



The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase



Abstract
:1. Introduction
2. Results
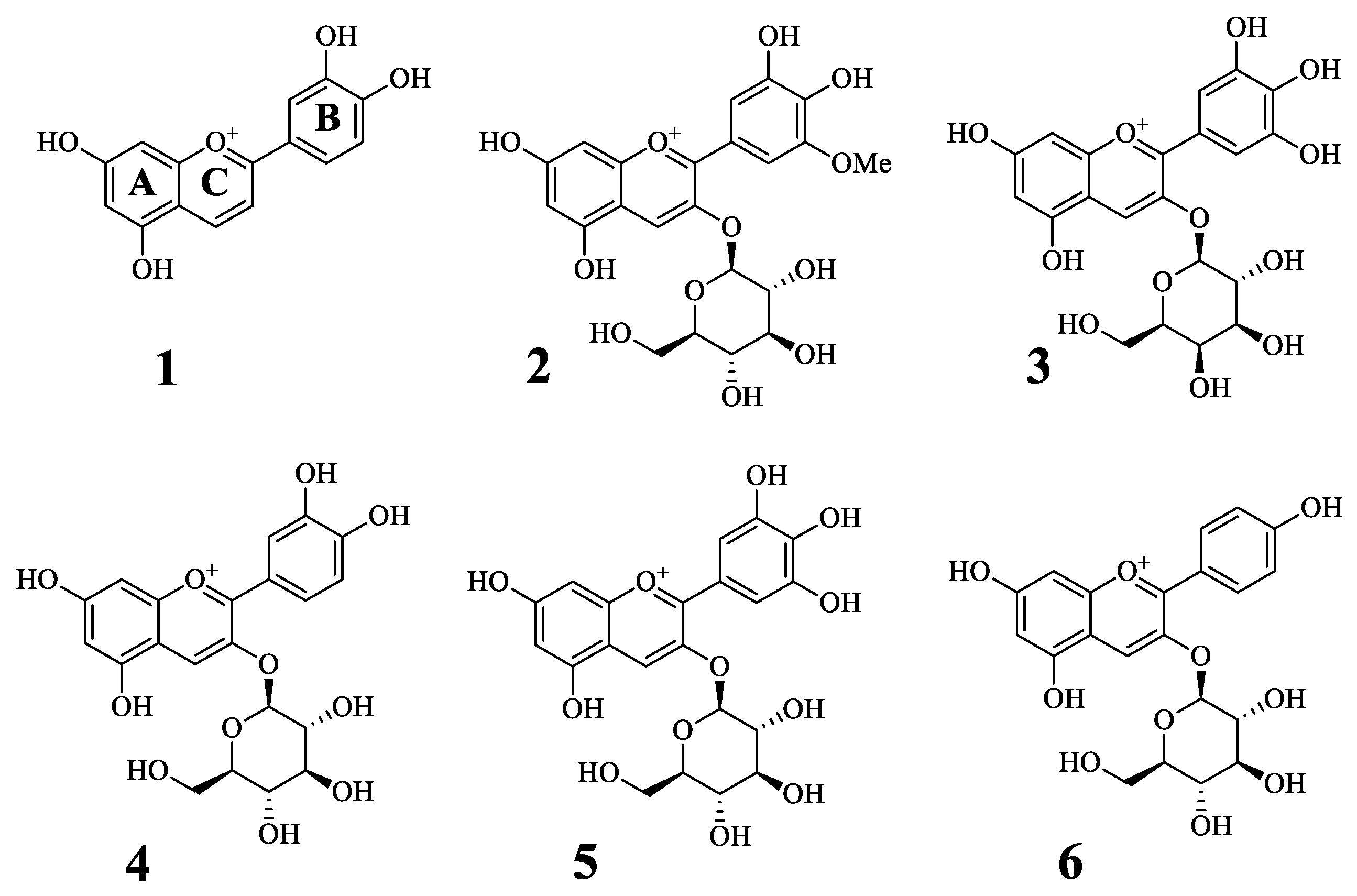
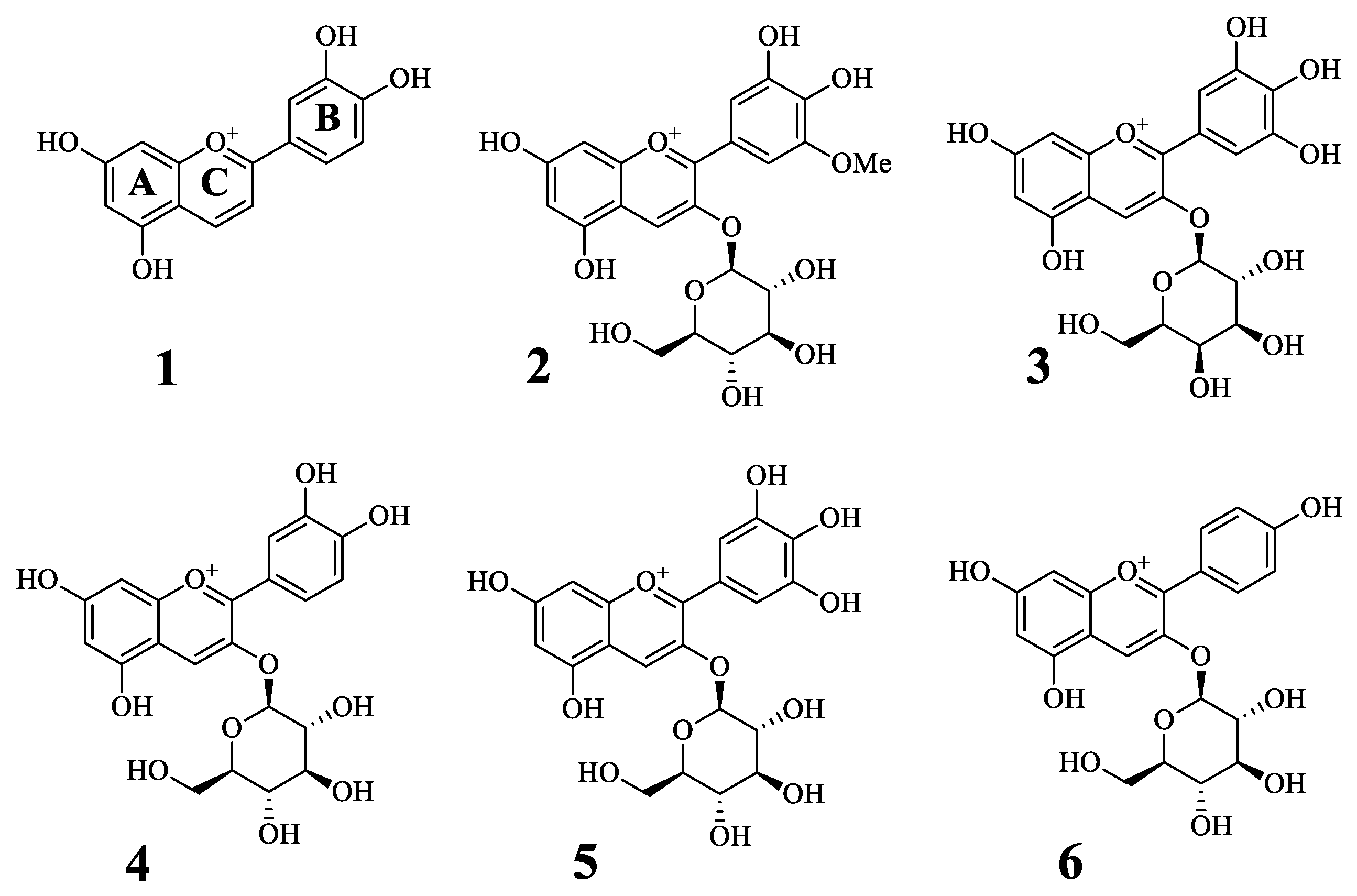
2.1. Inhibitory Activity of Antocyanins on Tyrosinase
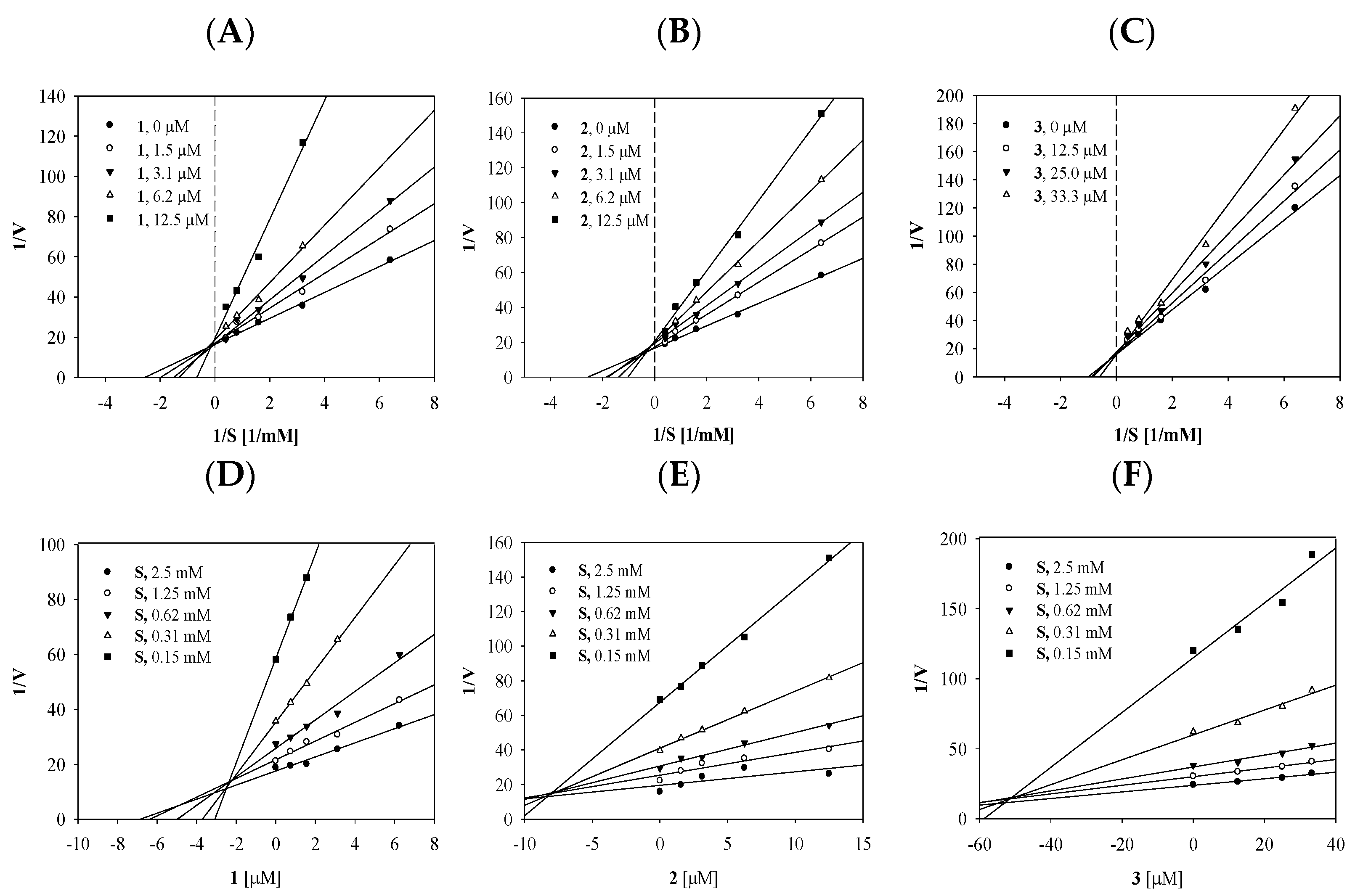
2.2. Enzyme Kinetics of the Compounds on Tyrosinase
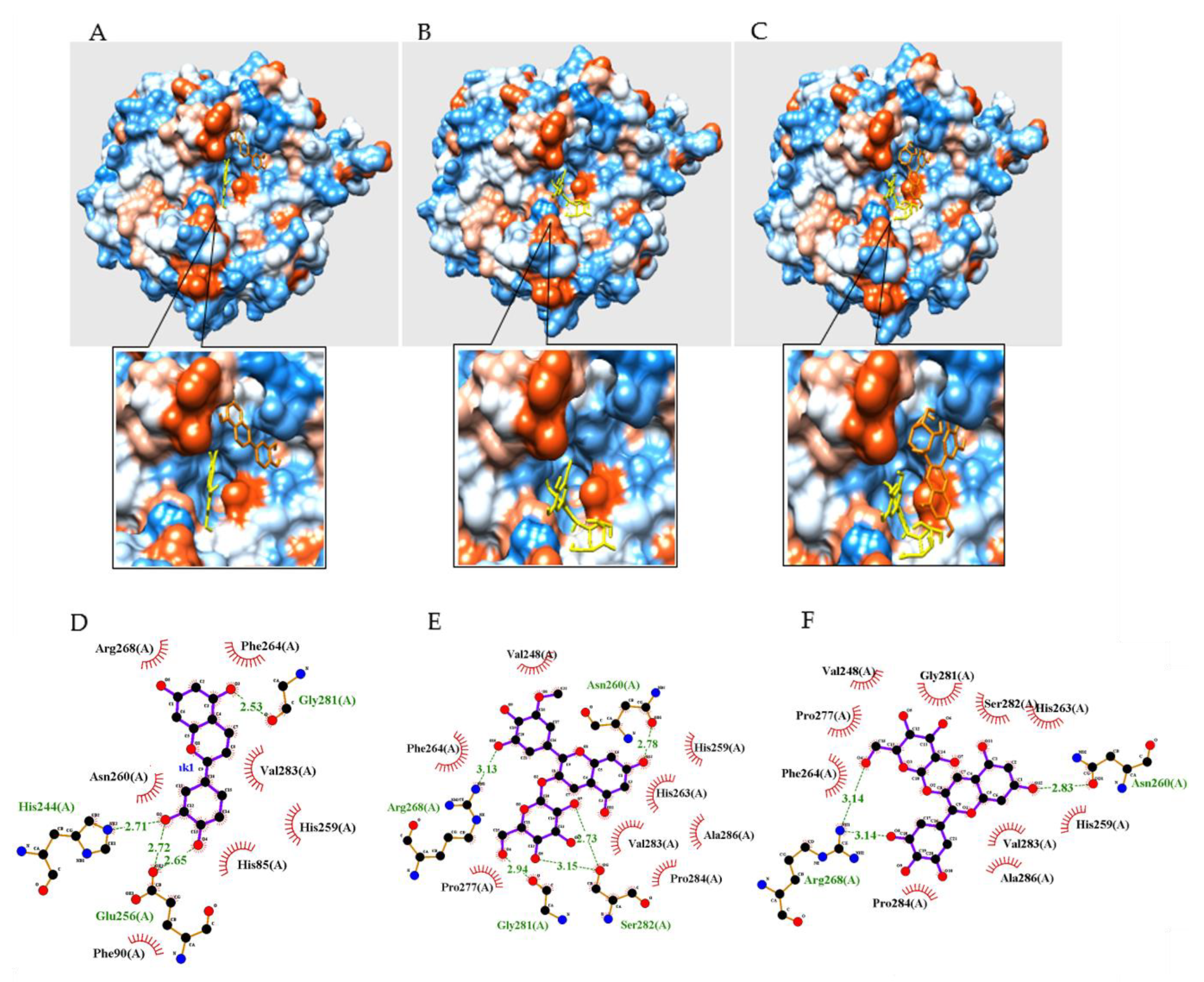
2.3. Molecular Docking of the Compounds with Tyrosinase
3. Experimental Section
3.1. Chemical Reagents
3.2. Tyrosinase Assay
3.3. Molecular Docking
3.4. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Chen, J.; Li, Q.; Ye, Y.; Huang, Z.; Ruan, Z.; Jin, N. Phloretin as both a substrate and inhibitor of tyrosinase: Inhibitory activity and mechanism. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 226, 117642. [Google Scholar] [CrossRef] [PubMed]
- Bahuguna, A.; Bharadwaj, S.; Chauhan, A.K.; Kang, S.C. Inhibitory insights of strawberry (Fragaria × ananassa var. Seolhyang) root extract on tyrosinase activity using computational and in vitro analysis. Int. J. Biol. Macromol. 2020, 165, 2773–2788. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Xie, L.; Lin, Q.; Tong, C.; Fu, Q.; Xu, J.; Xiao, J.; Shi, S. Profiling of tyrosinase inhibitors in mango leaves for a sustainable agro-industry. Food Chem. 2020, 312, 126042. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.R.; Menezes, T.M.; da Silva, L.P.; Pires, D.S.; Princival, J.L.; Seabra, G.; Neves, J.L. Furan inhibitory activity against tyrosinase and impact on B16F10 cell toxicity. Int. J. Biol. Macromol. 2019, 136, 1034–1041. [Google Scholar] [CrossRef]
- Shu, P.; Fei, Y.; Li, J.; Liu, A.; Zhang, L.; Niu, H.; Liu, W.; Wei, X.; Xiao, F.; Xu, Z. Two new phenylethanoid glycosides from Ginkgo biloba leaves and their tyrosinase inhibitory activities. Carbohydr. Res. 2020, 494, 108059. [Google Scholar] [CrossRef]
- Tian, J.-L.; Liu, T.-L.; Xue, J.-J.; Hong, W.; Zhang, Y.; Zhang, D.-X.; Cui, C.-C.; Liu, M.-C.; Niu, S.-L. Flavanoids derivatives from the root bark of Broussonetia papyrifera as a tyrosinase inhibitor. Ind. Crop. Prod. 2019, 138, 111445. [Google Scholar] [CrossRef]
- Kang, S.-M.; Heo, S.-J.; Kim, K.-N.; Lee, S.-H.; Yang, H.-M.; Kim, A.-D.; Jeon, Y.-J. Molecular docking studies of a phlorotannin, dieckol isolated from Ecklonia cava with tyrosinase inhibitory activity. Bioorganic Med. Chem. 2012, 20, 311–316. [Google Scholar] [CrossRef]
- Jiang, T.; Zhou, J.; Liu, W.; Tao, W.; He, J.; Jin, W.; Guo, H.; Yang, N.; Li, Y. The anti-inflammatory potential of protein-bound anthocyanin compounds from purple sweet potato in LPS-induced RAW264.7 macrophages. Food Res. Int. 2020, 137, 109647. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, L.; Zhang, J.; Zhang, J.; Zhang, Y.; Wang, Y.; Chen, Q.; Luo, Y.; Zhang, Y.; Li, M.; et al. Identi-fication of anthocyanins-related glutathione S-transferase (GST) genes in the genome of cultivated strawberry (Fragaria x ananassa). Int. J. Mol. Sci. 2020, 21, 8708. [Google Scholar] [CrossRef]
- Glover, B.J.; Martin, C. Anthocyanins. Curr. Biol. 2012, 22, R147–R150. [Google Scholar] [CrossRef] [Green Version]
- Oladzadabbasabadi, N.; Nafchi, A.M.; Ghasemlou, M.; Ariffin, F.; Singh, Z.; Al-Hassan, A.A. Natural anthocyanins: Sources, extraction, characterization, and suitability for smart packaging. Food Packag. Shelf. 2022, 33, 100872. [Google Scholar] [CrossRef]
- Wongwichai, T.; Teeyakasem, P.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Anthocyanins and metabolites from purple rice inhibit IL-1β-induced matrix metalloproteinases expression in human articular chondrocytes through the NF-kβ and ERK/MAPK pathway. Biomed. Pharmacoth. 2019, 112, 108610. [Google Scholar] [CrossRef] [PubMed]
- Takasawa, R.; Saeki, K.; Tao, A.; Yoshimori, A.; Uchiro, H.; Fujiwara, M.; Tanuma, S.-I. Delphinidin, a dietary anthocyanidin in berry fruits, inhibits human glyoxalase I. Bioorganic Med. Chem. 2010, 18, 7029–7033. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.S.; He, J.l.; Dong, L.C.; Yuan, L.P.; Wu, J.L.; Zu, Y.X.; Zheng, H.X. cyaniding-3-glucoside from black rice prevents renal dysfunction and renal fibrosis in streptozotoci-diabetic rats. J. Funct. Foods 2020, 72, 104062. [Google Scholar] [CrossRef]
- Kim, J.H.; Jo, Y.D.; Kim, H.-Y.; Kim, B.-R.; Nam, B. In vitro and in silico insights into she inhibitors with amide-scaffold from the leaves of Capsicum chinense Jacq. Comput. Struct. Biotec. 2018, 16, 404–411. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Z.; Song, W.; Zhao, Z.; Zhao, Y. Isolation of proanthocyanidins from Pinus thunbergii needles and tyrosinase inhibition activity. Process Biochem. 2021, 100, 245–251. [Google Scholar] [CrossRef]
- Bunea, A.; Rugină, D.; Sconţa, Z.; Pop, R.M.; Pintea, A.; Socaciu, C.; Tăbăran, F.; Grootaert, C.; Struijs, K.; VanCamp, J. An-thocyanin determination in blue berry extracts from various cultivars and their antiproliferative and apoptotic properties in b16-F10 metastatic murine melanoma cells. Phytochemistry 2013, 95, 436–444. [Google Scholar] [CrossRef]
- Chen, P.-N.; Kuo, W.-H.; Chiang, C.-L.; Chiou, H.-L.; Hsieh, Y.-S.; Chu, S.-C. Black rice anthocyanins inhibit cancer cells invasion via repressions of MMPs and u-PA expression. Chem. Interactions 2006, 163, 218–229. [Google Scholar] [CrossRef]
- Lee, C.J.; Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoom, S.K.; Han, H.J.; Kim, D.-O.; Heo, H.J. Melanogenesis regulatory activity of the ethyl acetate fraction from Arctium lappa L. leaf on α-MSH–induced B16/F10 melanoma cells. Ind. Crop. Prod. 2019, 138, 111581. [Google Scholar] [CrossRef]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Labuschagne, L.; Dubery, I.A. Unravellng the metabolic reconfiguration of the post-challenge primed state in Sorghum bicolor responding to colletotrichum sublineolum infection. Metabolites 2019, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Picariello, G.; Ferranti, P.; Garro, G.; Manganiello, G.; Chianese, L.; Coppola, R.; Addeo, F. Profiling of anthocyanins for the taxonomic assessment of ancient purebred V. vinifera red grape varieties. Food Chem. 2014, 146, 15–22. [Google Scholar] [CrossRef]
- Ulaszewska, M.; Garcia-Aloy, G.M.; Vázquez-Manjarrez, N.; Soria-Florido, M.T.; Llorach, R.; Mattivi, F.; Manach, C. Food intake biomarkers for berries and grapes. Genes Nutr. 2020, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Jhan, J.K.; Chung, Y.C.; Chen, G.H.; Chang, C.-H.; Lu, Y.-C.; Hsu, C.-K. Anthocyanin contents in the seed coat of black soya bean and their anti-human tyrosinase activity and antioxidative activity. Int. J. Cosmet. Sci. 2016, 38, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Fawole, O.A.; Makunga, N.P.; Opara, U.L. Antibacterial, antioxidant and tyrosinase-inhibiton activities of pomegranate fruit peel methanolic extract. BMC Complement Altern. Med. 2012, 12, 200. [Google Scholar] [CrossRef] [PubMed]
- Koyu, H.; Kazan, A.; Ozturk, T.K.; Yesil-Celiktas, O.; Haznedaroglu, M.Z. Optimizing subscritical water extraction of Morus nigra L. fruits for maximization of tyrosinase inhibitory activity. J. Supercrit. Fluid. 2017, 127, 15–22. [Google Scholar] [CrossRef]
- Koyu, H.; Kazan, A.; Demir, S.; Haznedaroglu, M.Z.; Yesil-Celiktas, O. Optimization of microwave assisted extraction of Morus nigra L. fruits maximizing tyrosinase inhibitory activity with isolation of bioactive constituents. Food Chem. 2018, 248, 183–191. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Compound a | Inhibitory Ratio at 50 μM | IC50 (μM) d | Binding Mode (ki, μM) |
|---|---|---|---|
| 1 | 176.5 ± 11.6 | 3.7 ± 0.8 | Competitive inhibitor (2.8 ± 0.5) |
| 2 | 69.2 ± 1.0 | 10.3 ± 1.0 | Competitive inhibitor (9.0 ± 2.1) |
| 3 | 55.2 ± 0.9 | 41.3 ± 3.2 | Competitive inhibitor (51.9 ± 3.8) |
| 4 | −1.9 ± 40.0 | N.T. | N.T. c |
| 5 | 47.5 ± 3.2 | N.T. | N.T. c |
| 6 | 6.5 ± 2.1 | N.T. | N.T. c |
| Kojic acid b | 31.4 ± 1.1 |
| Autodock Energy (kcal/mol) | Hydrogen Bonds (Å) | |
|---|---|---|
| 1 | −5.36 | His244(2.71), Glu256(2.65, 2.72), Gly281(2.53) |
| 2 | −4.96 | Asn260(2.78), Arg268(3.13), Gly281(2.94), Ser282(2.73,3.15) |
| 3 | −4.61 | Asn260(2.83), Arg268(3.14, 3.14) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.Y.; Kim, J.H.; Su, X.; Kim, J.A. The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase. Molecules 2022, 27, 5703. https://doi.org/10.3390/molecules27175703
Yang SY, Kim JH, Su X, Kim JA. The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase. Molecules. 2022; 27(17):5703. https://doi.org/10.3390/molecules27175703
Chicago/Turabian StyleYang, Seo Young, Jang Hoon Kim, Xiangdong Su, and Jeong Ah Kim. 2022. "The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase" Molecules 27, no. 17: 5703. https://doi.org/10.3390/molecules27175703
APA StyleYang, S. Y., Kim, J. H., Su, X., & Kim, J. A. (2022). The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase. Molecules, 27(17), 5703. https://doi.org/10.3390/molecules27175703