
Antiviral Effect of Bovine Lactoferrin against Enterovirus E

Abstract
:1. Introduction
2. Results
2.1. Choice of Lactoferrin Concentrations Tested in the Experiment
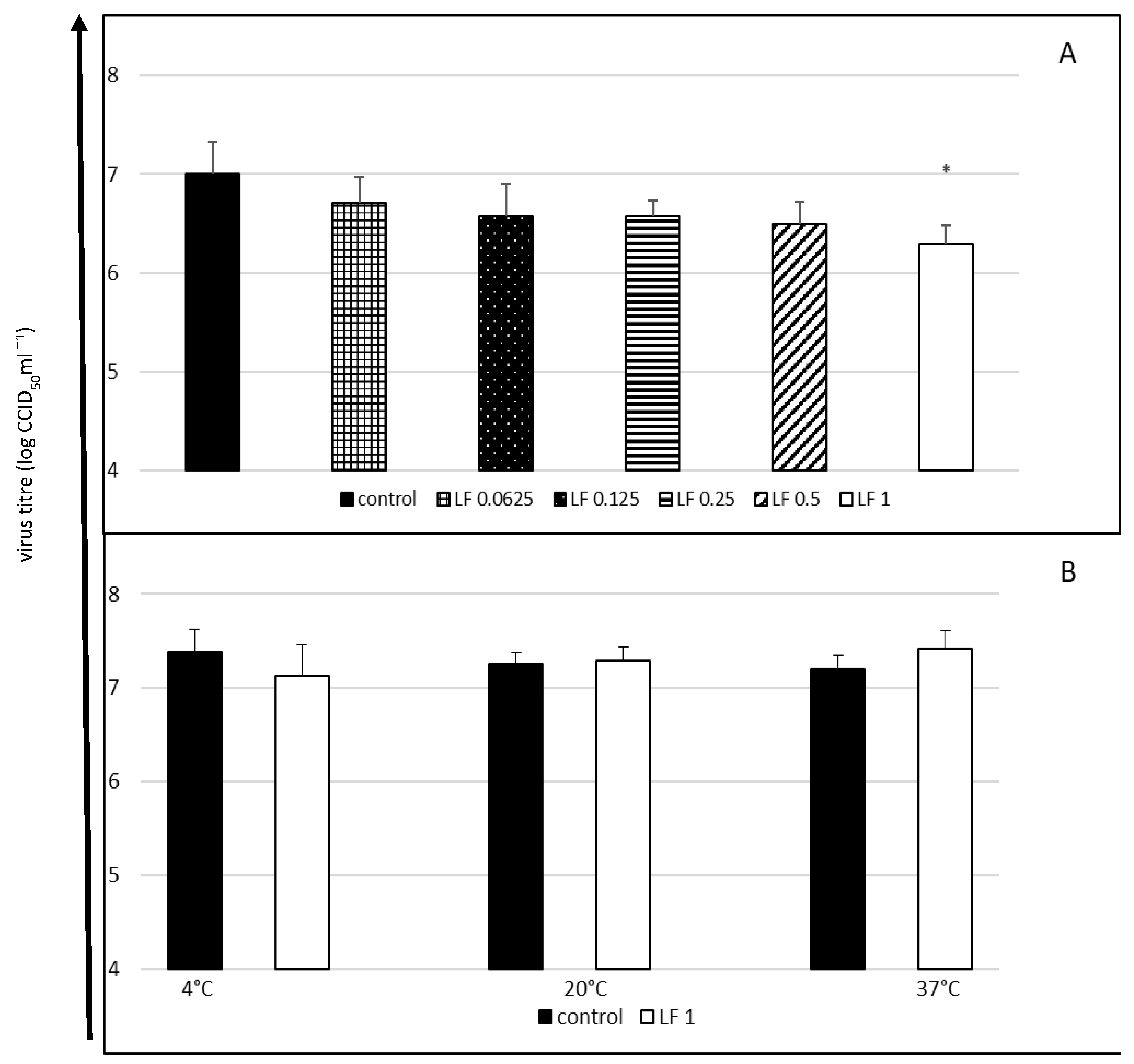
2.2. Cytopathic Effect Inhibition and Virucidal Activity of Bovine Lactoferrin against Enterovirus E
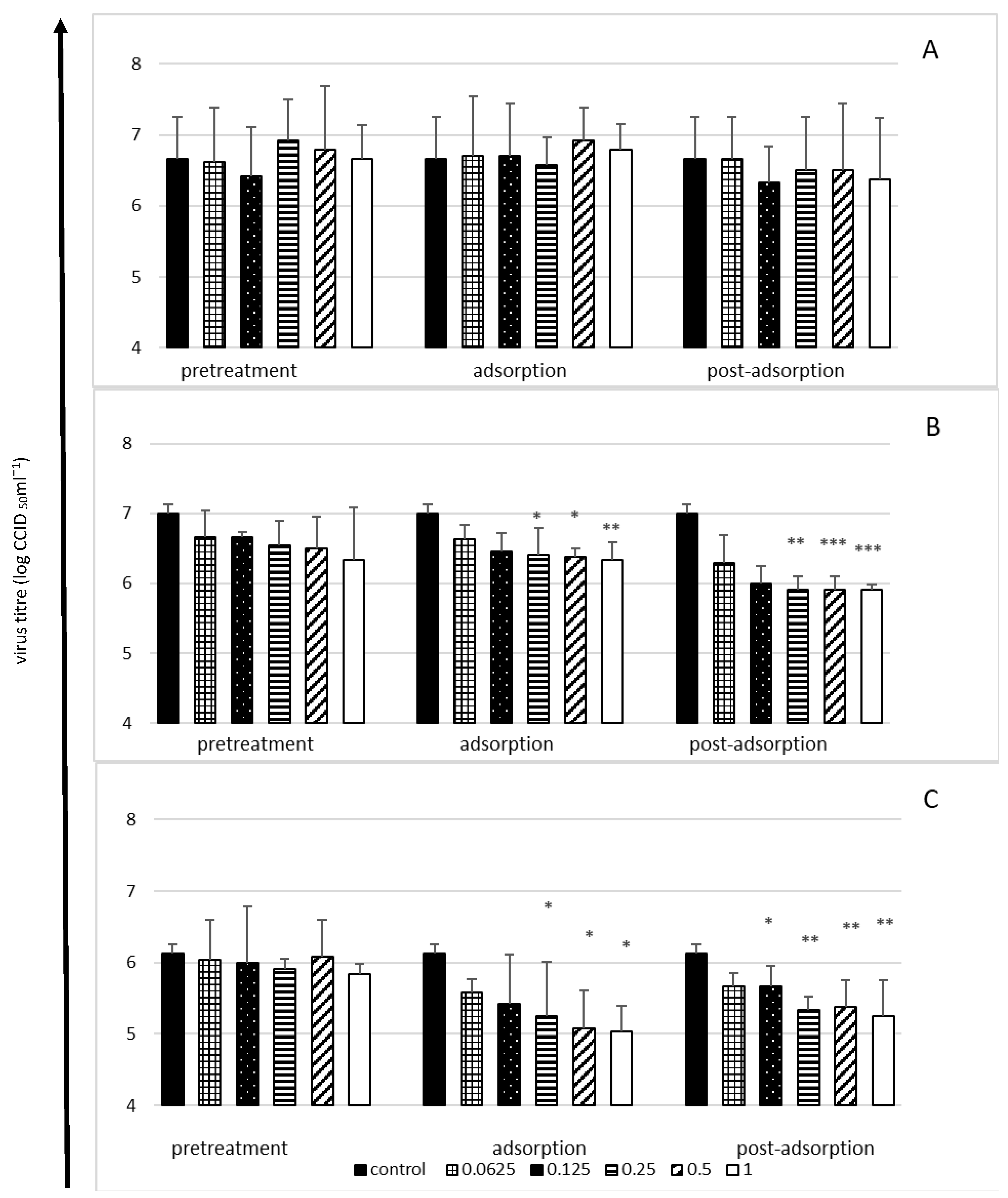
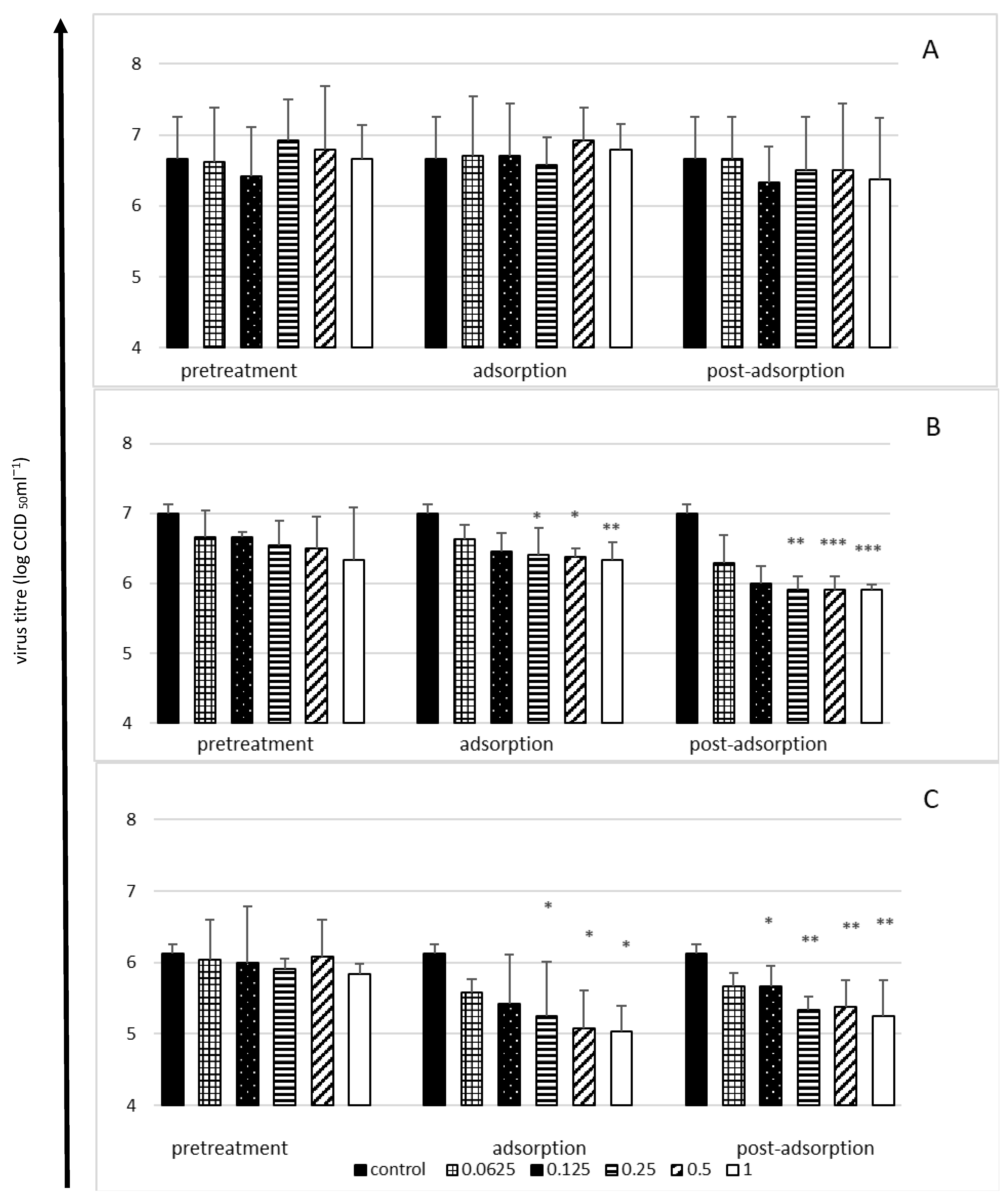
2.3. Viral Yield Reduction in the Presence of Bovine Lactoferrin—A Time-of-Addition Assay
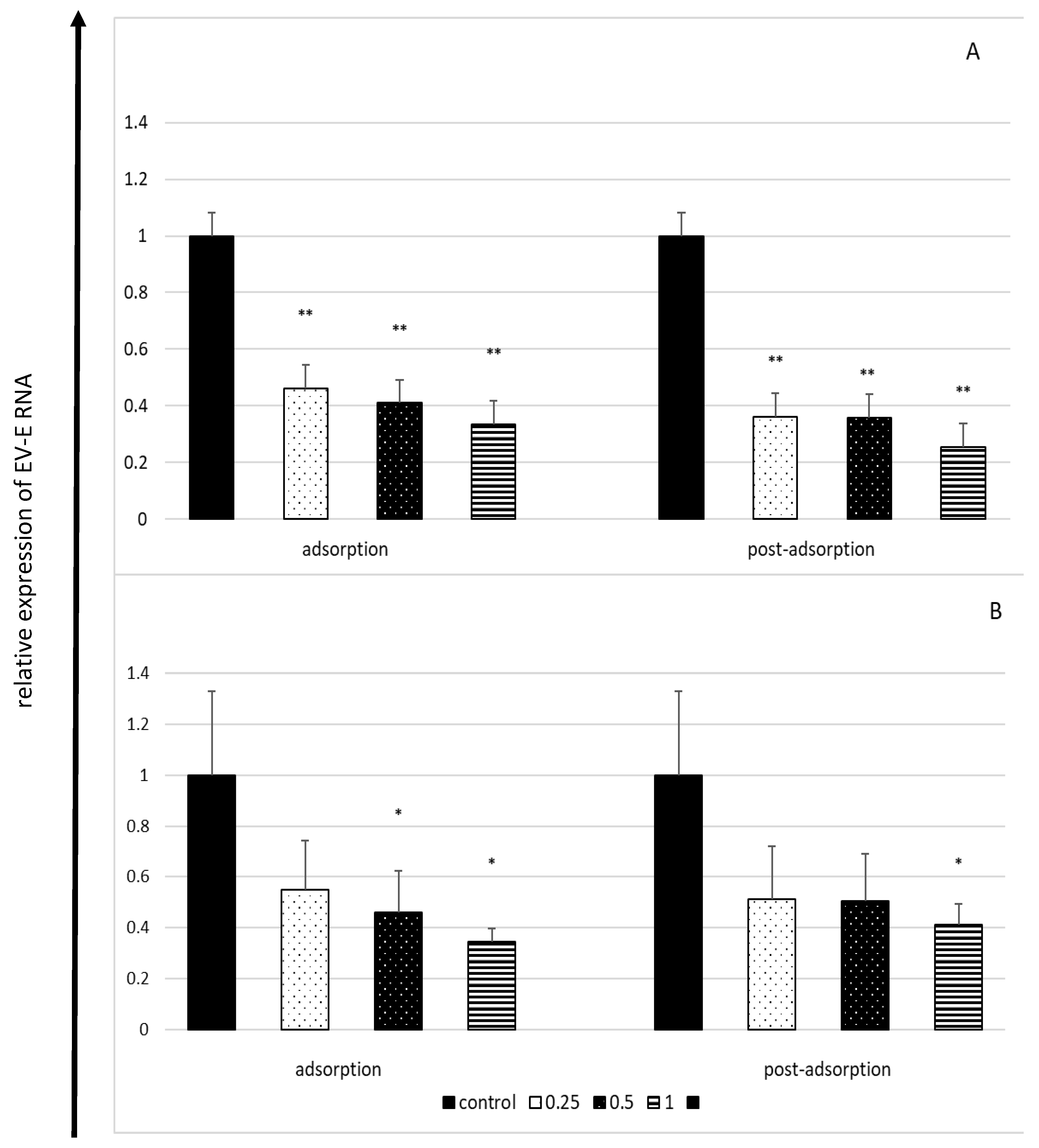
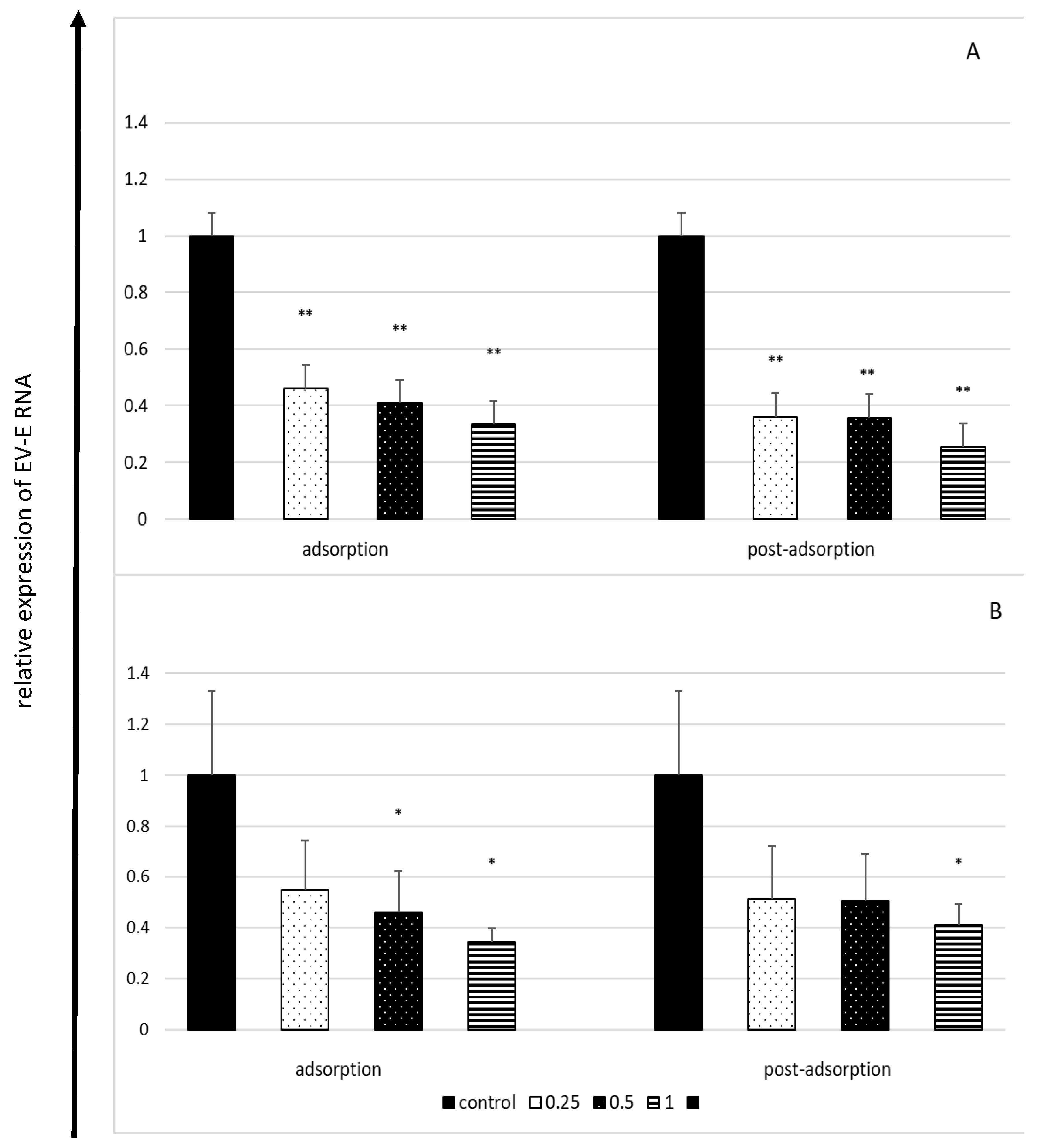
2.4. Effect of Lactoferrin on Viral RNA Load in Enterovirus E-Infected Cells
3. Discussion
4. Materials and Methods
4.1. Cells, Virus and Bovine Lactoferrin
4.2. Antiviral Assays
4.2.1. Cytopathic Effect Inhibition Assay
4.2.2. Virucidal (Virus Neutralisation) Assay
4.2.3. Yield Reduction Assay
- -
- Cell pretreatment stage: Cells were pretreated with various lactoferrin concentrations (2 h at 37 °C), washed three times with medium and then infected with virus (1 h at 37 °C)
- -
- Adsorption stage: Various lactoferrin concentrations were added simultaneously with the virus inoculum (1 h at 37 °C), followed by washing.
- -
- Post-adsorption stage: Various lactoferrin concentrations were added to cells just after virus adsorption (1 h at 37 °C) and were present in the medium to the end of the experiment.
4.3. RNA Isolation and RT-qPCR
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kalter, S.S. Animal “orphan” enteroviruses. Bull. World Health Organ. 1960, 22, 319–337. [Google Scholar] [PubMed]
- Blas-Machado, U.; Saliki, J.T.; Boileau, M.J.; Goens, S.D.; Caseltine, S.L.; Duffy, J.C.; Welsh, R.D. Fatal ulcerative and hemorrhagic typhlocolitis in a pregnant heifer associated with natural bovine enterovirus type-1 infection. Vet. Pathol. 2007, 44, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Zell, R.; Krumbholz, A.; Dauber, M.; Hoey, E.; Wutzler, P. Molecular-based reclassification of the bovine enteroviruses. J. Gen. Virol. 2006, 87 Pt 2, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, H.; Bao, J.; Guo, Y.; Peng, T.; Zhou, P.; Zhang, W.; Ma, B.; Wang, J.; Gao, M. Characterization of an Enterovirus species E isolated from naturally infected bovine in China. Virus Res. 2014, 191, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xing, Z.; Gai, X.; Li, S.; San, Z.; Wang, X. Identification of a novel enterovirus E isolates HY12 from cattle with severe respiratory and enteric diseases. PLoS ONE 2014, 15, e97730. [Google Scholar] [CrossRef]
- McGill, J.L.; Sacco, R.E. The immunology of bovine respiratory disease: Recent advancements. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 333–348. [Google Scholar] [CrossRef]
- Jiménez-Clavero, M.A.; Escribano-Romero, E.; Mansilla, C.; Gómez, N.; Córdoba, L.; Roblas, N.; Ponz, F.; Ley, V.; Sáiz, J.C. Survey of bovine enterovirus in biological and environmental samples by a highly sensitive real-time reverse transcription-PCR. Appl. Environ. Microbiol. 2005, 71, 3536–3543. [Google Scholar] [CrossRef]
- Jain, N.C.; Batra, S.K. Isolation and characterisation of bovine enteroviruses. Indian J. Virol. 1985, 1, 17. [Google Scholar]
- Hamblin, C.; Knowles, N.J.; Hedger, R.S. Isolation and identification of bovid enteroviruses from free-living wild animals in Botswana. Vet. Rec. 1985, 116, 238–239. [Google Scholar] [CrossRef]
- Gür, S.; Yapkiç, O.; Yilmaz, A. Serological survey of bovine enterovirus type 1 in different mammalian species in Turkey. Zoonoses Public Health 2008, 55, 106–111. [Google Scholar] [CrossRef]
- Gür, S.; Gürçay, M.; Seyrek, A. A study regarding bovine enterovirus type 1 infection in domestic animals and humans: An evaluation from the zoonotic aspect. J. Vet. Med. Sci. 2019, 81, 1824–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fieldhouse, J.K.; Wang, X.; Mallinson, K.A.; Tsao, R.W.; Gray, G.C. A systematic review of evidence that enteroviruses may be zoonotic. Emerg. Microbes Infect. 2018, 7, 164. [Google Scholar] [CrossRef] [PubMed]
- Racaniello, V.R. One hundred years of poliovirus pathogenesis. Virology 2006, 344, 9–16. [Google Scholar] [CrossRef]
- EN 14675:2015; Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Virucidal Activity of Chemical Disinfectants and Antiseptics Used in the Veterinary Area—Test Method and Requirements (Phase 2, Step 1). Polski Komitet Normalizacyjny: Warszawa, Poland, 2015.
- Centers for Disease Control and Prevention. Prevention and Treatment of Non-Polio Enterovirus Infection. 2022. Available online: https://www.cdc.gov/non-polio-enterovirus/about/prevention-treatment.html (accessed on 8 July 2022).
- Dal Pozzo, F.; Thiry, E. Antiviral chemotherapy in veterinary medicine: Current applications and perspectives. Rev. Sci. Tech. 2014, 33, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Małaczewska, J.; Rotkiewicz, Z. Lactoferrin—A multipotential protein. Medycyna Wet. 2007, 63, 136–139. [Google Scholar]
- Małaczewska, J.; Rotkiewicz, Z.; Siwicki, A.K. Lactoferrin—Mechanisms for antiviral activity. Medycyna Wet. 2006, 62, 1104–1107. [Google Scholar]
- Wakabayashi, H.; Oda, H.; Yamauchi, K.; Abe, F. Lactoferrin for prevention of common viral infections. J. Infect. Chemother. 2014, 20, 666–671. [Google Scholar] [CrossRef]
- Jenista, J.A.; Menegus, M.A. Neonatal enterovirus infection. Indian J. Pediatr. 1988, 55, 191–196. [Google Scholar] [CrossRef]
- Pietrantoni, A.; Ammendolia, M.G.; Tinari, A.; Siciliano, R.; Valenti, P.; Superti, F. Bovine lactoferrin peptidic fragments involved in inhibition of Echovirus 6 in vitro infection. Antivir. Res. 2006, 69, 98–106. [Google Scholar] [CrossRef]
- Chen, H.L.; Wang, L.C.; Chang, C.H.; Yen, C.C.; Cheng, W.T.; Wu, S.C.; Hung, C.M.; Kuo, M.F.; Chen, C.M. Recombinant porcine lactoferrin expressed in the milk of transgenic mice protects neonatal mice from a lethal challenge with enterovirus type 71. Vaccine 2008, 26, 891–898. [Google Scholar] [CrossRef]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Siwicki, A.K. Antiviral effects of nisin, lysozyme, lactoferrin and their mixtures against bovine viral diarrhoea virus. BMC Vet. Res. 2019, 15, 318. [Google Scholar] [CrossRef] [PubMed]
- Benschop, K.S.; van der Avoort, H.G.; Duizer, E.; Koopmans, M.P. Antivirals against enteroviruses: A critical review from a public-health perspective. Antivir. Ther. 2015, 20, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.C.; Weng, K.F.; Chang, S.C.; Lin, J.Y.; Huang, P.N.; Shih, S.R. Development of antiviral agents for enteroviruses. J. Antimicrob. Chemother. 2008, 62, 1169–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikonov, O.S.; Chernykh, E.S.; Garber, M.B.; Nikonova, E.Y. Enteroviruses: Classification, diseases they cause, and approaches to development of antiviral drugs. Biochemistry 2017, 82, 1615–1631. [Google Scholar] [CrossRef] [PubMed]
- Bębenek, E.; Jastrzębska, M.; Kadela-Tomanek, M.; Chrobak, E.; Orzechowska, B.; Zwolińska, K.; Latocha, M.; Mertas, A.; Czuba, Z.; Boryczka, S. Novel Triazole Hybrids of Betulin: Synthesis and Biological Activity Profile. Molecules 2017, 22, 1876. [Google Scholar] [CrossRef]
- Pęcak, P.; Orzechowska, B.; Chrobak, E.; Boryczka, S. Novel betulin dicarboxylic acid ester derivatives as potent antiviral agents: Design, synthesis, biological evaluation, structure-activity relationship and in-silico study. Eur. J. Med. Chem. 2021, 225, 113738. [Google Scholar] [CrossRef]
- Sobhy, N.M.; Mor, S.K.; Mohammed, M.E.; Bastawecy, I.M.; Fakhry, H.M.; Youssef, C.R.; Abouzeid, N.Z.; Goyal, S.M. Isolation and molecular characterization of bovine enteroviruses in Egypt. Vet. J. 2015, 206, 317–321. [Google Scholar] [CrossRef]
- Ji, C.; Zhang, Y.; Sun, R.; Pan, Z.; Ma, J.; Yao, H. Isolation and Identification of Two Clinical Strains of the Novel Genotype Enterovirus E5 in China. Microbiol. Spectr. 2022, 10, e0266221. [Google Scholar] [CrossRef]
- Lin, T.Y.; Chu, C.; Chiu, C.H. Lactoferrin inhibits enterovirus 71 infection of human embryonal rhabdomyosarcoma cells in vitro. J. Infect. Dis. 2002, 186, 1161–1164. [Google Scholar] [CrossRef]
- Weng, T.Y.; Chen, L.C.; Shyu, H.W.; Chen, S.H.; Wang, J.R.; Yu, C.K.; Lei, H.Y.; Yeh, T.M. Lactoferrin inhibits enterovirus 71 infection by binding to VP1 protein and host cells. Antivir. Res. 2005, 67, 31–37. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Pietrantoni, A.; Tinari, A.; Valenti, P.; Superti, F. Bovine lactoferrin inhibits echovirus endocytic pathway by interacting with viral structural polypeptides. Antivir. Res. 2007, 73, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Furlund, C.B.; Kristoffersen, A.B.; Devold, T.G.; Vegarud, G.E.; Jonassen, C.M. Bovine lactoferrin digested with human gastrointestinal enzymes inhibits replication of human echovirus 5 in cell culture. Nutr. Res. 2012, 32, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Superti, F.; Ammendolia, M.G.; Rossi, P.; Valenti, P.; Seganti, L. Inhibition of poliovirus type 1 infection by iron-, manganese- and zinc-saturated lactoferrin. Med. Microbiol. Immunol. 1999, 187, 199–204. [Google Scholar] [CrossRef]
- Ji, Z.S.; Mahley, R.W. Lactoferrin binding to heparan sulfate proteoglycans and the LDL receptor-related protein. Further evidence supporting the importance of direct binding of remnant lipoproteins to HSPG. Arteiosceler. Thromb. 1994, 14, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiaka, S.; Rahpaya, S.S.; Otomaru, K.; Aoki, H.; Kishimoto, M.; Naoi, Y.; Omatsu, T.; Sano, K.; Okazaki-Terashima, S.; Katayama, Y.; et al. Identification of a novel bovine enterovirus possessing highly divergent aminoacid sequences in capsid protein. BMC Microbiol. 2017, 17, 18. [Google Scholar] [CrossRef]
- Jiang, P.; Liu, Y.; Ma, H.C.; Paul, A.V.; Wimmer, E. Picornavirus morphogenesis. Microbiol. Mol. Biol. Rev. 2014, 78, 418–437. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′–3′) | Amplicon Size | GenBank Accession No. |
|---|---|---|---|
| EV-E802 for | AAAGGGGGCTGTCGAAACCA | 802 | DQ092769.1 |
| EV-E 802 rev | GCTAGTGGGCTCAGACTCCG | ||
| EV-E 183 for | TACGCCTTTCGTGGCTTGGA | 183 | |
| EV-E 183 rev | TTGCTTTTCCTGGCTTGCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wróbel, M.; Małaczewska, J.; Kaczorek-Łukowska, E. Antiviral Effect of Bovine Lactoferrin against Enterovirus E. Molecules 2022, 27, 5569. https://doi.org/10.3390/molecules27175569
Wróbel M, Małaczewska J, Kaczorek-Łukowska E. Antiviral Effect of Bovine Lactoferrin against Enterovirus E. Molecules. 2022; 27(17):5569. https://doi.org/10.3390/molecules27175569
Chicago/Turabian StyleWróbel, Małgorzata, Joanna Małaczewska, and Edyta Kaczorek-Łukowska. 2022. "Antiviral Effect of Bovine Lactoferrin against Enterovirus E" Molecules 27, no. 17: 5569. https://doi.org/10.3390/molecules27175569
APA StyleWróbel, M., Małaczewska, J., & Kaczorek-Łukowska, E. (2022). Antiviral Effect of Bovine Lactoferrin against Enterovirus E. Molecules, 27(17), 5569. https://doi.org/10.3390/molecules27175569