
Withaferin A—A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases

 ,
,  ,
, 

 ,
, 
 ,
,  and
and
Abstract
1. Introduction
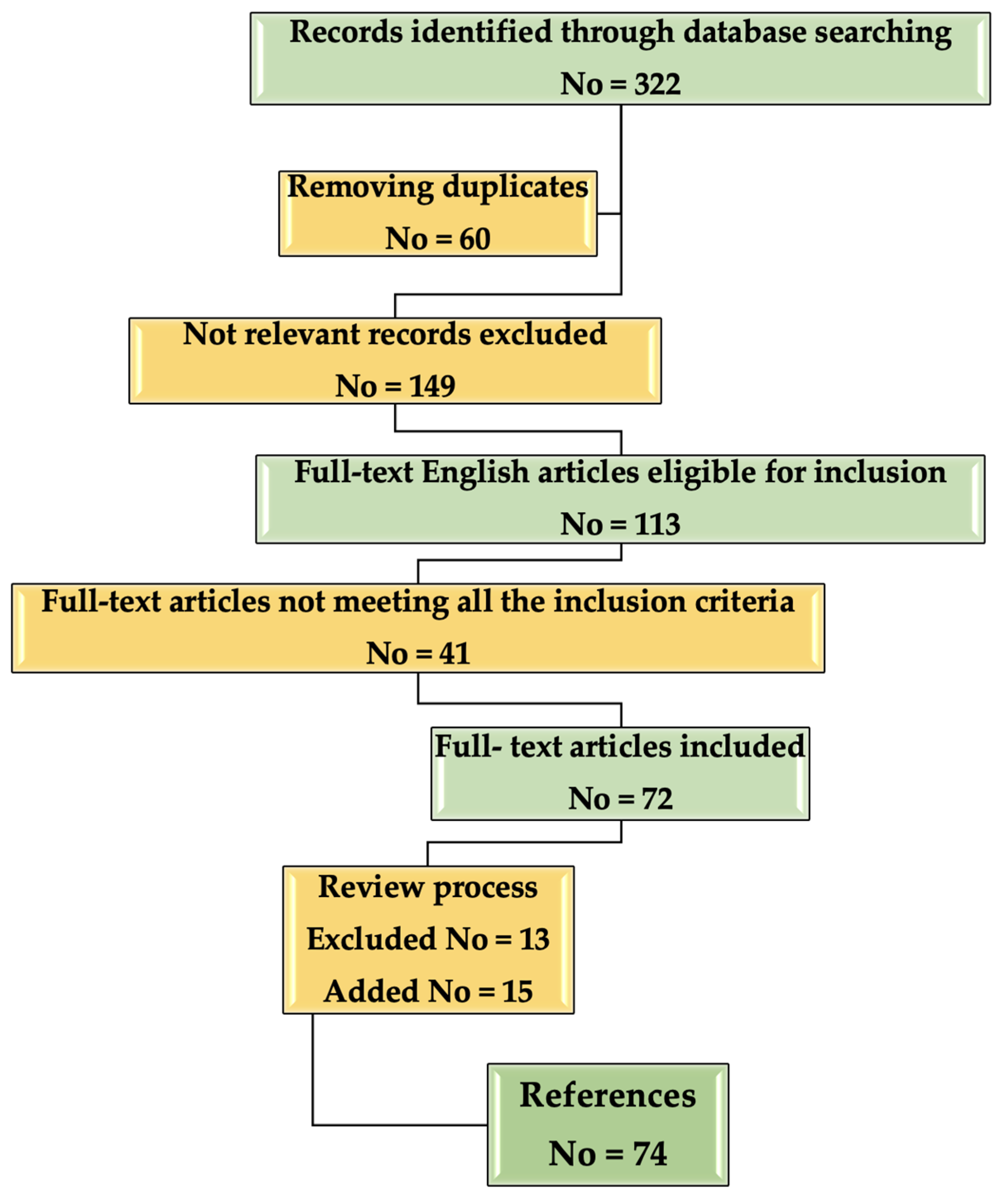
2. Methodology
3. Withaferin in Skin Cancers
3.1. Chemopreventive Effects
3.2. Antitumor Effects
3.3. Support Therapy
4. Withaferin Actions in Other Skin Diseases
4.1. In Scleroderma
4.2. In Disorders of Pigmentation
4.2.1. Hyperpigmentation
4.2.2. Hypopigmentation (Leucoderma)
4.3. Viral Infections
4.3.1. Herpes Simplex
4.3.2. Papilloma Virus
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sehgal, V. Diagnosis and Treatment of Common Skin Diseases, 5th ed.; Jaypee Brothers Medical Publishers Pvt Ltd.: Guwahati, India, 2016. [Google Scholar]
- Behl, T.; Sharma, A.; Sharma, L.; Sehgal, A.; Zengin, G.; Brata, R.; Fratila, O.; Bungau, S. Exploring the Multifaceted Therapeutic Potential of Withaferin A and Its Derivatives. Biomedicines 2020, 8, 571. [Google Scholar] [CrossRef] [PubMed]
- Sivasankarapillai, V.S.; Madhu Kumar Nair, R.; Rahdar, A.; Bungau, S.; Zaha, D.C.; Aleya, L.; Tit, D.M. Overview of the anticancer activity of Withaferin A, an active constituent of the Indian Ginseng Withania somnifera. Environ. Sci. Pollut. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bungau, S.G.; Popa, V.-C. Between Religion and Science Some Aspects Concerning Illness and Healing in Antiquity. Transylv. Rev. 2015, 24, 3–18. [Google Scholar]
- Kaur, K.; Singh, P.; Kaur, K.; Bhandawat, B.; Nogia, P.; Pati, P.K. Development of robust in vitro culture protocol for the propagation of genetically and phytochemically stable plants of Withania somnifera (L.) Dunal (Ashwagandha). Ind. Crops Prod. 2021, 166, 113428. [Google Scholar] [CrossRef]
- Samadi, A.K. Potential Anticancer Properties and Mechanisms of Action of Withanolides. Enzymes 2015, 37, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Khalil, R.; Green, R.; Mohapatra, S.S.; Mohapatra, S. Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology. Int. J. Mol. Sci. 2019, 20, 5310. [Google Scholar] [CrossRef]
- Vanden Berghe, W.; Sabbe, L.; Kaileh, M.; Haegeman, G.; Heyninck, K. Molecular insight in the multifunctional activities of Withaferin A. Biochem. Pharmacol. 2012, 84, 1282–1291. [Google Scholar] [CrossRef]
- Vyas, A.R.; Singh, S.V. Molecular targets and mechanisms of cancer prevention and treatment by withaferin a, a naturally occurring steroidal lactone. AAPS J. 2014, 16, 1–10. [Google Scholar] [CrossRef]
- Yu, Y.; Hamza, A.; Zhang, T.; Gu, M.; Zou, P.; Newman, B.; Li, Y.; Gunatilaka, A.A.; Zhan, C.G.; Sun, D. Withaferin A targets heat shock protein 90 in pancreatic cancer cells. Biochem. Pharmacol. 2010, 79, 542–551. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ranga, R.S.; Burikhanov, R.; Han, S.S.; Chendil, D. Par-4-dependent apoptosis by the dietary compound withaferin A in prostate cancer cells. Cancer Res. 2007, 67, 246–253. [Google Scholar] [CrossRef]
- Maitra, R.; Porter, M.A.; Huang, S.; Gilmour, P. Inhibition of NFκB by the natural product Withaferin A in cellular models of Cystic Fibrosis inflammation. J. Inflamm. 2009, 6, 15. [Google Scholar] [CrossRef]
- Stan, S.D.; Hahm, E.R.; Warin, R.; Singh, S.V. Withaferin A causes FOXO3a- and Bim-dependent apoptosis and inhibits growth of human breast cancer cells in vivo. Cancer Res. 2008, 68, 7661–7669. [Google Scholar] [CrossRef]
- Mayola, E.; Gallerne, C.; Esposti, D.D.; Martel, C.; Pervaiz, S.; Larue, L.; Debuire, B.; Lemoine, A.; Brenner, C.; Lemaire, C. Withaferin A induces apoptosis in human melanoma cells through generation of reactive oxygen species and down-regulation of Bcl-2. Apoptosis 2011, 16, 1014–1027. [Google Scholar] [CrossRef] [PubMed]
- Mandal, C.; Dutta, A.; Mallick, A.; Chandra, S.; Misra, L.; Sangwan, R.S.; Mandal, C. Withaferin A induces apoptosis by activating p38 mitogen-activated protein kinase signaling cascade in leukemic cells of lymphoid and myeloid origin through mitochondrial death cascade. Apoptosis 2008, 13, 1450–1464. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.C.; Choi, B.Y. Withaferin-A--A Natural Anticancer Agent with Pleitropic Mechanisms of Action. Int. J. Mol. Sci. 2016, 17, 290. [Google Scholar] [CrossRef] [PubMed]
- Dom, M.; Offner, F.; Vanden Berghe, W.; Van Ostade, X. Proteomic characterization of Withaferin A-targeted protein networks for the treatment of monoclonal myeloma gammopathies. J. Proteom. 2018, 179, 17–29. [Google Scholar] [CrossRef]
- Narayan, M.; Seeley, K.W.; Jinwal, U.K. Identification and quantitative analysis of cellular proteins affected by treatment with withaferin a using a SILAC-based proteomics approach. J. Ethnopharmacol. 2015, 175, 86–92. [Google Scholar] [CrossRef]
- Narayan, M.; Zhang, J.; Braswell, K.; Gibson, C.; Zitnyar, A.; Lee, D.C.; Varghese-Gupta, S.; Jinwal, U.K. Withaferin A Regulates LRRK2 Levels by Interfering with the Hsp90- Cdc37 Chaperone Complex. Curr. Aging Sci. 2015, 8, 259–265. [Google Scholar] [CrossRef]
- Zhang, L.; Nemzow, L.; Chen, H.; Lubin, A.; Rong, X.; Sun, Z.; Harris, T.K.; Gong, F. The deubiquitinating enzyme USP24 is a regulator of the UV damage response. Cell Rep. 2015, 10, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Dyba, T.; Randi, G.; Bettio, M.; Gavin, A.; Visser, O.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries and 25 major cancers in 2018. Eur. J. Cancer 2018, 103, 356–387. [Google Scholar] [CrossRef]
- Rotaru, M.; Jitian, C.R.; Iancu, G.M. A 10-year retrospective study of melanoma stage at diagnosis in the academic emergency hospital of Sibiu county. Oncol. Lett. 2019, 17, 4145–4148. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Long, G.V. Ipilimumab Combined with Nivolumab: A Standard of Care for the Treatment of Advanced Melanoma? Clin. Cancer Res. 2016, 22, 3992. [Google Scholar] [CrossRef] [PubMed]
- Long, G.V.; Flaherty, K.T.; Stroyakovskiy, D.; Gogas, H.; Levchenko, E.; de Braud, F.; Larkin, J.; Garbe, C.; Jouary, T.; Hauschild, A.; et al. Dabrafenib plus trametinib versus dabrafenib monotherapy in patients with metastatic BRAF V600E/K-mutant melanoma: Long-term survival and safety analysis of a phase 3 study. Ann. Oncol. 2017, 28, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Marshall, G.M.; Bell, J.L.; Koach, J.; Tan, O.; Kim, P.; Malyukova, A.; Thomas, W.; Sekyere, E.O.; Liu, T.; Cunningham, A.M.; et al. TRIM16 acts as a tumour suppressor by inhibitory effects on cytoplasmic vimentin and nuclear E2F1 in neuroblastoma cells. Oncogene 2010, 29, 6172–6183. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Zhao, Y.F. Withaferin A suppresses tumor promoter 12-O-tetradecanoylphorbol 13-acetate-induced decreases in isocitrate dehydrogenase 1 activity and mitochondrial function in skin epidermal JB6 cells. Cancer Sci. 2013, 104, 143–148. [Google Scholar] [CrossRef]
- Devi, P.U.; Kamath, R.; Rao, B.S. Radiosensitization of a mouse melanoma by withaferin A: In vivo studies. Indian J. Exp. Biol. 2000, 38, 432–437. [Google Scholar]
- Samadi, A.K.; Cohen, S.M.; Mukerji, R.; Chaguturu, V.; Zhang, X.; Timmermann, B.N.; Cohen, M.S.; Person, E.A. Natural withanolide withaferin A induces apoptosis in uveal melanoma cells by suppression of Akt and c-MET activation. Tumor Biol. 2012, 33, 1179–1189. [Google Scholar] [CrossRef]
- Prakash, J.; Gupta, S.K.; Dinda, A.K. Withania somnifera root extract prevents DMBA-induced squamous cell carcinoma of skin in Swiss albino mice. Nutr. Cancer 2002, 42, 91–97. [Google Scholar] [CrossRef]
- Davis, L.; Kuttan, G. Effect of Withania somnifera on DMBA induced carcinogenesis. J. Ethnopharmacol. 2001, 75, 165–168. [Google Scholar] [CrossRef]
- Xu, C.; Huang, M.T.; Shen, G.; Yuan, X.; Lin, W.; Khor, T.O.; Conney, A.H.; Kong, A.N. Inhibition of 7,12-dimethylbenz(a)anthracene-induced skin tumorigenesis in C57BL/6 mice by sulforaphane is mediated by nuclear factor E2-related factor 2. Cancer Res. 2006, 66, 8293–8296. [Google Scholar] [CrossRef]
- Li, W.; Zhang, C.; Du, H.; Huang, V.; Sun, B.; Harris, J.P.; Richardson, Q.; Shen, X.; Jin, R.; Li, G.; et al. Withaferin A suppresses the up-regulation of acetyl-coA carboxylase 1 and skin tumor formation in a skin carcinogenesis mouse model. Mol. Carcinog. 2016, 55, 1739–1746. [Google Scholar] [CrossRef]
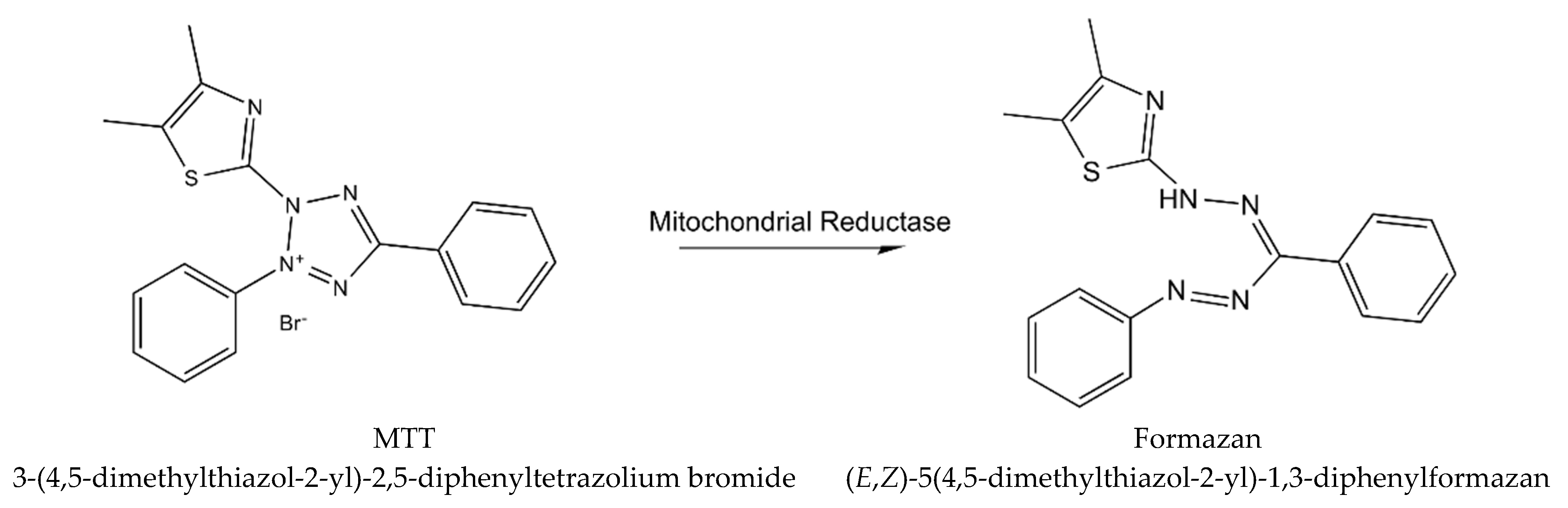
- Vistica, D.T.; Skehan, P.; Scudiero, D.; Monks, A.; Pittman, A.; Boyd, M.R. Tetrazolium-based assays for cellular viability: A critical examination of selected parameters affecting formazan production. Cancer Res. 1991, 51, 2515–2520. [Google Scholar] [PubMed]
- Halder, B.; Singh, S.; Thakur, S.S. Withania somnifera Root Extract Has Potent Cytotoxic Effect against Human Malignant Melanoma Cells. PLoS ONE 2015, 10, e0137498. [Google Scholar] [CrossRef]
- Malik, A.; Sultana, M.; Qazi, A.; Qazi, M.H.; Parveen, G.; Waquar, S.; Ashraf, A.B.; Rasool, M. Role of Natural Radiosensitizers and Cancer Cell Radioresistance: An Update. Anal. Cell. Pathol. 2016, 2016, 6146595. [Google Scholar] [CrossRef] [PubMed]
- Kalthur, G.; Pathirissery, U.D. Enhancement of the response of B16F1 melanoma to fractionated radiotherapy and prolongation of survival by withaferin A and/or hyperthermia. Integr. Cancer Ther. 2010, 9, 370–377. [Google Scholar] [CrossRef]
- Mathur, S.; Kaur, P.; Sharma, M.; Katyal, A.; Singh, B.; Tiwari, M.; Chandra, R. The treatment of skin carcinoma, induced by UV B radiation, using 1-oxo-5beta, 6beta-epoxy-witha-2-enolide, isolated from the roots of Withania somnifera, in a rat model. Phytomedicine 2004, 11, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Samadi, A.K.; Cohen, M.S.; Timmermann, B.N. Anti-proliferative withanolides from the Solanaceae: A structure-activity study. Pure Appl. Chem. 2012, 84, 1353–1367. [Google Scholar] [CrossRef]
- Uma Devi, P.; Kamath, R. Radiosensitizing effect of withaferin A combined with hyperthermia on mouse fibrosarcoma and melanoma. J. Radiat. Res. 2003, 44, 1–6. [Google Scholar] [CrossRef]
- Samadi, A.K.; Tong, X.; Mukerji, R.; Zhang, H.; Timmermann, B.N.; Cohen, M.S. Withaferin A, a cytotoxic steroid from Vassobia breviflora, induces apoptosis in human head and neck squamous cell carcinoma. J. Nat. Prod. 2010, 73, 1476–1481. [Google Scholar] [CrossRef]
- Sutton, S.K.; Koach, J.; Tan, O.; Liu, B.; Carter, D.R.; Wilmott, J.S.; Yosufi, B.; Haydu, L.E.; Mann, G.J.; Thompson, J.F.; et al. TRIM16 inhibits proliferation and migration through regulation of interferon beta 1 in melanoma cells. Oncotarget 2014, 5, 10127–10139. [Google Scholar] [CrossRef][Green Version]
- Thaiparambil, J.T.; Bender, L.; Ganesh, T.; Kline, E.; Patel, P.; Liu, Y.; Tighiouart, M.; Vertino, P.M.; Harvey, R.D.; Garcia, A.; et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Int. J. Cancer 2011, 129, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Careta, M.F.; Romiti, R. Localized scleroderma: Clinical spectrum and therapeutic update. An. Bras. Dermatol. 2015, 90, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Denton, C.P.; Khanna, D. Systemic sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Jinnin, M. Mechanisms of skin fibrosis in systemic sclerosis. J. Dermatol. 2010, 37, 11–25. [Google Scholar] [CrossRef]
- Bale, S.; Pulivendala, G.; Godugu, C. Withaferin A attenuates bleomycin-induced scleroderma by targeting FoxO3a and NF-κβ signaling: Connecting fibrosis and inflammation. Biofactors 2018, 44, 507–517. [Google Scholar] [CrossRef]
- Halder, R.M.; Nootheti, P.K. Ethnic skin disorders overview. J. Am. Acad. Dermatol. 2003, 48, S143–S148. [Google Scholar] [CrossRef]
- Sivamani, R.; Clark, A. Phytochemicals in the treatment of hyperpigmentation. Bot. Targets Ther. 2016, 6, 89–96. [Google Scholar] [CrossRef]
- Rendon, M.I.; Berson, D.S.; Cohen, J.L.; Roberts, W.E.; Starker, I.; Wang, B. Evidence and considerations in the application of chemical peels in skin disorders and aesthetic resurfacing. J. Clin. Aesthet. Dermatol. 2010, 3, 32–43. [Google Scholar]
- Plensdorf, S.; Livieratos, M.; Dada, N. Pigmentation Disorders: Diagnosis and Management. Am. Fam. Physician 2017, 96, 797–804. [Google Scholar]
- Whitton, M.E.; Ashcroft, D.M.; Barrett, C.W.; Gonzalez, U. Interventions for vitiligo. Cochrane Database Syst. Rev. 2006, 1, CD003263. [Google Scholar]
- Matsuzawa, A.; Ichijo, H. Redox control of cell fate by MAP kinase: Physiological roles of ASK1-MAP kinase pathway in stress signaling. Biochim. Biophys. Acta 2008, 1780, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Wakabayashi, Y.; Wakamatsu, K.; Imokawa, G. An extract of Withania somnifera attenuates endothelin-1-stimulated pigmentation in human epidermal equivalents through the interruption of PKC activity within melanocytes. Phytother. Res. 2011, 25, 1398–1411. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Meitei, K.V. On the action and mechanism of withaferin-A from Withania somnifera, a novel and potent melanin dispersing agent in frog melanophores. J. Recept. Signal Transduct. Res. 2011, 31, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Ramdass, P.; Mullick, S.; Farber, H.F. Viral Skin Diseases. Prim. Care 2015, 42, 517–567. [Google Scholar] [CrossRef]
- Meeting, A.W. Prevention and control of herpesvirus diseases. Part 1. Clinical and laboratory diagnosis and chemotherapy. A WHO meeting. Bull. World Health Organ. 1985, 63, 185–201. [Google Scholar]
- Whitley, R.J.; David, W.K.; Roizman, B. Herpes Simplex Viruses. Clin. Infect. Dis. 1998, 26, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Wild, K.; Bohner, T.; Folkers, G.; Schulz, G.E. The structures of thymidine kinase from herpes simplex virus type 1 in complex with substrates and a substrate analogue. Protein Sci. 1997, 6, 2097–2106. [Google Scholar] [CrossRef]
- Grover, A.; Agrawal, V.; Shandilya, A.; Bisaria, V.S.; Sundar, D. Non-nucleosidic inhibition of Herpes simplex virus DNA polymerase: Mechanistic insights into the anti-herpetic mode of action of herbal drug withaferin A. BMC Bioinform. 2011, 12, S22. [Google Scholar] [CrossRef]
- Rotaru, M.; Iancu, G.; Mihalache, M.; Anton, G.; Morariu, S.-H. α-HPV positivity analysis in a group of patients with melanoma and non-melanoma skin cancers / Analiza pozitivităţii α-HPV la un grup de pacienţi cu tumori cutanate melanocitare şi non-melanocitare. Rom. Rev. Lab. Med. 2014, 22. [Google Scholar] [CrossRef]
- Cubie, H.A. Diseases associated with human papillomavirus infection. Virology 2013, 445, 21–34. [Google Scholar] [CrossRef]
- Padmavathi, B.; Rath, P.C.; Rao, A.R.; Singh, R.P. Roots of Withania somnifera Inhibit Forestomach and Skin Carcinogenesis in Mice. Evid. Based Complement Alternat. Med. 2005, 2, 99–105. [Google Scholar] [CrossRef]
- Saleem, S.; Muhammad, G.; Hussain, M.A.; Altaf, M.; Bukhari, S.N.A. Withania somnifera L.: Insights into the phytochemical profile, therapeutic potential, clinical trials, and future prospective. Iran J. Basic Med. Sci. 2020, 23, 1501–1526. [Google Scholar] [CrossRef]
- Mirjalili., M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazón, J. Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef] [PubMed]
- Withania Somnifera Root Extract. Available online: https://cosmetics.specialchem.com/inci/withania-somnifera-root-extract (accessed on 9 April 2021).
- Mishra, L.C.; Singh, B.B.; Dagenais, S. Scientific Basis for the Therapeutic Use of Withania somnifera(Ashwagandha): A Review. Altern. Med. Rev. 2000, 5, 33–38. [Google Scholar]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat. Rev. Rheumatol. 2019, 15, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Cicchese, J.M.; Evans, S.; Hult, C.; Joslyn, L.R.; Wessler, T.; Millar, J.A.; Marino, S.; Cilfone, N.A.; Mattila, J.T.; Linderman, J.J.; et al. Dynamic balance of pro- and anti-inflammatory signals controls disease and limits pathology. Immunol. Rev. 2018, 285, 147–167. [Google Scholar] [CrossRef]
- Keane, M.P.; Strieter, R.M. The importance of balanced pro-inflammatory and anti-inflammatory mechanisms in diffuse lung disease. Respir. Res. 2002, 3, 5. [Google Scholar] [CrossRef]
- Singh, N.; Bhalla, M.; de Jager, P.; Gilca, M. An overview on ashwagandha: A Rasayana (rejuvenator) of Ayurveda. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 208–213. [Google Scholar] [CrossRef]
- Imokawa, G.; Ishida, K. Inhibitors of Intracellular Signaling Pathways that Lead to Stimulated Epidermal Pigmentation: Perspective of Anti-Pigmenting Agents. Int. J. Mol. Sci. 2014, 15, 8293. [Google Scholar] [CrossRef]
- Balkrishna, A.; Nain, P.; Chauhan, A.; Sharma, N.; Gupta, A.; Ranjan, R.; Varshney, A. Super Critical Fluid Extracted Fatty Acids from Withania somnifera Seeds Repair Psoriasis-Like Skin Lesions and Attenuate Pro-Inflammatory Cytokines (TNF-α and IL-6) Release. Biomolecules 2020, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Kuchewar, V.V.; Borkar, M.A.; Nisargandha, M.A. Evaluation of antioxidant potential of Rasayana drugs in healthy human volunteers. Ayu 2014, 35, 46–49. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Active Substances | Product | Actions | ||
|---|---|---|---|---|
| WS root extract | 300 mg | Capsules | Has a general rejuvenating effect | |
| 500 mg | Antioxidant, anti-aging | |||
| WS extract (unmentioned origin) | Cream | Body | Protects the skin against pollutants and dry climate | |
| Face | Nourishing, antioxidant, and rejuvenating | |||
| Skincare | Moisturizes the skin and soothes irritated skin | |||
| Oil | Baby body | Improves skin tone, protects against dehydration and harmful external factors | ||
| Massage | Strengthens, giving vitality and increasing the resilience of the skin | |||
| Soap | Toning effect, for stressed and exhausted skin | |||
| Shower gel | Toning effect | |||
| WS extract ENERGINIUS™ | Cream | Restore skin vitality; balance fatigue generated by digital pollution (time lost in the front of monitors,) environmental pollution, etc. | ||
| Dermatological Disorder | Actions/Results of WS/WFA | Ref. |
|---|---|---|
| Carcinoma | WS roots demonstrated a 49% inhibitory effect on CHO cells colonization capacity. Cell development and adhesion are inhibited by WS, which induces long-term inhibition of CHO cell development, depending on cell density and length of Ashwagandha subjection. This information helps oncologists who intend to use Ashwagandha as synergizer, in completion to traditional therapies—radio- and chemotherapy. | [64] |
| Human malignant melanoma | WS plant extracts have anticancer properties, resulting in that fresh aqueous extract enhanced the cytotoxic effect correlated to melanoma (human malignant A375 cells). | [26] |
| Hypomelanosis | WS extract reduces endothelin-1-induced pigmentation in human epidermis by impeding the PKC action at the melanocytes level. | [43] |
| Hypopigmentation | WS and its natural chemicals (WFA and Astaxanthin) are the newest suggestions for possible anti-pigmenting substances, thus avoiding hypopigmentation risks. | [65] |
| Psoriasis Skin inflammation | WS was demonstrated to contain favorable fatty acids by the GC-FID analysis, while HPLC analysis revealed a small quantity of withanolides in WS. The WS seed * fatty acids diminished psoriatic wounds and skin inflammation in TPA-triggered, psoriatic-like mouse model. Research based on the study of TPA- or LPS-induced cells showed considerable anti-inflammatory action of WS in adjusting NFκB action and reducing the discharge of pro-inflammatory cytokines, IL-6, and TNF-α. Associating the skin reparatory and anti-inflammatory properties, WS seed fatty acids have powerful antipsoriatic action. | [66] |
| Scleroderma | It is suggested that WFA could repress the pro-inflammatory stage of fibrosis, TGF-β/Smad signaling, and considerably inhibit fibroblast conversion to myofibroblasts. Furthermore, it was revealed that the main signaling route in fibrogenesis (FoxO3a-Akt-dependent NF-κβ/IKK-mediated inflammatory response) is modulated by WFA. The results present WFA as an antifibrotic factor having favorable activity in scleroderma. | [38] |
| Skin rejuvenating agent | Ashwagandha has rejuvenating action and growth-promoting activity, both being evaluated in a 60-day study on 60 children in good health. Findings show that it can be administered as a hematinic and growth promotor in children. Medical research using root extracts of Ashwagandha shows that it presents considerable antiaging activity in healthy aged subjects. | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bungau, S.; Vesa, C.M.; Abid, A.; Behl, T.; Tit, D.M.; Purza, A.L.; Pasca, B.; Todan, L.M.; Endres, L. Withaferin A—A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases. Molecules 2021, 26, 2407. https://doi.org/10.3390/molecules26092407
Bungau S, Vesa CM, Abid A, Behl T, Tit DM, Purza AL, Pasca B, Todan LM, Endres L. Withaferin A—A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases. Molecules. 2021; 26(9):2407. https://doi.org/10.3390/molecules26092407
Chicago/Turabian StyleBungau, Simona, Cosmin Mihai Vesa, Areha Abid, Tapan Behl, Delia Mirela Tit, Anamaria Lavinia Purza, Bianca Pasca, Laura Maghiar Todan, and Laura Endres. 2021. "Withaferin A—A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases" Molecules 26, no. 9: 2407. https://doi.org/10.3390/molecules26092407
APA StyleBungau, S., Vesa, C. M., Abid, A., Behl, T., Tit, D. M., Purza, A. L., Pasca, B., Todan, L. M., & Endres, L. (2021). Withaferin A—A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases. Molecules, 26(9), 2407. https://doi.org/10.3390/molecules26092407