enzymes
2.1.3. Tannases
2.2. Oxidases
3. Applications of A. ochraceus
3.1. Bioethanol Production
3.2. Dye Decolorization
3.3. Kerosene Biodegradation
3.4. Nanoparticles (NPs) of A. ochraceus
4. Secondary Metabolites from A. ochraceus and Their Bioactivities
4.1. Isocoumarin Derivatives
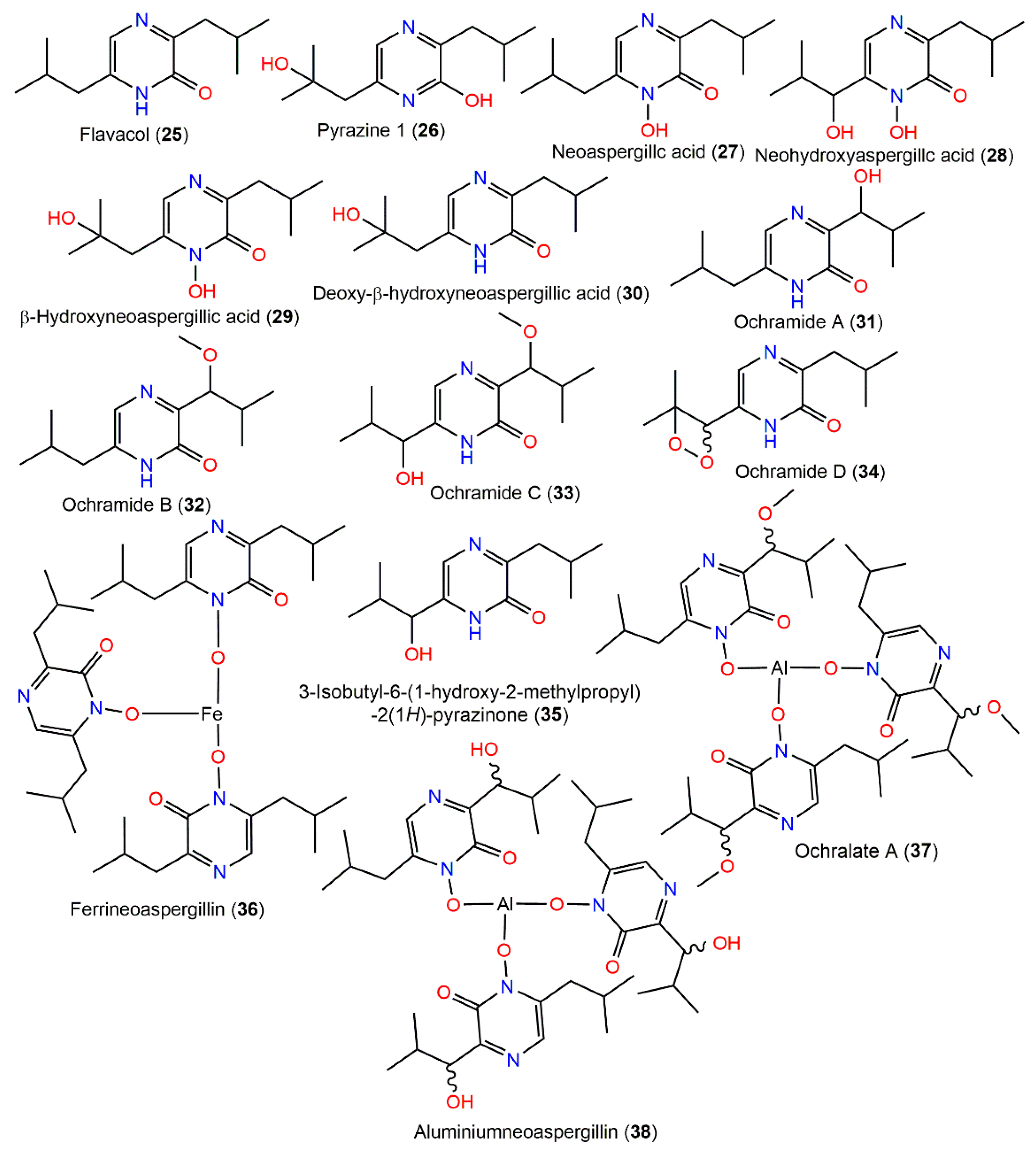
4.2. Pyrazine Derivatives
4.3. Diketopiperazines
4.4. Benzodiazepine Derivatives
4.5. Indole and Other Alkaloids
4.6. Peptides
4.7. Sesquiterpenoids
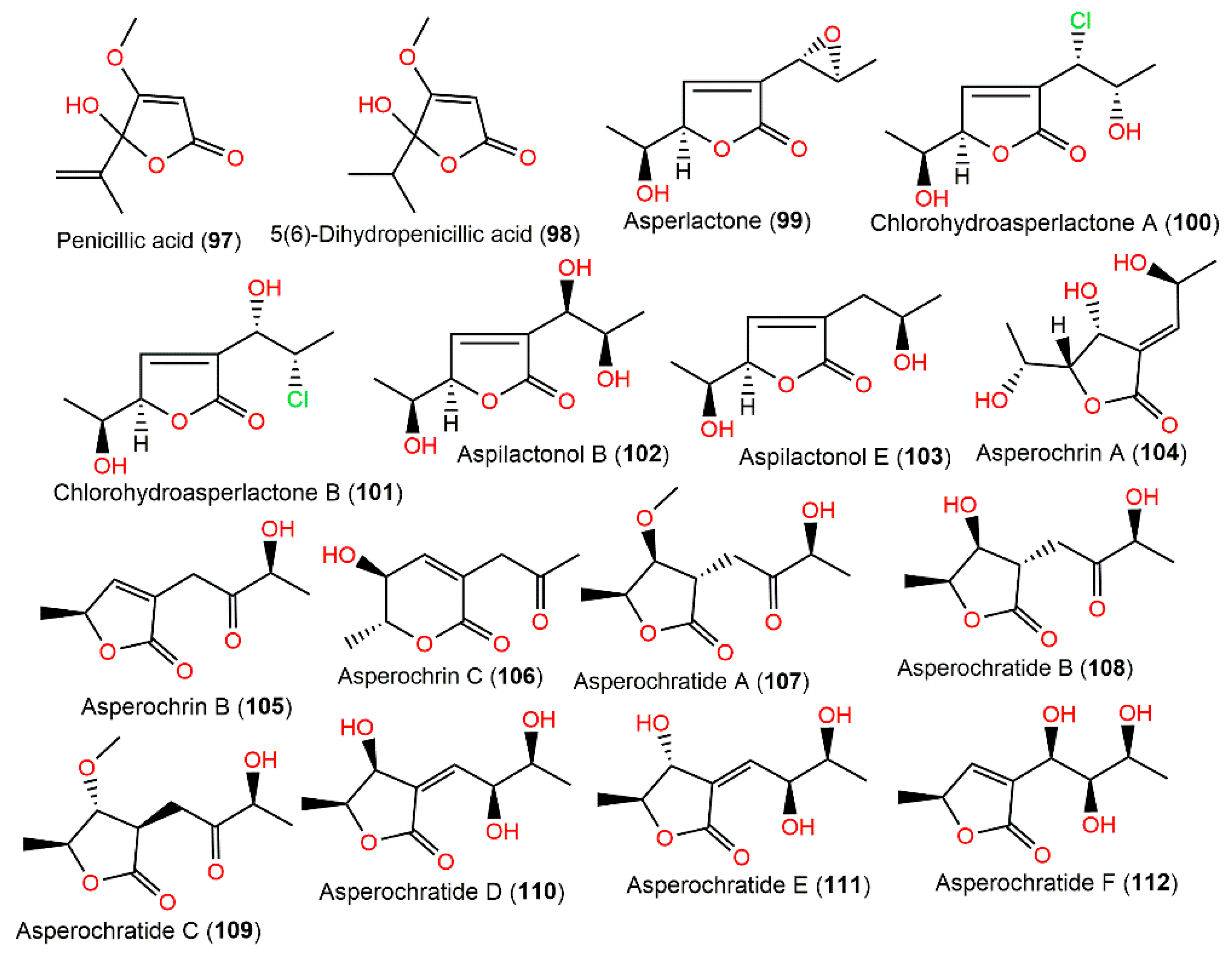
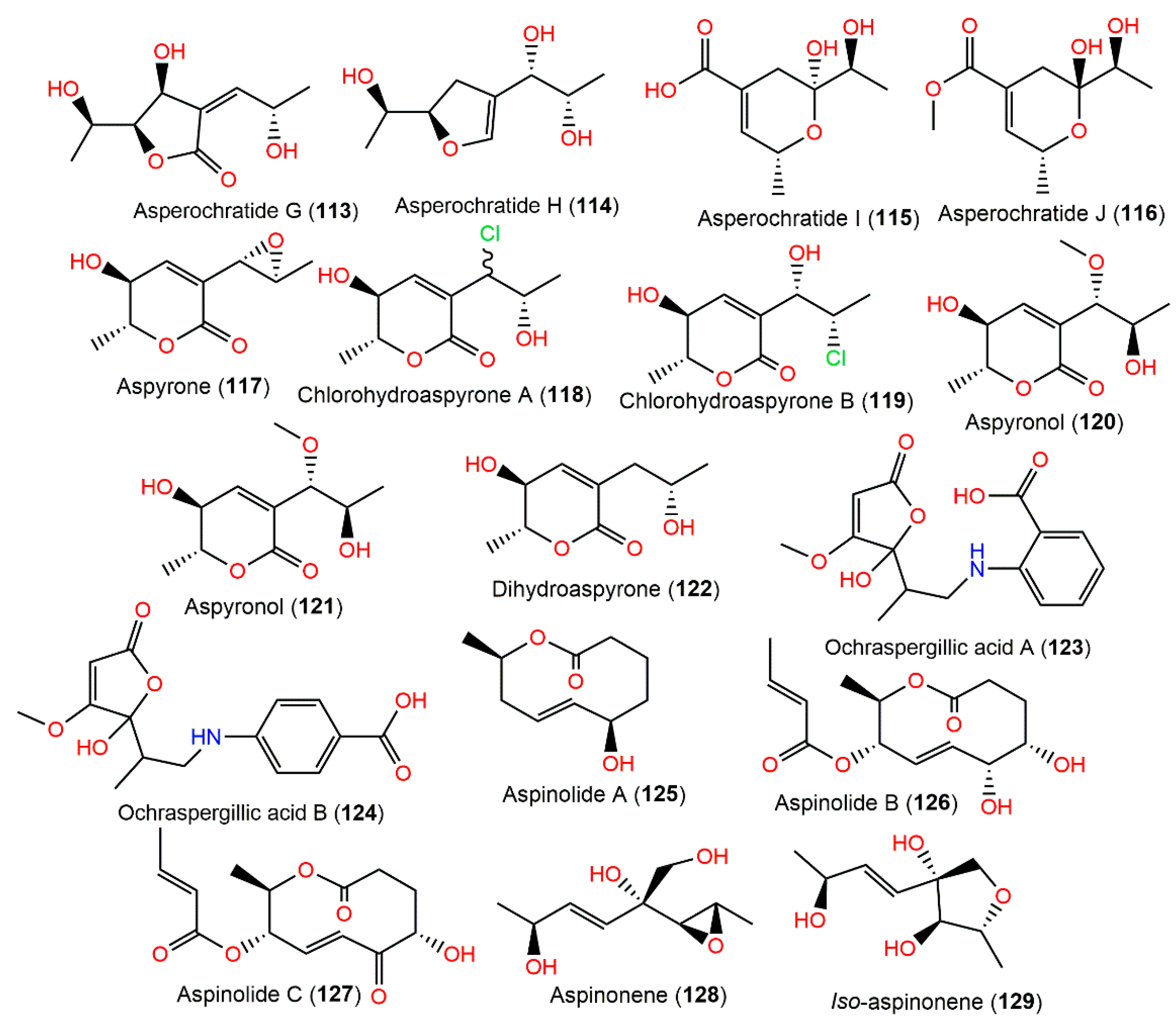
4.8. Polyketides
4.9. Xanthine and Quinone Derivatives and Flavonoids
4.10. Sterols
4.11. Other Metabolites
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Omar, A.M.; Mohamed, G.A.; Ibrahim, S.R.M. Chaetomugilins and Chaetoviridins-Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics. J. Fungi 2022, 8, 127. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Depsides-Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities. Metabolites 2021, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically Active Fungal Depsidones: Chemistry, Biosynthesis, Structural Characterization, and Bioactivities. Fitoterapia 2018, 129, 317–365. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.; Fadil, S.A.; Fadil, H.A.; Hareeri, R.H.; Alolayan, S.O.; Abdallah, H.M.; Mohamed, G.A. Dactylospongia elegans—A Promising Drug Source: Metabolites, Bioactivities, Biosynthesis, Synthesis, and Structural-Activity Relationship. Mar. Drugs 2022, 20, 221. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.; Choudhry, H.; Asseri, A.H.; Elfaky, M.A.; Mohamed, S.G.; Mohamed, G.A. Stachybotrys chartarum—A Hidden Treasure: Secondary Metabolites, Bioactivities, and Biotechnological Relevance. J. Fungi 2022, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- Greco, C.; Keller, N.P.; Rokas, A. Unearthing Fungal Chemodiversity and Prospects for Drug Discovery. Curr. Opin. Microbiol. 2019, 51, 22–29. [Google Scholar] [CrossRef]
- Romero, S.M.; Giudicessi, S.L.; Vitale, R.G. Is the Fungus Aspergillus a Threat to Cultural Heritage? J. Cult. Herit. 2021, 51, 107–124. [Google Scholar] [CrossRef]
- El-Agamy, D.S.; Ibrahim, S.R.; Ahmed, N.; Khoshhal, S.; Abo-Haded, H.M.; Elkablawy, M.A.; Aljuhani, N.; Mohamed, G.A. Aspernolide F, as a New Cardioprotective Butyrolactone Against Doxorubicin-Induced Cardiotoxicity. Int. Immunopharmacol. 2019, 72, 429–436. [Google Scholar] [CrossRef]
- Elkhayat, E.S.; Ibrahim, S.R.; Mohamed, G.A.; Ross, S.A. Terrenolide S, a New Antileishmanial Butenolide from the Endophytic Fungus Aspergillus terreus. Nat. Prod. Res. 2016, 30, 814–820. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Mohamed, G.A.; Khedr, A.I. Γ-Butyrolactones from Aspergillus Species: Structures, Biosynthesis, and Biological Activities. Nat. Prod. Commun. 2017, 12, 791–800. [Google Scholar] [CrossRef]
- Ahluwalia, S.K.; Matsui, E.C. Indoor Environmental Interventions for Furry Pet Allergens, Pest Allergens, and Mold: Looking to the Future. J. Allergy Clin. Immunol. Pract. 2018, 6, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.H.; Balbool, B.A.; Abdel-Azeem, A.M. Aspergillus from different habitats and their industrial applications. In Industrially Important Fungi for Sustainable Development; Springer International Publishing: Cham, Switzerland, 2021; pp. 85–106. [Google Scholar]
- Hakamifard, A.; Hashemi, M.; Fakhim, H.; Aboutalebian, S.; Hajiahmadi, S.; Mohammadi, R. Fatal Disseminated Aspergillosis in an Immunocompetent Patient with COVID-19 due to Aspergillus ochraceus. J. Med. Mycol. 2021, 31, 101124. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Ibrahim, S.R.; Asfour, H.Z. Antimicrobial Metabolites from the Endophytic Fungus Aspergillus versicolor. Phytochem. Lett. 2020, 35, 152–155. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Z.; Gao, J. Chemistry and Biology of Secondary Metabolites from Aspergillus Genus. Nat. Prod. J. 2018, 8, 275–304. [Google Scholar] [CrossRef]
- Cole, R.J.; Moore, J.H.; Davis, N.D.; Kirksey, J.W.; Diener, U.L. 4-Hydroxymellein. New Metabolite of Aspergillus ochraceus. J. Agric. Food Chem. 1971, 19, 909–911. [Google Scholar] [CrossRef]
- Frank, M.; Özkaya, F.C.; Müller, W.E.; Hamacher, A.; Kassack, M.U.; Lin, W.; Liu, Z.; Proksch, P. Cryptic Secondary Metabolites from the Sponge-Associated Fungus Aspergillus ochraceus. Mar. Drugs 2019, 17, 99. [Google Scholar] [CrossRef]
- Stoev, S.D.; Vitanov, S.; Anguelov, G.; Petkova-Bocharova, T.; Creppy, E.E. Experimental Mycotoxic Nephropathy in Pigs Provoked by a Diet Containing Ochratoxin A and Penicillic Acid. Vet. Res. Commun. 2001, 25, 205–223. [Google Scholar] [CrossRef]
- Stoev, S.D. Balkan Endemic Nephropathy–Still Continuing Enigma, Risk Assessment and Underestimated Hazard of Joint Mycotoxin Exposure of Animals Or Humans. Chem. Biol. Interact. 2017, 261, 63–79. [Google Scholar] [CrossRef]
- Dai, J.; Carté, B.K.; Sidebottom, P.J.; Sek Yew, A.L.; Ng, S.; Huang, Y.; Butler, M.S. Circumdatin G, a New Alkaloid from the Fungus Aspergillus ochraceus. J. Nat. Prod. 2001, 64, 125–126. [Google Scholar] [CrossRef]
- Tong, Z.; Xiao, X.; Lu, Y.; Zhang, Y.; Hu, P.; Jiang, W.; Zhou, H.; Pan, S.; Huang, Z.; Hu, L. New Metabolites from Aspergillus ochraceus with Antioxidative Activity and Neuroprotective Potential on H2O2 Insult SH-SY5Y Cells. Molecules 2022, 27, 52. [Google Scholar] [CrossRef]
- Peng, X.; Wang, Y.; Zhu, T.; Zhu, W. Pyrazinone Derivatives from the Coral-Derived Aspergillus ochraceus LCJ11-102 under High Iodide Salt. Arch. Pharm. Res. 2018, 41, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yuan, C.; Zhang, J.; Liu, S.; Cao, P.; Hua, H.; Di, Y.; Hao, X. Speramides A–B, Two New Prenylated Indole Alkaloids from the Freshwater-Derived Fungus Aspergillus ochraceus KM007. Tetrahedron. Lett. 2016, 57, 4952–4955. [Google Scholar] [CrossRef]
- Hu, L.; Tian, S.; Wu, R.; Tong, Z.; Jiang, W.; Hu, P.; Xiao, X.; Zhang, X.; Zhou, H.; Tong, Q. Identification of Anti-parkinson’s Disease Lead Compounds from Aspergillus ochraceus Targeting Adenosin Receptors A2A. ChemistryOpen 2021, 10, 630–638. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Jensen, P.R.; Renner, M.K.; Fenical, W. New Cytotoxic Sesquiterpenoid Nitrobenzoyl Esters from a Marine Isolate of the Fungus Aspergillus versicolor. Tetrahedron 1998, 54, 1715–1724. [Google Scholar] [CrossRef]
- Cui, C.; Li, X.; Li, C.; Sun, H.; Gao, S.; Wang, B. Benzodiazepine Alkaloids from Marine-derived Endophytic Fungus Aspergillus ochraceus. Helv. Chim. Acta 2009, 92, 1366–1370. [Google Scholar] [CrossRef]
- Cui, C.; Li, X.; Meng, L.; Li, C.; Huang, C.; Wang, B. 7-nor-Ergosterolide, a Pentalactone-Containing Norsteroid and Related Steroids from the Marine-Derived Endophytic Aspergillus ochraceus EN-31. J. Nat. Prod. 2010, 73, 1780–1784. [Google Scholar] [CrossRef] [PubMed]
- De Guzman, F.S.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. New Diketopiperazine Metabolites from the Sclerotia of Aspergillus ochraceus. J. Nat. Prod. 1992, 55, 931–939. [Google Scholar] [CrossRef]
- De Guzman, F.S.; Bruss, D.R.; Rippentrop, J.M.; Gloer, K.B.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Ochrindoles A-D: New Bis-Indolyl Benzenoids from the Sclerotia of Aspergillus ochraceus NRRL 3519. J. Nat. Prod. 1994, 57, 634–639. [Google Scholar] [CrossRef]
- Delgadillo, I. Isolation of Secondary Metabolites of Aspergillus ochraceus by HPLC. Mycotoxin Res. 1986, 2, 9–17. [Google Scholar] [CrossRef]
- Fan, Y.; Zhou, Y.; Du, Y.; Wang, Y.; Fu, P.; Zhu, W. Circumdatin-Aspyrone Conjugates from the Coral-Associated Aspergillus ochraceus LCJ11-102. Mar. Drugs 2019, 17, 400. [Google Scholar] [CrossRef]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Lin, J.; Zhang, T.; Tu, Z. Cytotoxic and Antiviral Nitrobenzoyl Sesquiterpenoids from the Marine-Derived Fungus Aspergillus ochraceus Jcma1F17. MedChemComm 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Fuchser, J.; Zeeck, A. Aspinolides and Aspinonene/Aspyrone Co-Metabolites, New Pentaketides Produced by Aspergillus ochraceus. Liebigs Ann./Recl. 1997, 1, 87–95. [Google Scholar] [CrossRef]
- González-Jartín, J.M.; Alfonso, A.; Sainz, M.J.; Vieytes, M.R.; Botana, L.M. UPLC–MS–IT–TOF Identification of Circumdatins Produced by Aspergillus ochraceus. J. Agric. Food Chem. 2017, 65, 4843–4852. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Mao, W.; Yan, M.; Zhao, C.; Li, N.; Shan, J.; Lin, C.; Liu, X.; Guo, T.; Guo, T. Galactomannan with Novel Structure Produced by the Coral Endophytic Fungus Aspergillus ochraceus. Carbohydr. Polym. 2014, 105, 325–333. [Google Scholar] [CrossRef]
- Aracri, F.M.; Cavalcanti, R.M.; Guimaraes, L.H.S. Extracellular Tannase from Aspergillus ochraceus: Influence of the Culture Conditions on Biofilm Formation, Enzyme Production, and Application. J. Microbiol. Biotechnol. 2019, 29, 1749–1759. [Google Scholar] [CrossRef]
- Asha, P.; Divya, J.; Singh, I.B. Purification and Characterisation of Processive-Type Endoglucanase and Β-Glucosidase from Aspergillus ochraceus MTCC 1810 through Saccharification of Delignified Coir Pith to Glucose. Bioresour. Technol. 2016, 213, 245–248. [Google Scholar] [CrossRef]
- Batomunkueva, B.P.; Egorov, N.S. Isolation, Purification, and Resolution of the Extracellular Proteinase Complex of Aspergillus ochraceus 513 with Fibrinolytic and Anticoagulant Activities. Microbiology 2001, 70, 519–522. [Google Scholar] [CrossRef]
- Betini, J.H.A.; Michelin, M.; Peixoto-Nogueira, S.D.C.; Jorge, J.A.; Terenzi, H.F.; Polizeli, M.L.T.M. Xylanases from Aspergillus niger, Aspergillus niveus and Aspergillus ochraceus Produced under Solid-State Fermentation and their Application in Cellulose Pulp Bleaching. Bioprocess Biosyst. Eng. 2009, 32, 819–824. [Google Scholar] [CrossRef]
- Biswas, S.R.; Jana, S.C.; Mishra, A.K.; Nanda, G. Production, Purification, and Characterization of Xylanase from a Hyperxylanolytic Mutant of Aspergillus ochraceus. Biotechnol. Bioeng. 1990, 35, 244–251. [Google Scholar] [CrossRef]
- Ghosh, K.; Dhar, A.; Samanta, T.B. Purification and Characterization of an Invertase Produced by Aspergillus ochraceus TS. Indian J. Biochem. Biophys 2001, 38, 180–185. [Google Scholar]
- Gonçalves, H.B.; Riul, A.J.; Quiapim, A.C.; Jorge, J.A.; Guimarães, L.H.S. Characterization of a Thermostable Extracellular Tannase Produced Under Submerged Fermentation by Aspergillus ochraceus. EJB 2012, 15, 4. [Google Scholar]
- Ibrahim, S.R.; Mohamed, S.G.; Altyar, A.E.; Mohamed, G.A. Natural Products of the Fungal Genus Humicola: Diversity, Biological Activity, and Industrial Importance. Curr. Microbiol. 2021, 78, 2488–2509. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.; Altyar, A.E.; Mohamed, S.G.; Mohamed, G.A. Genus Thielavia: Phytochemicals, Industrial Importance and Biological Relevance. Nat. Prod. Res. 2021, 36, 5108–5123. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Sustainability of Cereal Straws for the Fermentative Production of Second Generation Biofuels: A Review of the Efficiency and Economics of Biochemical Pretreatment Processes. Appl. Energy 2017, 198, 284–298. [Google Scholar] [CrossRef]
- Sharma, N.; Bhardwaj, N.K.; Singh, R.B.P. Environmental Issues of Pulp Bleaching and Prospects of Peracetic Acid Pulp Bleaching: A Review. J. Clean. Prod. 2020, 256, 120338. [Google Scholar] [CrossRef]
- Haile, A.; Gelebo, G.G.; Tesfaye, T.; Mengie, W.; Mebrate, M.A.; Abuhay, A.; Limeneh, D.Y. Pulp and Paper Mill Wastes: Utilizations and Prospects for High Value-Added Biomaterials. Bioresour. Bioprocess. 2021, 8, 1–22. [Google Scholar] [CrossRef]
- Michelin, M.; Peixoto-Nogueira, S.C.; Betini, J.; Da Silva, T.M.; Jorge, J.A.; Terenzi, H.F.; Polizeli, M. Production and Properties of Xylanases from Aspergillus terricola Marchal and Aspergillus ochraceus and their use in Cellulose Pulp Bleaching. Bioprocess Biosyst. Eng. 2010, 33, 813–821. [Google Scholar] [CrossRef]
- Michelin, M.; Polizeli, M.D.L.; Ruzene, D.S.; Silva, D.P.; Vicente, A.A.; Jorge, J.A.; Terenzi, H.F.; Teixeira, J.A. Xylanase and Β-Xylosidase Production by Aspergillus ochraceus: New Perspectives for the Application of Wheat Straw Autohydrolysis Liquor. Appl. Biochem. Biotechnol. 2012, 166, 336–347. [Google Scholar] [CrossRef]
- Veana, F.; Flores-Gallegos, A.C.; Gonzalez-Montemayor, A.M.; Michel-Michel, M.; Lopez-Lopez, L.; Aguilar-Zarate, P.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R. Invertase: An enzyme with importance in confectionery food industry. In Enzymes in Food Technology; Springer: Cham, Switzerland, 2018; pp. 187–212. [Google Scholar]
- Guimarães, L.H.S.; Terenzi, H.F.; de Moraes, M.D.L.T.; Jorge, J.A. Production and Characterization of a Thermostable Extracellular Β-D-Fructofuranosidase Produced by Aspergillus ochraceus with Agroindustrial Residues as Carbon Sources. Enzym. Microb. Technol. 2007, 42, 52–57. [Google Scholar] [CrossRef]
- Kotb, E. The Biotechnological Potential of Fibrinolytic Enzymes in the Dissolution of Endogenous Blood Thrombi. Biotechnol. Prog. 2014, 30, 656–672. [Google Scholar] [CrossRef]
- Pascreau, T.; de la Morena-Barrio, M.E.; Lasne, D.; Serrano, M.; Bianchini, E.; Kossorotoff, M.; Boddaert, N.; Bruneel, A.; Seta, N.; Vicente, V. Elevated Thrombin Generation in Patients with Congenital Disorder of Glycosylation and Combined Coagulation Factor Deficiencies. J. Thromb. Haemost. 2019, 17, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Marlar, R.A.; Kleiss, A.J.; Griffin, J.H. Mechanism of Action of Human Activated Protein C, a Thrombin-Dependent Anticoagulant Enzyme. Blood 1982, 59, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Osmolovskiy, A.A.; Rukavitsyna, E.D.; Kreier, V.G.; Baranova, N.A.; Egorov, N.S. Production of Proteinases with Fibrinolytic and Fibrinogenolytic Activity by a Micromycete Aspergillus ochraceus. Microbiology 2017, 86, 512–516. [Google Scholar] [CrossRef]
- Chávez-González, M.; Rodríguez-Durán, L.V.; Balagurusamy, N.; Prado-Barragán, A.; Rodríguez, R.; Contreras, J.C.; Aguilar, C.N. Biotechnological Advances and Challenges of Tannase: An Overview. Food Bioproc. Tech. 2012, 5, 445–459. [Google Scholar] [CrossRef]
- Belmares, R.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Coronel, A.R.; Aguilar, C.N. Microbial Production of Tannase: An Enzyme with Potential use in Food Industry. LWT-Food Sci. Technol. 2004, 37, 857–864. [Google Scholar] [CrossRef]
- Banerjee, D.; Mahapatra, S. Fungal Tannase: A Journey from Strain Isolation to Enzyme Applications. Dyn. Biochem. Proc. Biotechnol. Mol. Biol. 2012, 6, 49–60. [Google Scholar]
- Goswami, P.; Chinnadayyala, S.S.R.; Chakraborty, M.; Kumar, A.K.; Kakoti, A. An Overview on Alcohol Oxidases and their Potential Applications. Appl. Microbiol. Biotechnol. 2013, 97, 4259–4275. [Google Scholar] [CrossRef]
- Isobe, K.; Kato, A.; Ogawa, J.; Kataoka, M.; Iwasaki, A.; Hasegawa, J.; Shimizu, S. Characterization of Alcohol Oxidase from Aspergillus ochraceus AIU 031. J. Gen. Appl. Microbiol. 2007, 53, 177–183. [Google Scholar] [CrossRef]
- Gupta, A.; Verma, J.P. Sustainable Bio-Ethanol Production from Agro-Residues: A Review. Renew. Sust. Energ. Rev. 2015, 41, 550–567. [Google Scholar] [CrossRef]
- Rathna, G.S.; Saranya, R.; Kalaiselvam, M. Bioethanol from Sawdust using Cellulase Hydrolysis of Aspergillus ochraceus and Fermentation by Saccharomyces cerevisiae. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 733–742. [Google Scholar]
- Kadam, A.A.; Telke, A.A.; Jagtap, S.S.; Govindwar, S.P. Decolorization of Adsorbed Textile Dyes by Developed Consortium of Pseudomonas sp. SUK1 and Aspergillus ochraceus NCIM-1146 under Solid State Fermentation. J. Hazard. Mater. 2011, 189, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Saratale, G.D.; Kalme, S.D.; Govindwar, S.P. Decolorisation of Textile Dyes by Aspergillus ochraceus (NCIM-1146). J. Basic Microbiol. 2006, 5, 407–410. [Google Scholar]
- Telke, A.A.; Kadam, A.A.; Jagtap, S.S.; Jadhav, J.P.; Govindwar, S.P. Biochemical Characterization and Potential for Textile Dye Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146. Biotechnol. Bioprocess Eng. 2010, 15, 696–703. [Google Scholar] [CrossRef]
- Abha, S.; Singh, C.S. Hydrocarbon Pollution: Effects on Living Organisms, Remediation of Contaminated Environments, and Effects of Heavy Metals Co-Contamination on Bioremediation. In Introduction to Enhanced Oil Recovery (EOR) Processes and Bioremediation of Oil-Contaminated Sites; Romero-Zerón, L., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 186–206. [Google Scholar]
- Darsa, K.V.; Thatheyus, A.J.; Ramya, D. Biodegradation of Petroleum Compound using the Bacterium Bacillus subtilis. Sci. Int. 2014, 2, 20–25. [Google Scholar] [CrossRef]
- Saratale, G.; Kalme, S.; Bhosale, S.; Govindwar, S. Biodegradation of Kerosene by Aspergillus ochraceus NCIM-1146. J. Basic Microbiol. 2007, 47, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Sebiomo, A.; Awosanya, A.O.; Awofodu, A.D. Utilization of Crude Oil and Gasoline by Ten Bacterial and Five Fungal Isolates. J. Microbiol. Antimicrob. 2011, 3, 55–63. [Google Scholar]
- Vijayakumar, P.S.; Prasad, B. Intracellular Biogenic Silver Nanoparticles for the Generation of Carbon Supported Antiviral and Sustained Bactericidal Agents. Langmuir 2009, 25, 11741–11747. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; Meng, L.; Wang, B. Polyketides from the Marine Mangrove-Derived Fungus Aspergillus ochraceus MA-15 and their Activity Against Aquatic Pathogenic Bacteria. Phytochem. Lett. 2015, 12, 232–236. [Google Scholar] [CrossRef]
- Yun, K.; Feng, Z.; Choi, H.D.; Kang, J.S.; Son, B.W. New Production of (R)-(–)-5-Bromomellein, a Dihydroisocoumarin Derivative from the Marine-Derived Fungus Aspergillus ochraceus. Chem. Nat. Compd. 2013, 49, 24–26. [Google Scholar] [CrossRef]
- Yamazaki, M.; Maebayashi, Y.; Miyaki, K. Isolation of a New Metabolite, 6-Methoxy-8-Hydroxyisocoumarin-3-Carboxylic Acid from Aspergillus ochraceus WILH. Chem. Pharm. Bull. 1972, 20, 2276–2278. [Google Scholar] [CrossRef]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a Toxic Metabolite Produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.D.; Searcy, J.W.; Diener, U.L. Production of Ochratoxin A by Aspergillus ochraceus in a Semisynthetic Medium. Appl. Microbiol. 1969, 17, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Marquardt, R.R.; Abramson, D.; Frohlich, A.A. Metabolites of Ochratoxins in Rat Urine and in a Culture of Aspergillus ochraceus. Appl. Environ. Microbiol. 1996, 62, 648–655. [Google Scholar] [CrossRef]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins. Part II. the Constitution of Ochratoxins A, B, and C, Metabolites of Aspergillus ochraceus Wilh. J. Chem. Soc. 1965, 1965, 7083–7088. [Google Scholar] [CrossRef]
- Moore, J.H.; Davis, N.D.; Diener, U.L. Mellein and 4-Hydroxymellein Production by Aspergillus ochraceus Wilhelm. Appl. Microbiol. 1972, 23, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; Liu, P.; Wang, Z.; Zhu, W. Effects of Environmental Stress on Secondary Metabolites of Aspergillus ochraceus LCJ11-102 Associated with the Coral Dichotella gemmacea. Acta Microbiol. Sin. 2010, 50, 1023–1029. [Google Scholar]
- Zou, Z.; Zhang, G.; Li, S.; He, Z.; Yan, Q.; Lin, Y.; Xie, C.; Xia, J.; Luo, Z.; Luo, L. Asperochratides A–J, Ten New Polyketides from the Deep-Sea-Derived Aspergillus ochraceus. Bioorg. Chem. 2020, 105, 104349. [Google Scholar] [CrossRef]
- Harris, J.P.; Mantle, P.G. Biosynthesis of Diaporthin and Orthosporin by Aspergillus ochraceus. Phytochemistry 2001, 57, 165–169. [Google Scholar] [CrossRef]
- Stander, M.A.; Steyn, P.S.; Lübben, A.; Miljkovic, A.; Mantle, P.G.; Marais, G.J. Influence of Halogen Salts on the Production of the Ochratoxins by Aspergillus ochraceus Wilh. J. Agric. Food Chem. 2000, 48, 1865–1871. [Google Scholar] [CrossRef]
- Hadidane, R.; Bacha, H.; Creppy, E.E.; Hammami, M.; Ellouze, F.; Dirheimer, G. Isolation and Structure Determination of Natural Analogues of the Mycotoxin Ochratoxin A Produced by Aspergillus ochraceus. Toxicology 1992, 76, 233–243. [Google Scholar] [CrossRef]
- Yamazaki, M.; Maebayashi, Y.; Miyaki, K. Isolation of a New Type of Pyrazine Metabolite from Aspergillus ochraceus WILH. Chem. Pharm. Bull. 1972, 20, 2274–2276. [Google Scholar] [CrossRef]
- Maebayashi, Y.; Sumita, M.; Fukushima, K.; Yamasaki, M. Isolation and Structure of Red Pigment from Aspergillus ochraceus Wilh. Chem. Pharm. Bull. 1978, 26, 1320–1322. [Google Scholar] [CrossRef][Green Version]
- López-Gresa, M.P.; González, M.C.; Primo, J.; Moya, P.; Romero, V.; Estornell, E. Circumdatin H, a New Inhibitor of Mitochondrial NADH Oxidase, from Aspergillus ochraceus. J. Antibiot. 2005, 58, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.K.; Størmer, F.C.; Petersen, D.; Aasen, A.J. Cycloechinulin, a Trp-Ala-Derived Alkaloid from the Fungus Aspergillus ochraceus. Acta Crystallogr. Sect. E Struct. Rep. Online 2001, 57, o909–o912. [Google Scholar] [CrossRef]
- Sugie, Y.; Hirai, H.; Inagaki, T.; Ishiguro, M.; Kim, Y.; Kojima, Y.; Sakakibara, T.; Sakemi, S.; Sugiura, A.; Suzuki, Y. A New Antibiotic CJ-17, 665 from Aspergillus ochraceus. J. Antibiot. 2001, 54, 911–916. [Google Scholar] [CrossRef]
- Qian-Cutrone, J.; Huang, S.; Shu, Y.; Vyas, D.; Fairchild, C.; Menendez, A.; Krampitz, K.; Dalterio, R.; Klohr, S.E.; Gao, Q. Stephacidin A and B: Two Structurally Novel, Selective Inhibitors of the Testosterone-Dependent Prostate LNCaP Cells. J. Am. Chem. Soc. 2002, 124, 14556–14557. [Google Scholar] [CrossRef]
- Wen, H.; Liu, X.; Zhang, Q.; Deng, Y.; Zang, Y.; Wang, J.; Liu, J.; Zhou, Q.; Hu, L.; Zhu, H. Three New Indole Diketopiperazine Alkaloids from Aspergillus ochraceus. Chem. Biodivers. 2018, 15, e1700550. [Google Scholar] [CrossRef]
- Rahbaek, L.; Breinholt, J.; Frisvad, J.C.; Christophersen, C. Circumdatin A, B, and C: Three New Benzodiazepine Alkaloids Isolated from a Culture of the Fungus Aspergillus ochraceus. J. Org. Chem. 1999, 64, 1689–1692. [Google Scholar] [CrossRef]
- Rahbæk, L.; Breinholt, J. Circumdatins D, E, and F: Further Fungal Benzodiazepine Analogues from Aspergillus ochraceus. J. Nat. Prod. 1999, 62, 904–905. [Google Scholar] [CrossRef]
- Schwartz, R.E.; Liesch, J.; Hensens, O.; Zitano, L.; Honeycutt, S.; Garrity, G.; Fromtling, R.A.; Onishi, J.; Monaghan, R. L-657, 398, a Novel Antifungal Agent: Fermentation, Isolation, Structural Elucidation and Biological Properties. J. Antibiot. 1988, 41, 1774–1779. [Google Scholar] [CrossRef]
- Chang, C.; Myokei, R.; Sakurai, A.; Takahashi, N.; Tamura, S. Aspochracin, a New Insecticidal Metabolite of Aspergillus ochraceus Part II. Synthesis of Hexahydroaspochracin. Agric. Biol. Chem. 1969, 33, 1501–1506. [Google Scholar] [CrossRef]
- Myokei, R.; Sakurai, A.; Chang, C.; Kodaira, Y.; Takahashi, N.; Tamura, S. Aspochracin, a New Insecticidal Metabolite of Aspergillus ochraceus: Part I. Isolation, Structure and Biological Activities. Agric. Biol. Chem. 1969, 33, 1491–1500. [Google Scholar]
- Myokei, R.; Sakurai, A.; Chang, C.; Kodaira, Y.; Takahashi, N.; Tamura, S. Structure of Aspochracin, an Insecticidal Metabolite of Aspergillus ochraceus. Tetrahedron Lett. 1969, 10, 695–698. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, B.; Lin, X.; Luo, X.; Pang, X.; Tang, L.; Liu, Y.; Li, X.; Zhou, X. Nitrobenzoyl Sesquiterpenoids with Cytotoxic Activities from a Marine-Derived Aspergillus ochraceus Fungus. J. Nat. Prod. 2018, 81, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Obana, H.; Kumeda, Y.; Nishimune, T. Aspergillus ochraceus Production of 5, 6-Dihydropenicillic Acid in Culture and Foods. J. Food Prot. 1995, 58, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Balcells, M.; Sala, N.; Sanchis, V.; Canela, R. Bactericidal and Fungicidal Activity of Aspergillus ochraceus Metabolites and some Derivatives. Pestic. Sci. 1998, 53, 9–14. [Google Scholar] [CrossRef]
- Fuchser, J.; Thiericke, R.; Zeeck, A. Biosynthesis of Aspinonene, a Branched Pentaketide Produced by Aspergillus ochraceus, Related to Aspyrone. J. Chem. Soc. Perkin Trans. 1 1995, 13, 1663–1666. [Google Scholar] [CrossRef]
- Simpson, T.J.; Holker, J.S. The Biosynthesis of a Pyrone Metabolite of Aspergillus melleus an Application of Long-Range 13C-13C Coupling Constants. Tetrahedron Lett. 1975, 16, 4693–4696. [Google Scholar] [CrossRef]
- Moore, J.H.; Murray, T.P.; Marks, M.E. Production of 3-(1,2-Epoxypropyl)-5, 6-Dihydro-5-Hydroxy-6-Methylpyran-2-One by Aspergillus ochraceus. J. Agric. Food Chem. 1974, 22, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Maebayashi, Y.; Miyaki, K. The Isolation of Secalonic Acid A from Aspergillus ochraceus Cultured on Rice. Chem. Pharm. Bull. 1971, 19, 199–201. [Google Scholar] [CrossRef]
- Lu, P.; Zhao, X.; Cui, T. Production of Emodin from Aspergillus ochraceus at Preparative Scale. Afr. J. Biotechnol. 2010, 9, 512–517. [Google Scholar]
- Robbers, J.E.; Hong, S.; Tuite, J.; Carlton, W.W. Production of Xanthomegnin and Viomellein by Species of Aspergillus Correlated with Mycotoxicosis Produced in Mice. Appl. Environ. Microbiol. 1978, 36, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Stack, M.E.; Mislivec, P.B. Production of Xanthomegnin and Viomellein by Isolates of Aspergillus ochraceus, Penicillium cyclopium, and Penicillium viridicatum. Appl. Environ. Microbiol. 1978, 36, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Stack, M.E.; Mislivec, P.B.; Denizel, T.; Gibson, R.; Pohland, A.E. Ochratoxins A and B, Xanthomegnin, Viomellein and Vioxanthin Production by Isolates of Aspergillus ochraceus from Green Coffee Beans. J. Food Prot. 1983, 46, 965–968. [Google Scholar] [CrossRef]
- Meenupriya, J.; Thangaraj, M. Analytical Characterization and Structure Elucidation of Metabolites from Aspergillus ochraceus MP2 Fungi. Asian Pac. J. Trop. Biomed. 2011, 1, 376–380. [Google Scholar]
- Awad, G.; Mathieu, F.; Coppel, Y.; Lebrihi, A. Characterization and Regulation of New Secondary Metabolites from Aspergillus ochraceus M18 obtained by UV Mutagenesis. Can. J. Microbiol. 2005, 51, 59–67. [Google Scholar] [CrossRef]
- Takase, S.; Kawai, Y.; Uchida, I.; Tanaka, H.; Aoki, H. Structure of Amauromine, a New Hypotensive Vasodilator Produced by Amauroascus sp. Tetrahedron 1985, 41, 3037–3048. [Google Scholar] [CrossRef]
- Tanner, F.W., Jr.; Sobin, B.A.; Gardocki, J.F. Some Chemical and Biologic Properties of Anisomycin. Antibiot. Ann. 1954, 1955, 809–812. [Google Scholar]
- Zhao, H.; Anbuchezhian, R.; Sun, W.; Shao, C.; Zhang, F.; Yin, Y.; Yu, Z.; Li, Z.; Wang, C. Cytotoxic Nitrobenzoyloxy-Substituted Sesquiterpenes from Sponge Derived Endozoic Fungus Aspergillus insulicola md10-2. Curr. Pharm. Biotechnol. 2016, 17, 271–274. [Google Scholar] [CrossRef]
- Franck, B.; Baumann, G. Mutterkorn-farbstoffe, XIV. Isolierung, Struktur Und Absolute Konfiguration Der Ergochrome AD, BD, CD Und DD. Chem. Ber. 1966, 99, 3875–3883. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name/Chemical Class | Mol. Wt. | Mol. Formula | Host (Part, Family)/Location | Ref. |
|---|---|---|---|---|
| Isocoumarin derivatives | ||||
| 3R-Mellein (1) | 178 | C10H10O3 | Wet maize meal | [77,78] |
| - | - | Sea of Japan sediment | [20] | |
| - | - | Chondria crassicualis (Red alga, Rhodomelaceae), Yokji Island, Kyeongnam, Korea | [72] | |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [79] | |
| (3R,4S)-4-Hydroxymellein (2) | 194 | C10H10O4 | Cultured | [16,78] |
| - | - | Rhizospheric soil of Bruguiera gymnorrhiza (Marine mangrove plant, Rhizophoraceae), Hainan Island, China | [71] | |
| - | - | Pacific Ocean, China | [21] | |
| (3R,4R)-4-Hydroxymellein (3) | 194 | C10H10O4 | Rhizospheric soil of Bruguiera gymnorrhiza (Marine mangrove plant, Rhizophoraceae), Hainan Island, China | [71] |
| (R)-7-Hydroxymellein (4) | 194 | C10H10O4 | Rhizospheric soil of Bruguiera gymnorrhiza (Marine mangrove plant, Rhizophoraceae), Hainan Island, China | [71] |
| (R)-(–)-5-Bromomellein (5) | 255 | C10H9BrO3 | Chondria crassicualis (Red alga, Rhodomelaceae), Yokji Island, Kyeongnam, Korea | [72] |
| (±)-4,7-Dihydroxymellein (6) | 210 | C10H10O5 | Pacific Ocean, China | [21] |
| (3R,4S)-3,4-Dihydro-4,5,8-trihydroxy-3-methyl-1H-2-benzopyran-1-one (7) | 210 | C10H10O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| 6-Methoxy-8-hydroxyisocoumarin-3-carboxylic acid (8) | 236 | C11H8O6 | Cultured | [73] |
| (±)-Botryoisocoumarin A (9) | 208 | C11H12O4 | Rhizospheric soil of Bruguiera gymnorrhiza (Marine mangrove plant, Rhizophoraceae), Hainan Island, China | [71] |
| Diaporthin (10) | 250 | C13H14O5 | Cultured | [81] |
| Orthosporin (11) | 236 | C12H12O5 | Cultured | [81] |
| Ochratoxin α (12) | 256 | C11H9ClO5 | Cultured | [76] |
| Ochratoxin β (13) | 222 | C11H10O5 | Cultured | [76] |
| Ochratoxin A (14) | 403 | C20H18ClNO6 | Wet maize meal | [74,75,77] |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| 4-R-Hydroxyochratoxin A (15) | 419 | C20H18ClNO7 | Cultured | [76] |
| 4-S-Hydroxyochratoxin A (16) | 419 | C20H18ClNO7 | Cultured | [76] |
| 10-Hydroxyochratoxin A (17) | 419 | C20H18ClNO7 | Cultured | [76] |
| Ochratoxin B (18) | 369 | C20H19NO6 | Wet maize meal | [75,77] |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Bromo-ochratoxin B (19) | 447 | C20H18BrNO6 | Cultured | [82] |
| 4-R-Hydroxyochratoxin B (20) | 385 | C20H19NO7 | Cultured | [76] |
| Ochratoxin C (21) | 431 | C22H22ClNO6 | Wet maize meal | [75,77] |
| HO-Proline-ochratoxin (22) | 369 | C16H16ClNO7 | Cultured | [83] |
| Serine-ochratoxin (23) | 343 | C14H14ClNO7 | Cultured | [83] |
| Lysine-ochratoxin (24) | 384 | C17H21ClN2O6 | Cultured | [83] |
| Pyrazine derivatives | ||||
| Flavacol (25) | 208 | C12H20N2O | Moldy rice | [84,85] |
| - | - | Laboratory infected soil | [86] | |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] | |
| 6-(2-Hydroxy-2-methylpropyl)-3-isobutylpyrazin-2-ol (26) | 224 | C12H20N2O2 | Moldy rice | [84] |
| Neoaspergillc acid (27) | 224 | C12H20N2O2 | Moldy rice | [84,85] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] | |
| Neohydroxyaspergillc acid (28) | 240 | C12H20N2O3 | Moldy rice | [85] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] | |
| β-Hydroxyneoaspergillic acid (29) | 240 | C12H20N2O3 | Moldy rice | [85] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] | |
| Deoxy-β-hydroxyneoaspergillic acid (30) | 224 | C12H20N2O2 | Moldy rice | [84,85] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] | |
| Ochramide A (31) | 224 | C12H20N2O2 | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] |
| Ochramide B (32) | 238 | C13H22N2O2 | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] |
| Ochramide C (33) | 254 | C13H22N2O3 | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] |
| Ochramide D (34) | 238 | C12H18N2O3 | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22] |
| 3-Isobutyl-6-(1-hydroxy-2-methylpropyl)-2(1H)-pyrazinone (35) | 224 | C12H20N2O2 | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [22] |
| Ferrineoaspergillin (36) | 725 | C36H57FeN6O6 | Moldy rice | [84,85] |
| Ochralate A (37) | 786 | C39H63AlN6O9 | Dichotella gemmacea (Coral, Ellisellidae), South China Sea, Lingao, Hainan, China | [22,86] |
| Aluminiumneoaspergillin (38) | 744 | C36H57AlN6O9 | South China Sea Dichotella gemmacea (Ellisellidae), Lingao, Hainan, China | [22] |
| Diketopiperazines | ||||
| Waspergillamide B (39) | 450 | C20H26N4O8 | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Epiamauromine (40) | 508 | C32H36N4O2 | Cultured | [28] |
| N-Methylepiamauromine (41) | 522 | C33H38N4O2 | Cultured | [28] |
| Cycloechinulin (42) | 351 | C20H21N3O3 | Cultured | [28,87] |
| CJ-17,665 = Avrainvillamide (43) | 445 | C26H27N3O4 | Soil, Venezuela | [88] |
| Stephacidin A (44) | 431 | C26H29N3O3 | Light brown clay, Sirsaganj, Uttar Pradesh, India | [89] |
| - | - | Laboratory infected soil | [86] | |
| - | - | Soil, China | [90] | |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Stephacidin B (45) | 890 | C52H54N6O8 | Light brown clay, Sirsaganj, Uttar Pradesh, India | [89] |
| Speramide A (46) | 447 | C26H29N3O4 | Freshwater, Fuxian Lake, China | [23] |
| Speramide B (47) | 465 | C26H31N3O5 | Freshwater, Fuxian Lake, China | [23] |
| Asperochramide A (48) | 451 | C26H33N3O4 | Soil, China | [90] |
| Asperochramide B (49) | 451 | C26H33N3O4 | Soil, China | [90] |
| Asperochramide C (50) | 397 | C21H23N3O5 | Soil, China | [90] |
| Asperochramide D (51) | 315 | C16H17N3O4 | Soil, China | [90] |
| Notoamide A (52) | 463 | C26H29N3O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Notoamide B (53) | 447 | C26H29N3O4 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] |
| - | - | Soil, China | [90] | |
| Notoamide C (54) | 449 | C26H31N3O4 | Soil, China | [90] |
| Notoamide F (55) | 461 | C27H31N3O4 | Marine Culture, Pacific Ocean, China | [24] |
| Notoamide I (56) | 445 | C26H27N3O4 | Deep-sea water, Northeastern Pacific, China | [80] |
| Notoamide M (57) | 465 | C26H31N3O5 | Soil, China | [90] |
| - | - | Deep-sea water, Northeastern Pacific, China | [80] | |
| Notoamide X (58) | 461 | C26H27N3O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Sclerotiamide (59) | 463 | C26H29N3O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Taichunamide D (60) | 509 | C27H31N3O5S | Soil, China | [90] |
| Versicolamide B (61) | 447 | C26H29N3O4 | Marine Culture, Pacific Ocean, China | [24] |
| Brevianamide F (62) | 283 | C16H17N3O2 | Soil, China | [90] |
| Benzodiazepines | ||||
| (11aS)-2,3-Dihydro-7-methoxy-1H-pyrrolo [2,1-c][1,4] benzodiazepine-5,11(10H,11aH)-dione (63) | 246 | C13H14N2O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] |
| Circumdatin A (64) | 363 | C20H17N3O4 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [91] |
| - | - | Chondria crassicualis (Red alga, Rhodomelaceae), Yokji Island, Kyeongnam, Korea | [72] | |
| Circumdatin B (65) | 393 | C21H19N3O5 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [91] |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Circumdatin C (66) | 307 | C17H13N3O3 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [91] |
| - | - | Sea of Japan sediment | [20] | |
| - | - | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] | |
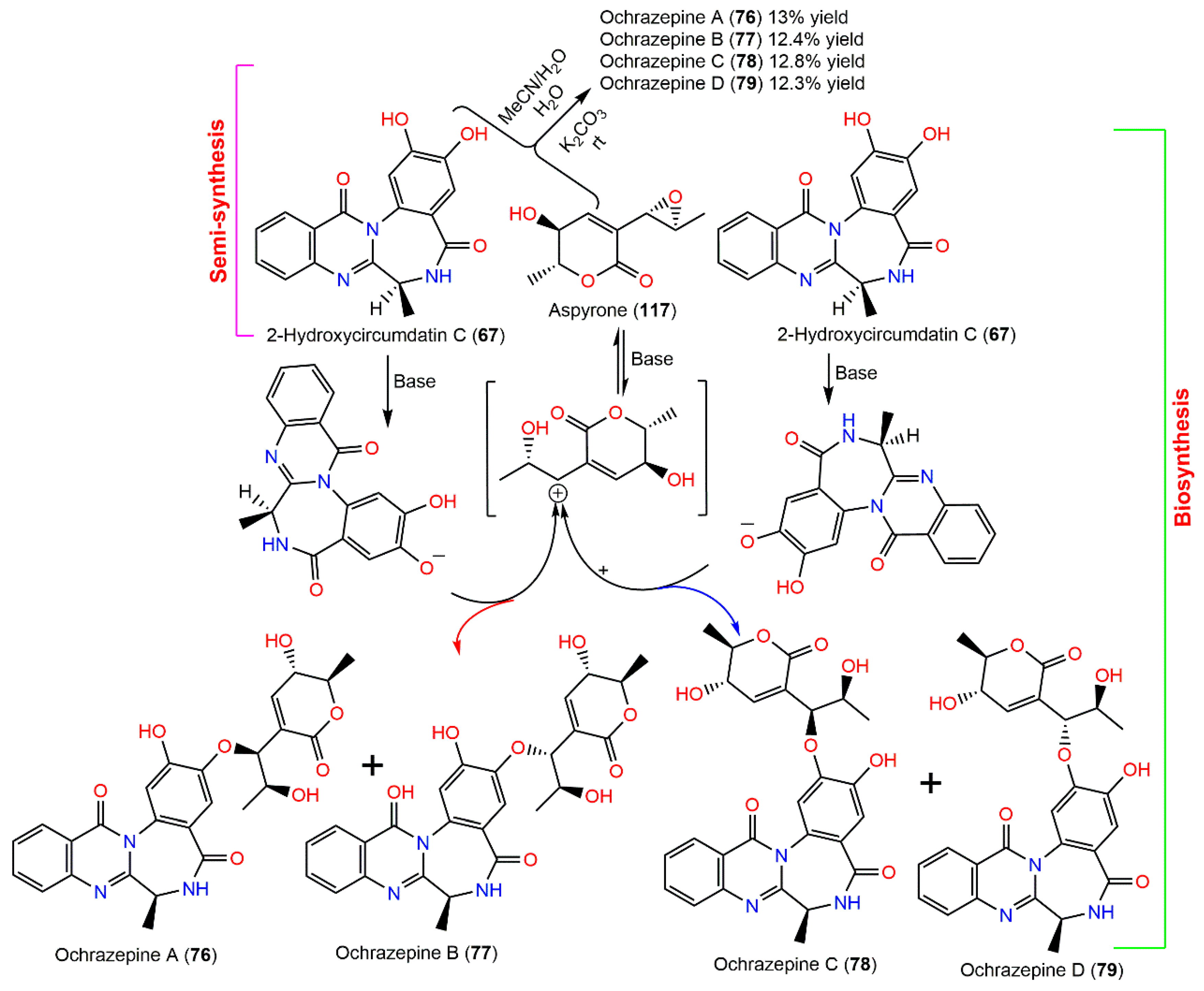
| 2-Hydroxycircumdatin C (67) | 323 | C17H13N3O4 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] | |
| Circumdatin D (68) | 393 | C21H19N3O5 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [92] |
| - | - | Laboratory infected soil | [86] | |
| - | - | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] | |
| Circumdatin E (69) | 363 | C20H17N3O4 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [92] |
| - | - | Laboratory infected soil | [86] | |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Circumdatin F (70) | 291 | C17H13N3O2 | Sterilized milo sorghum seeds buried in the soil for 1–4 months, Sevilleta National Wildlife Refuge, Socorro County, New Mexico, USA | [92] |
| - | - | Sea of Japan sediment | [20] | |
| - | - | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [26] | |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Circumdatin G (71) | 307 | C17H13N3O3 | Sea of Japan sediment | [20] |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Circumdatin H (72) | 347 | C20H17N3O3 | Laboratory infected soil | [86] |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Circumdatin I (73) | 323 | C17H13N3O4 | Cultured | [34] |
| Circumdatin L (74) | 307 | C17H13N3O3 | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Circumdatin N (75) | 307 | C17H13N3O3 | Marine Culture, Pacific Ocean, China | [24] |
| Ochrazepine A (76) | 507 | C26H25N3O8 | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] |
| Ochrazepine B (77) | 507 | C26H25N3O8 | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] |
| Ochrazepine C (78) | 507 | C26H25N3O8 | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] |
| Ochrazepine D (79) | 507 | C26H25N3O8 | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] |
| Indole and other alkaloids | ||||
| Ochrindole A (80) | 452 | C29H28N2O3 | Cultured | [29] |
| Ochrindole B (81) | 468 | C29H28N2O4 | Cultured | [29] |
| Ochrindole C (82) | 438 | C28H26N2O3 | Cultured | [29] |
| Ochrindole D (83) | 422 | C27H22N2O3 | Cultured | [29] |
| Perlolyrine (84) | 264 | C16H12N2O2 | Marine Culture, Pacific Ocean, China | [24] |
| Ochracesol A (85) | 205 | C11H11NO3 | Marine Culture, Pacific Ocean, China | [24] |
| L-657,398 (86) | 317 | C21H35NO | Cultured | [93] |
| Cycloanthranilylproline (87) | 230 | C13H14N2O2 | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Peptides | ||||
| Aspochracin (88) | 432 | C23H36N4O4 | Cultured | [94,95,96] |
| Violaceotide A (89) | 476 | C24H36N4O6 | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Sesquiterpenoids | ||||
| 6β,9α-Dihydroxy-14-p-nitrobenzoylcinnamolide (90) | 431 | C22H25NO8 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [32,97] |
| Insulicolide A (91) | 431 | C22H25NO8 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [32] |
| 14-O-Acetylinsulicolide A (92) | 475 | C23H25NO10 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [97] |
| 9-Deoxyinsulicolide A (93) | 415 | C22H25NO7 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [97] |
| Insulicolide B (94) | 415 | C22H25NO7 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [97] |
| Insulicolide C (95) | 459 | C23H25NO9 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [97] |
| 6β,14-Dihydroxy-7α-methoxyconfertifolin (96) | 296 | C16H24O5 | Coelarthrum sp. (Green alga, Rhodymeniaceae), South China Sea, China | [97] |
| Polyketides | ||||
| Penicillic acid (97) | 170 | C8H10O4 | Cultured | [30,98] |
| - | - | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] | |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| 5(6)-Dihydropenicillic acid (98) | 172 | C8H12O4 | Cultured | [98] |
| - | - | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] | |
| - | - | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Asperlactone (99) | 184 | C9H12O4 | Environmental contamination sample, apple-packing house, Lleida, Spain | [99] |
| Chlorohydroasperlactone A (100) | 220 | C9H13ClO4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| Chlorohydroasperlactone B (101) | 220 | C9H13ClO4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [79] | |
| Aspilactonol B (102) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Aspilactonol E (103) | 186 | C9H14O4 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochrin A (104) | 202 | C9H14O5 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant, Rhizophoraceae), Hainan Island, China | [71] |
| Asperochrin B (105) | 184 | C9H12O4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant, Rhizophoraceae), Hainan Island, China | [71] |
| Asperochrin C (106) | 184 | C9H12O4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| Asperochratide A (107) | 216 | C10H16O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide B (108) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide C (109) | 216 | C10H16O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide D (110) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide E(111) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide F (112) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide G (113) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide H (114) | 188 | C9H16O4 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide I (115) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Asperochratide J (116) | 216 | C10H16O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Aspyrone (117) | 184 | C9H12O4 | Cultured | [33,100,101,102] |
| - | - | Environmental contamination sample, apple-packing house, Lleida, Spain | [99] | |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), Lingao, Hainan, China | [31] | |
| Chlorohydroaspyrone A (118) | 220 | C9H13ClO4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| Chlorohydroaspyrone B (119) | 220 | C9H13ClO4 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [79] | |
| Aspyronol (120) | 216 | C10H16O5 | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] |
| Dihydroaspyrone (121) | 186 | C9H14O4 | Cultured | [33] |
| - | - | Rhizospheric soil of Bruguiera gymnorrhiza (Mangrove plant Rhizophoraceae), Hainan Island, China | [71] | |
| - | - | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [79] | |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| 8,9-Dihydroxy-8,9-deoxyaspyrone (122) | 202 | C9H14O5 | Deep-sea water, Northeastern Pacific, China | [80] |
| Ochraspergillic acid A (123) | 307 | C15H17NO6 | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Ochraspergillic acid B (124) | 307 | C15H17NO6 | Agelas oroides (Sponge Agelasidae), Sığaçık-Izmir, Turkey | [17] |
| Aspinolide A (125) | 184 | C10H16O3 | Cultured | [33] |
| Aspinolide B (126) | 284 | C14H20O6 | Cultured | [33] |
| Aspinolide C (127) | 282 | C14H18O6 | Cultured | [33] |
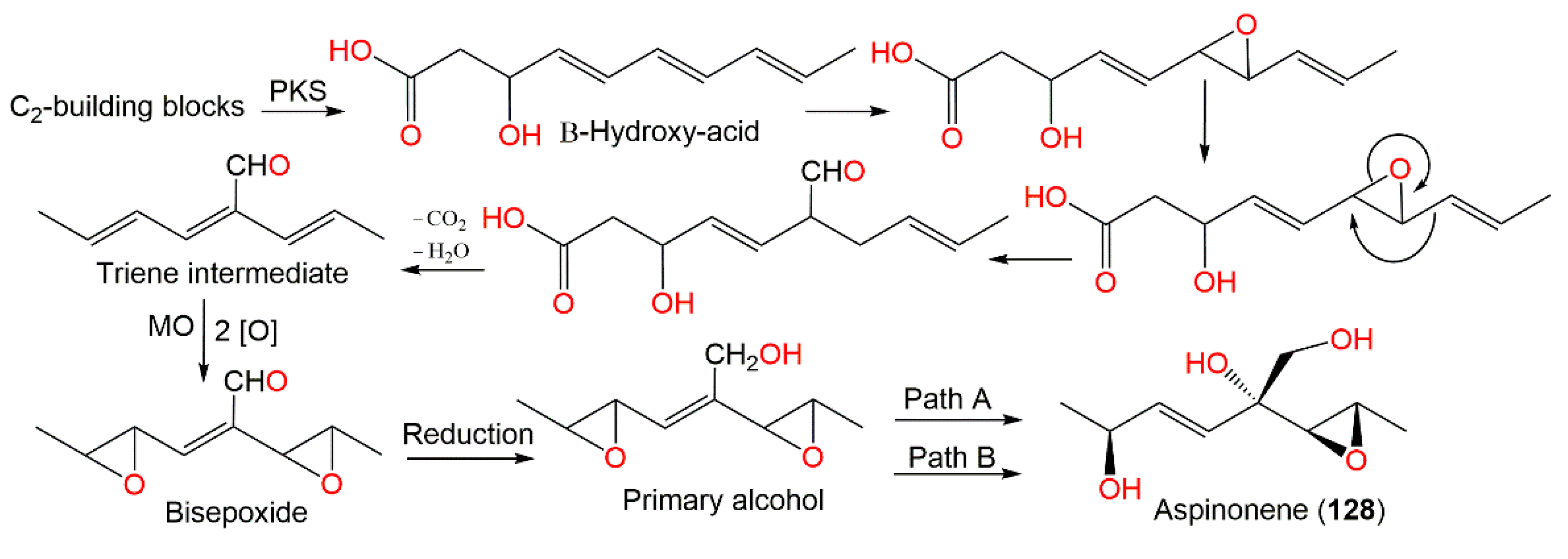
| Aspinonene (128) | 188 | C9H16O4 | Cultured | [33,100] |
| Iso-aspinonene (129) | 188 | C9H16O4 | Cultured | [33] |
| Quinone Derivatives | ||||
| Emodin (130) | 270 | C15H10O5 | Moldy rice | [103] |
| Solanum tuberosum (Chinese potato, Solanaceae) | [104] | |||
| Xanthomegnin (131) | 574 | C30H22O12 | Cultured | [105,106] |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Viomellein (132) | 560 | C30H24O11 | Cultured | [105,106] |
| - | - | Agelas oroides (Sponge, Agelasidae), Sığaçık-Izmir, Turkey | [17] | |
| Xanthine Derivatives | ||||
| Vioxanthin (133) | 546 | C30H26O10 | Green coffee beans | [107] |
| R-(+)-Semi-vioxanthin (134) | 274 | C15H14O5 | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [79] |
| Secalonic acid (135) | 638 | C32H30O14 | Moldy rice | [103] |
| Flavonoids | ||||
| Lucenin-2 (136) | 610 | C27H30O16 | Marine sponge, Kanyakumari, peninsular land, southeast coast of Tamilnadu state, India | [108] |
| Sterols | ||||
| 7-Nor-ergosterolide (137) | 414 | C27H42O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| 3β,11α-Dihydroxyergosta-8,24(28)-dien-7-one (138) | 428 | C28H44O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| 3β-Hydroxyergosta-8,24(28)-dien-7-one (139) | 412 | C28H44O2 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-3β,5α,9α-Trihydroxyergosta-7,22-dien-6-one (140) | 444 | C28H44O4 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China (Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-3β,5α -Dihydroxyergosta-7,22-dien-6-one (141) | 428 | C28H44O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| Ergosterol (142) | 396 | C28H44O | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-Ergosta-4,6,8(14),22-tetraen-3-one (143) | 392 | C28H40O | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-Ergosta-7,22-diene-3β,5α,6β-triol (144) | 430 | C28H46O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-Ergosta-7,22-diene-6β-methoxy-3β,5α-diol (145) | 444 | C29H48O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-ergosta-7,22-diene-3β,6β-diol (146) | 414 | C28H46O2 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-Ergosta-5α,6α-epoxide-8,22-diene-3β,7α-diol (147) | 428 | C28H44O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-5α,8α-Epidioxyergosta-6,22-dien-3β-ol (148) | 428 | C28H44O3 | Sargassum kjellmanianum (Brown alga, Sargassaceae), Dalian coastline, China | [27] |
| (22E,24R)-Ergosta-7,22-diene-3β,5α,6β,9α-tetraol (149) | 446 | C28H46O4 | Deep-sea water, Northeastern Pacific, China | [80] |
| Ochrasterone (150) | 456 | C30H48O3 | Pacific Ocean, China | [21] |
| Gymnasterone D (151) | 406 | C28H38O2 | Marine Culture, Pacific Ocean, China | [24] |
| Isocyathisterol (152) | 410 | C28H42O2 | Marine Culture, Pacific Ocean, China | [24] |
| Herbarulide (153) | 424 | C28H40O3 | Marine Culture, Pacific Ocean, China | [24] |
| Demethylincisterol A2 (154) | 332 | C21H32O3 | Marine Culture, Pacific Ocean, China | [24] |
| Other Metabolites | ||||
| 4-(3-Methyl-2- butenyl) oxy-1-phenyl acetic acid (155) | 220 | C13H16O3 | Cultured | [109] |
| Clavatol (156) | 180 | C10H12O3 | Chondria crassicualis (Korean name: Seosil) (Red alga, Rhodomelaceae), Yokji Island, Kyeongnam, Korea | [72] |
| 2,2-[bis-4-(2,3-Dihydroxypropoxy) phenyl]propane (157) | 376 | C21H28O6 | Deep-sea water of the Northeastern Pacific, China | [80] |
| 6-Ethyloct-3-yl-2- ethylhexyl ester (158) | 418 | C26H42O4 | Marine sponge, Kanyakumari, peninsular land, southeast coast of Tamilnadu state, India | [108] |
| Di-(2-Ethylhexyl) phthalate (159) | 390 | C24H38O4 | Marine Culture, Pacific Ocean, China | [24] |
| Campholene aldehyde (160) | 152 | C10H16O | Marine sponge, Kanyakumari, peninsular, southeast coast of Tamilnadu state, India | [108] |
| Dientriol (161) | 172 | C9H16O3 | Cultured | [33] |
| Triendiol (162) | 154 | C9H14O2 | Cultured | [33] |
| 2,10-Dimethyl 4-hydroxy-6-oxo-4-undecen-7-yne (163) | 208 | C13H20O2 | Cultured | [109] |
| Eythrol (164) | 122 | C4H10O4 | Deep-sea water, Northeastern Pacific, China | [80] |
| Galactomannan (165) | 828 | C30H52O26 | Dichotella gemmacea (Coral, Ellisellidae), South Sea, China | [35] |
| Compound Name | Biological Activity | Assay, Organism or Cell Line | Biological Results | Ref. | |
|---|---|---|---|---|---|
| Compound | Positive Control | ||||
| 3R-Mellein (1) | Anti HCV protease | SPA | 35.0 µM (IC50) | - | [20] |
| Antioxidant | Spectrophotometry/DPPH | 25.0 µM (IC50) | l-ascorbic acid 20.0 μM (IC50) | [72] | |
| (3R,4S)-4-Hydroxymellein (2) | Antioxidant | Spectrophotometry/DPPH | 129.36 µM (IC50) | BHT 91.35 µM (IC50) | [21] |
| Spectrophotometry/ABTS | 140.23 µM (IC50) | Trolox 101.23 µM (IC50) | [21] | ||
| Spectrophotometry/FRAP | 11.96 µM (IC50) | Trolox 1.80 µM (IC50) | [21] | ||
| (R)-7-Hydroxymellein (4) | Antibacterial | Well diffusion/A. hydrophilia | 64.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 32.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 8.0 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| (R)-(–)-5-Bromomellein (5) | Antioxidant | Spectrophotometry/DPPH | 24.0 µM (IC50) | l-ascorbic acid 20.0 μM (IC50) | [72] |
| (±)-4,7-dihydroxymellein (6) | Antioxidant | Spectrophotometry/DPPH | 62.90 µM (IC50) | BHT 91.35 µM (IC50) | [21] |
| Spectrophotometry/ABTS | 70.92 µM (IC50) | Trolox 101.23 µM (IC50) | [21] | ||
| Spectrophotometry/FRAP | 2.53 µM (IC50) | Trolox 1.80 µM (IC50) | [21] | ||
| Ochratoxin B (18) | Cytotoxicity | MTT/A2780 | 3.0 µM (IC50) | Cisplatin 2.2 µM (IC50) | [17] |
| Flavacol (25) | Inhibition of mitochondrial NADH oxidase | SMP/NADH oxidase | 34.6 µM (IC50) | - | [86] |
| CJ-17,665 = Avrainvillamide (43) | Antibacterial | Microdilution/S. aureus | 12.5 μg/mL (MIC) | Vancomycin 1.56 μg/mL (MIC) | [88] |
| Microdilution/S. pyogenes | 12.5 μg/mL (MIC) | Vancomycin 0.39 μg/mL (MIC) | [88] | ||
| Microdilution/E. pyogenes | 25.0 μg/mL (MIC) | Vancomycin 12.5 μg/mL (MIC) | [88] | ||
| Cytotoxicity | MTT/HeLa | 1.1 μg/mL (IC90) | - | [88] | |
| Stephacidin A (44) | Cytotoxicity | MTT/PC-3 | 2.1 µM (IC50) | - | [89] |
| MTT/LNCaP | 1.0 µM (IC50) | - | [89] | ||
| MTT/A2780 | 4.0 µM (IC50) | - | [89] | ||
| MTT/A2780/DDP | 6.8 µM (IC50) | - | [89] | ||
| MTT/A2780/Tax | 3.6 µM (IC50) | - | [89] | ||
| MTT/HCT-116 | 2.1 µM (IC50) | - | [89] | ||
| MTT/HCT116/mdr+ | 6.7 µM (IC50) | - | [89] | ||
| MTT/HCT116/topo | 13.1 µM (IC50) | - | [89] | ||
| MTT/MCF-7 | 4.2 µM (IC50) | - | [89] | ||
| MTT/SKBR3 | 2.15 µM (IC50) | - | [89] | ||
| MTT/LX-1 | 4.22 µM (IC50) | - | [89] | ||
| Inhibition of mitochondrial NADH Oxidase | SMP/NADH oxidase | 13.0 µM (IC50) | - | [86] | |
| Anti-Parkinson’s disease | MPP+-induced SH-SY5Y cells | 2.45 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] | |
| Stephacidin B (45) | Cytotoxicity | MTT/PC-3 | 0.37 µM (IC50) | - | [89] |
| MTT/LNCaP | 0.06 µM (IC50) | - | [89] | ||
| MTT/A2780 | 0.33 µM (IC50) | - | [89] | ||
| MTT/A2780/DDP | 0.43 µM (IC50) | - | [89] | ||
| MTT/A2780/Tax | 0.26 µM (IC50) | - | [89] | ||
| MTT/HCT-116 | 0.46 µM (IC50) | - | [89] | ||
| MTT/HCT116/mdr+ | 0.46 µM (IC50) | - | [89] | ||
| MTT/HCT116/topo | 0.42 µM (IC50) | - | [89] | ||
| MTT/MCF-7 | 0.27 µM (IC50) | - | [89] | ||
| MTT/SKBR3 | 0.32 µM (IC50) | - | [89] | ||
| MTT/LX-1 | 0.38 µM (IC50) | - | [89] | ||
| Speramide A (46) | Antibacterial | 2-Fold dilution/ P. aeruginosa | 0.8 µM (MIC) | - | [23] |
| Notoamide B (53) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 5.31 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Notoamide C (54) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 7.39 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Notoamide F (55) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 2.98 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Notoamide I (56) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 2.30 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Versicolamide B (61) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 6.01 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Circumdatin A (64) | Antioxidant | Spectrophotometry/DPPH | 32.0 µM (IC50) | l-ascorbic acid 20.0 μM (IC50) | [72] |
| 2-Hydroxycircumdatin C (67) | Antioxidant | Spectrophotometry/DPPH | 9.9 µM (IC50) | BHT 88.2 µM (IC50) | [26] |
| Cytotoxicity | CTG/U251 | 8.95 µM (IC50) | Adramycin 0.19 µM (IC50) | [31] | |
| Circumdatin E (69) | Inhibition of Mitochondrial NADH Oxidase | SMP/NADH oxidase | 2.5 µM (IC50) | - | [86] |
| Circumdatin F (70) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 5.44 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Circumdatin G (71) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 7.39 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Circumdatin H (72) | Inhibition of Mitochondrial NADH Oxidase | SMP/NADH oxidase | 1.5 µM (IC50) | - | [86] |
| Circumdatin N (75) | Anti-Parkinson’s disease | MPP+-induced SH-SY5Y cells | 10.77 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Ochrazepine A (76) | Cytotoxicity | CTG/MV-4-11 | 3.94 µM (IC50) | Adramycin 0.16 µM (IC50) | [31] |
| CTG/K562 | 6.05 µM (IC50) | Adramycin 0.02 µM (IC50) | [31] | ||
| CTG/A673 | 3.10 µM (IC50) | Adramycin 0.13 µM (IC50) | [31] | ||
| CTG/U87 | 8.67 µM (IC50) | Adramycin 0.12 µM (IC50) | [31] | ||
| CTG/A549 | 9.62 µM (IC50) | Adramycin 0.10 µM (IC50) | [31] | ||
| CTG/N87 | 6.10 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/H1299 | 7.14 µM (IC50) | Adramycin 0.49 µM (IC50) | [31] | ||
| CTG/HUCCT1 | 11.32 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/B16F10 | 11.22 µM (IC50) | Adramycin 0.02 µM (IC50) | [31] | ||
| CTG/Karpass299 | 5.89 µM (IC50) | Adramycin 0.39 µM (IC50) | [31] | ||
| CTG/HEK-293F | 12.91 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/L02 | 27.42 µM (IC50) | Adramycin 0.10 µM (IC50) | [31] | ||
| Ochrazepine B (77) | Cytotoxicity | CTG/U251 | 9.91 µM (IC50) | Adramycin 0.19 µM (IC50) | [31] |
| CTG/HEK-293F | 73.96 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| Ochrazepine C (78) | Cytotoxicity | CTG/A673 | 8.24 µM (IC50) | Adramycin 0.13 µM (IC50) | [31] |
| CTG/U87 | 9.04 µM (IC50) | Adramycin 0.12 µM (IC50) | [31] | ||
| CTG/HepB3 | 10.28 µM (IC50) | Adramycin 17.58 µM (IC50) | [31] | ||
| CTG/HEK-293F | 73.03 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| Ochrazepine D (79) | Cytotoxicity | CTG/U251 | 8.26 µM (IC50) | Adramycin 0.19 µM (IC50) | [31] |
| CTG/HEK-293F | 54.58 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| Perlolyrine (84) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 9.97 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Ochracesol A (85) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 17.84 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| 6β,9α-Dihydroxy-14-p-nitrobenzoylcinnamolide (90) | Cytotoxicity | CCK-8/H1975 | 2.08 µM (IC50) | Trichostatin A 0.09 µM (IC50) | [32] |
| CCK-8/U937 | 1.95 µM (IC50) | Trichostatin A 0.05 µM (IC50) | [32] | ||
| CCK-8/K562 | 4.33 µM (IC50) | Trichostatin A 0.16 µM (IC50) | [32] | ||
| CCK-8/BGC823 | 2.32 µM (IC50) | Trichostatin A 0.08 µM (IC50) | [32] | ||
| CCK-8/Molt-4 | 2.39 µM (IC50) | Trichostatin A 0.03 µM (IC50) | [32] | ||
| CCK-8/MCF-7 | 4.25 µM (IC50) | Trichostatin A 0.06 µM (IC50) | [32] | ||
| CCK-8/A549 | 2.41 µM (IC50) | Trichostatin A 0.05 µM (IC50) | [32] | ||
| CCK-8/HeLa | 6.12 µM (IC50) | Trichostatin A 0.10 µM (IC50) | [32] | ||
| CCK-8/HL-60 | 2.44 µM (IC50) | Trichostatin A 0.03 µM (IC50) | [32] | ||
| CCK-8/Huh-7 | 3.28 µM (IC50) | Trichostatin A 0.09 µM (IC50) | [32] | ||
| CCK-8/ACHN | 11.0 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] | ||
| CCK-8/OS-RC-2 | 8.2 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 4.3 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| Antiviral | CCK-8/H3N2 | 17.0 µM (IC50) | Oseltamivir A 0.008 µM (IC50) | [32] | |
| CCK-8/EV71 | 9.4 µM (IC50) | Oseltamivir A 0.06 µM (IC50) | [32] | ||
| Insulicolide A (91) | Cytotoxicity | CCK-8/H1975 | 4.63 µM (IC50) | Trichostatin A 0.09 µM (IC50) | [32] |
| CCK-8/U937 | 3.97 µM (IC50) | Trichostatin A 0.05 µM (IC50) | [32] | ||
| CCK-8/K562 | 4.76 µM (IC50) | Trichostatin A 0.16 µM (IC50) | [32] | ||
| CCK-8/BGC823 | 2.78 µM (IC50) | Trichostatin A 0.08 µM (IC50) | [32] | ||
| CCK-8/Molt-4 | 2.11 µM (IC50) | Trichostatin A 0.03 µM (IC50) | [32] | ||
| CCK-8/MCF-7 | 6.08 µM (IC50) | Trichostatin A 0.06 µM (IC50) | [32] | ||
| CCK-8/A549 | 2.86 µM (IC50) | Trichostatin A 0.05 µM (IC50) | [32] | ||
| CCK-8/HeLa | 6.35 µM (IC50) | Trichostatin A 0.10 µM (IC50) | [32] | ||
| CCK-8/HL-60 | 2.34 µM (IC50) | Trichostatin A 0.03 µM (IC50) | [32] | ||
| CCK-8/Huh-7 | 2.35 µM (IC50) | Trichostatin A 0.09 µM (IC50) | [32] | ||
| CCK-8/ACHN | 1.5 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] | ||
| CCK-8/OS-RC-2 | 1.5 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 0.89 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| 14-O-Acetylinsulicolide A (92) | Cytotoxicity | CCK-8/ACHN | 4.1 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] |
| CCK-8/OS-RC-2 | 5.3 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 2.3 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| 9-Deoxyinsulicolide A (93) | Cytotoxicity | CCK-8/ACHN | 25.0 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] |
| CCK-8/OS-RC-2 | 30.0 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 20.0 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| Insulicolide B (94) | Cytotoxicity | CCK-8/ACHN | 30.0 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] |
| CCK-8/OS-RC-2 | 23.0 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 24.0 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| Insulicolide C (95) | Cytotoxicity | CCK-8/ACHN | 13.0 µM (IC50) | Sorafenib 3.4 µM (IC50) | [97] |
| CCK-8/OS-RC-2 | 11.0 µM (IC50) | Sorafenib 7.0 µM (IC50) | [97] | ||
| CCK-8/786-O | 14.0 µM (IC50) | Sorafenib 4.9 µM (IC50) | [97] | ||
| Penicillic acid (97) | Antibacterial | Well diffusion/A. hydrophilia | 1.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 32.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 0.5 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| Chlorohydroasperlactone A (100) | Antibacterial | Well diffusion/A. hydrophilia | 16.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 16.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 16.0 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| Asperochrin A (104) | Antibacterial | Well diffusion/A. hydrophilia | 8.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 16.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 8.0 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| Aspyrone (117) | Cytotoxicity | CTG/MV-4-11 | 2.54 µM (IC50) | Adramycin 0.16 µM (IC50) | [31] |
| CTG/K562 | 5.22 µM (IC50) | Adramycin 0.02 µM (IC50) | [31] | ||
| CTG/A673 | 8.55 µM (IC50) | Adramycin 0.13 µM (IC50) | [31] | ||
| CTG/N87 | 4.57 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/H1299 | 5.83 µM (IC50) | Adramycin 0.49 µM (IC50) | [31] | ||
| CTG/HUCCT1 | 9.79 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/B16F10 | 5.89 µM (IC50) | Adramycin 0.02 µM (IC50) | [31] | ||
| CTG/Karpass299 | 2.57 µM (IC50) | Adramycin 0.39 µM (IC50) | [31] | ||
| CTG/HepB3 | 5.48 µM (IC50) | Adramycin 17.58 µM (IC50) | [31] | ||
| CTG/HA431 | 5.92 µM (IC50) | Adramycin 0.17 µM (IC50) | [31] | ||
| CTG/143B | 6.32 µM (IC50) | Adramycin 0.10 µM (IC50) | [31] | ||
| CTG/MKN-45 | 5.79 µM (IC50) | Adramycin 0.20 µM (IC50) | [31] | ||
| CTG/H1975 | 2.99 µM (IC50) | Adramycin 0.09 µM (IC50) | [31] | ||
| CTG/Hl-60 | 6.89 µM (IC50) | Adramycin 0.21 µM (IC50) | [31] | ||
| CTG/DU145 | 5.61 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/SPC-A1 | 9.51 µM (IC50) | Adramycin 0.19 µM (IC50) | [31] | ||
| CTG/HEK-293F | 50.35 µM (IC50) | Adramycin 0.05 µM (IC50) | [31] | ||
| CTG/L02 | 14.20 µM (IC50) | Adramycin 0.10 µM (IC50) | [31] | ||
| Chlorohydroaspyrone A (118) | Antibacterial | Well diffusion/A. hydrophilia | 16.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 32.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 16.0 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| Chlorohydroaspyrone B (119) | Antibacterial | Well diffusion/A. hydrophilia | 16.0 μg/mL (MIC) | Chloramphenicol 4.0 μg/mL (MIC) | [71] |
| Well diffusion/V. anguillarum | 32.0 μg/mL (MIC) | Chloramphenicol 1.0 μg/mL (MIC) | [71] | ||
| Well diffusion/V. harveyi | 32.0 μg/mL (MIC) | Chloramphenicol 8.0 μg/mL (MIC) | [71] | ||
| Viomellein (132) | Cytotoxicity | MTT/L5178Y | 5.3 µM (IC50) | Kahalalide F 4.3 µM (IC50) | [17] |
| MTT/A2780 | 5.0 µM (IC50) | Cisplatin 2.2 µM (IC50) | [17] | ||
| (22E,24R)-ergosta-4,6,8(14),22-tetraen-3-one (143) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 35.71 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Ochrasterone (150) | Antioxidant | DPPH | 189.10 µM (IC50) | BHT 91.35 µM (IC50) | [21] |
| ABTS | 60.21 µM (IC50) | Trolox 101.23 µM (IC50) | [21] | ||
| FRAP | 57.20 µM (IC50) | Trolox 1.80 µM (IC50) | [21] | ||
| Gymnasterone D (151) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 38.75 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Isocyathisterol (152) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 46.18 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Herbarulide (153) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 37.67 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Demethylincisterol A2 (154) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 49.53 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
| Clavatol (156) | Antioxidant | DPPH | 30.0 µM (IC50) | l-ascorbic acid 20.0 μM (IC50) | [72] |
| Di-(2-ethylhexyl) phthalate (159) | Anti-Parkinson’s disease | MPP+--induced SH-SY5Y cells | 80.21 µM (EC50) | Levodopa 2.06 µM (EC50) | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hareeri, R.H.; Aldurdunji, M.M.; Abdallah, H.M.; Alqarni, A.A.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Aspergillus ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential. Molecules 2022, 27, 6759. https://doi.org/10.3390/molecules27196759
Hareeri RH, Aldurdunji MM, Abdallah HM, Alqarni AA, Mohamed SGA, Mohamed GA, Ibrahim SRM. Aspergillus ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential. Molecules. 2022; 27(19):6759. https://doi.org/10.3390/molecules27196759
Chicago/Turabian StyleHareeri, Rawan H., Mohammed M. Aldurdunji, Hossam M. Abdallah, Ali A. Alqarni, Shaimaa G. A. Mohamed, Gamal A. Mohamed, and Sabrin R. M. Ibrahim. 2022. "Aspergillus ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential" Molecules 27, no. 19: 6759. https://doi.org/10.3390/molecules27196759
APA StyleHareeri, R. H., Aldurdunji, M. M., Abdallah, H. M., Alqarni, A. A., Mohamed, S. G. A., Mohamed, G. A., & Ibrahim, S. R. M. (2022). Aspergillus ochraceus: Metabolites, Bioactivities, Biosynthesis, and Biotechnological Potential. Molecules, 27(19), 6759. https://doi.org/10.3390/molecules27196759