
The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix


 , ,
, ,  ,
,
Abstract

1. Introduction
2. Results and Discussion
2.1. Tannins Analyzed by HPLC
2.2. Study of the Effect of the Enzymes on the Cell Wall Structure and its Polysaccharides
2.2.1. Comprehensive Microarray Polymer Profiling (CoMPP) Analysis of Cell Wall Structure
2.2.2. Size Exclusion Chromatography Analysis of the Soluble Polysaccharides Released from Cell Walls
3. Materials and Methods
3.1. Grapes
3.2. Tannins Used in the Adsorption/Desorption Studies
3.3. Isolation of the Cell Wall Material as the 70% Ethanol Insoluble Residue
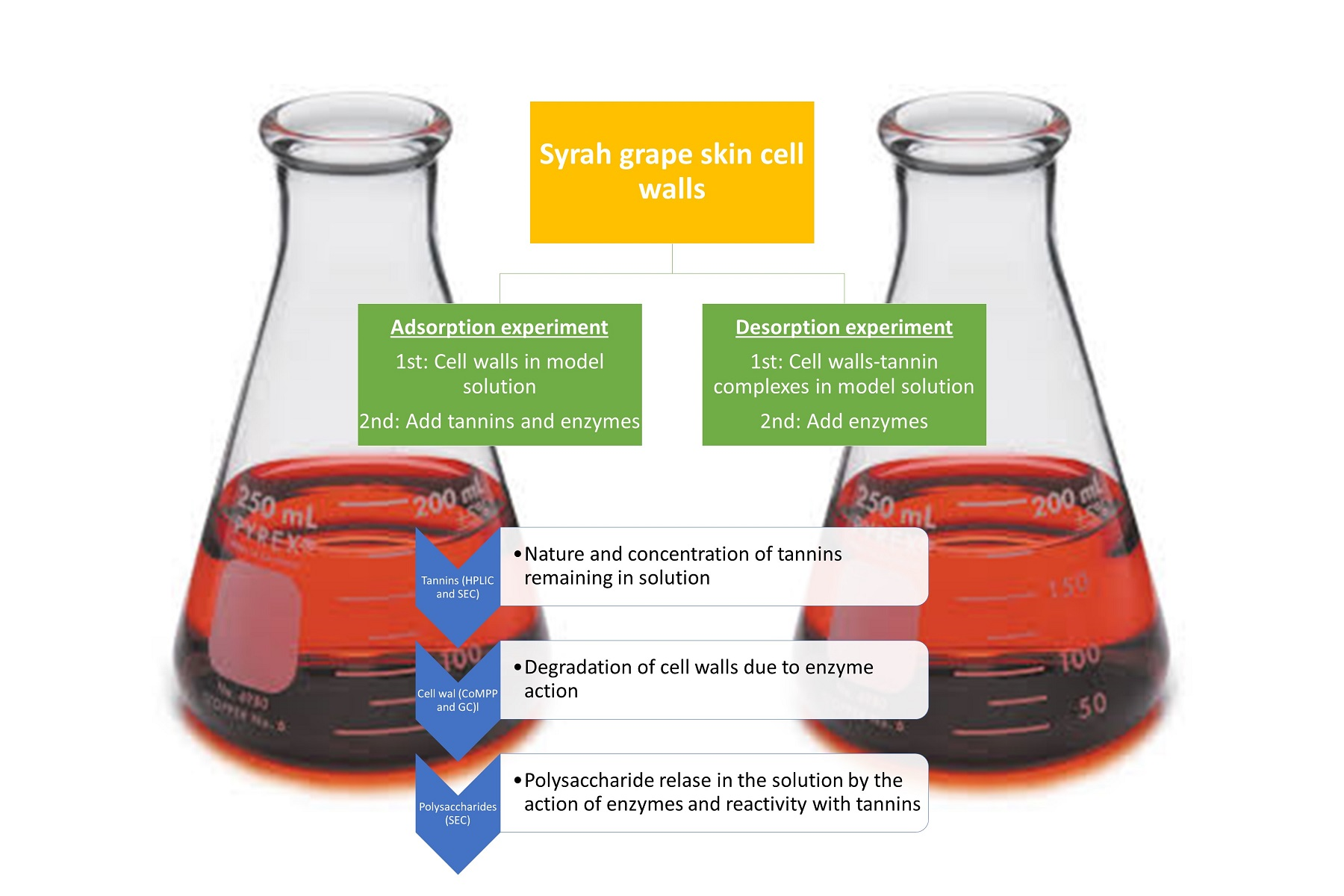
3.4. Adsorption and Desorption Experiments
3.4.1. Adsorption Test
3.4.2. Desorption Test
3.5. Analysis of Tannins Using the Phloroglucinolysis Reagent
3.6. Analysis of Soluble Polysaccharides by Size Exclusion Chromatography
3.7. Cell Wall Profiling Using Glycan Microarrays
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Monagas, M.; Bartolomé, B.; Gómez-Cordovés, C. Updated knowledge about the presence of phenolic compounds in wine. Crit. Rev. Food Sci. Nutr. 2005, 45, 85–118. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, M.D.; Dambergs, R.G.; Cozzolino, D.; Herderich, M.J.; Smith, P.A. Relationship between red wine grades and phenolics. 1. Tannin and total phenolics concentrations. J. Agric. Food Chem. 2010, 58, 12313–12319. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O.; Scholz, R.C. Tannins–the problem of extraction. In Proceedings of the 13th Australian Wine Industry Technical Conference, Adeline, Australia, 28 July–2 August 2007; The Australian Wine Industry Technical Conference Inc.: Adeline, Australia, 2007; Volume 28, pp. 160–164. [Google Scholar]
- Busse-Valverde, N.; Gómez-Plaza, E.; López-Roca, J.M.; Gil-Muññoz, R.; Fernández-Fernández, J.I.; Bautista-Ortín, A.B. Effect of different enological practices on skin and seed proanthocyanidins in three varietal wines. J. Agric. Food Chem. 2010, 58, 11333–11339. [Google Scholar] [CrossRef] [PubMed]
- Romero-Cascales, I.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. The effect of a commercial pectolytic enzyme on grape skin cell wall degradation and colour evolution during the maceration process. Food Chem. 2012, 130, 626–631. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.B.; Busse-Valverde, N.; Fernández-Fernández, J.I.; Gómez-Plaza, E.; Gil-Muñoz, R. The extraction kinetics of anthocyanins and proanthocyanidins from grape to wine in three different varieties. OENO One 2016, 50, 91–100. [Google Scholar] [CrossRef]
- Bindon, K.A.; Smith, P.A.; Kennedy, J.A. Interaction between grape-derived proanthocyanidins and cell wall material. 1. Effect on proanthocyanidin composition and molecular mass. J. Agric. Food Chem. 2010, 58, 2520–2528. [Google Scholar] [CrossRef]
- Castro-López, L.; Gómez-Plaza, E.; Ortega-Regules, A.; Lozada, D.; Bautista-Ortín, A.B. Role of cell wall deconstructing enzymes in the proanthocyanidin–cell wall adsorption–desorption phenomena. Food Chem. 2016, 196, 526–532. [Google Scholar] [CrossRef]
- Osete-Alcaraz, A.; Bautista-Ortín, A.B.; Ortega-Regules, A.; Gómez-Plaza, E. Elimination of suspended cell wall material in musts improves the phenolic content and color of red wines. Am. J. Enol. Vitic. 2019, 70, 201–204. [Google Scholar] [CrossRef]
- Romero-Cascales, I.; Fernández-Fernández, J.I.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. Characterisation of the main enzymatic activities present in six commercial macerating enzymes and their effects on extracting colour during winemaking of Monastrell grapes. J. Food Sci. Technol. 2008, 43, 1295–1305. [Google Scholar] [CrossRef]
- Godfrey, T.; Reichelt, J. Industrial Enzymology: The Application of Enzymes in Industry, 1st ed.; Palgrave Macmillan: London, UK, 1982; pp. 410–421. [Google Scholar]
- Bautista-Ortín, A.; Ben Abdallah, R.; Castro-López, L.; Jiménez-Martínez, M.D.; Gómez-Plaza, E. Technological implications of modifying the extent of cell wall-proanthocyanidin interactions using enzymes. Int. J. Mol. Sci. 2016, 17, 123. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Smith, P.A.; Bindon, K.A. Selective extraction of polysaccharide affects the adsorption of proanthocyanidin by grape cell walls. Carbohydr. Polym. 2014, 114, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Renard, C.M.; Watrelot, A.A.; Le Bourvellec, C. Interactions between polyphenols and polysaccharides: Mechanisms and consequences in food processing and digestion. Trends Food Sci. Technol. 2017, 60, 43–51. [Google Scholar] [CrossRef]
- Beaver, J.W.; Miller, K.V.; Medina-Plaza, C.; Dokoozlian, N.; Ponangi, R.; Blair, T.; Block, D.; Oberholster, A. Heat-Dependent Desorption of Proanthocyanidins from Grape-Derived Cell Wall Material under Variable Ethanol Concentrations in Model Wine Systems. Molecules 2019, 24, 3561. [Google Scholar] [CrossRef] [PubMed]
- Osete-Alcaraz, A.; Gómez-Plaza, E.; Martínez-Pérez, P.; Weiller, F.; Schückel, J.; Willats, W.G.T.; Moore, J.P.; Ros-García, J.M.; Bautista-Ortín, A.B. The impact of carbohydrate-active enzymes on mediating cell wall polysaccharide-tannin interactions in a wine-like matrix. Int. Food Res. J. 2020, 129, 108889. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Covington, A.; Hancock, R. Structure-activity relationships in the hydrophobic interactions of polyphenols with cellulose and collagen. Biopolymers 2003, 70, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Medina-Plaza, C.; Beaver, J.W.; Lerno, L.; Dokoozlian, N.; Ponangi, R.; Blair, T.; Block, D.; Oberholster, A. Impact of temperature, ethanol and cell wall material composition on cell wall-anthocyanin interactions. Molecules 2019, 24, 3350. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Bouchet, B.; Renard, C.M.G.C. Non-covalent interaction between procyanidins and apple cell wall material. Part III: Study on model polysaccharides. Biochim. Biophys. Acta Gen. Subj. 2005, 1725, 10–18. [Google Scholar] [CrossRef]
- Le Bourvellec, C.L.; Guyot, S.; Renard, C.M.G.C. Interactions between apple (Malus× domestica Borkh.) polyphenols and cell walls modulate the extractability of polysaccharides. Carbohydr. Polym. 2009, 75, 251–261. [Google Scholar] [CrossRef]
- Watrelot, A.A.; Le Bourvellec, C.; Imberty, A.; Renard, C.M. Interactions between pectic compounds and procyanidins are influenced by methylation degree and chain length. Biomacromolecules 2013, 14, 709–718. [Google Scholar] [CrossRef]
- Watrelot, A.A.; Le Bourvellec, C.; Imberty, A.; Renard, C.M. Neutral sugar side chains of pectins limit interactions with procyanidins. Carbohydr. Polym. 2014, 99, 527–536. [Google Scholar] [CrossRef]
- Gao, Y.; Fangel, J.U.; Willats, W.G.; Vivier, M.A.; Moore, J.P. Dissecting the polysaccharide-rich grape cell wall matrix using recombinant pectinases during winemaking. Carbohydr. Polym. 2016, 152, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Regules, A.; Ros-García, J.M.; Bautista-Ortín, A.B.; López-Roca, J.M.; Gómez-Plaza, E. Differences in morphology and composition of skin and pulp cell walls from grapes (Vitis vinifera L.): Technological implications. Eur. Food Res. Technol. 2008, 227, 223. [Google Scholar] [CrossRef]
- Gao, Y.; Zietsman, J.J.; Vivier, M.; Moore, J. Deconstructing wine grape cell walls with enzymes during winemaking: New insights from glycan microarray technology. Molecules 2019, 24, 165. [Google Scholar] [CrossRef] [PubMed]
- Zietsman, A.J.; Moore, J.P.; Fangel, J.U.; Willats, W.G.; Vivier, M.A. Profiling the hydrolysis of isolated grape berry skin cell walls by purified enzymes. J. Agric. Food Chem. 2015, 63, 8267–8274. [Google Scholar] [CrossRef] [PubMed]
- Zietsman, A.J.; Moore, J.P.; Fangel, J.U.; Willats, W.G.; Trygg, J.; Vivier, M.A. Following the compositional changes of fresh grape skin cell walls during the fermentation process in the presence and absence of maceration enzymes. J. Agric. Food Chem. 2015, 63, 2798–2810. [Google Scholar] [CrossRef]
- Renard, C.M.C.G.; Baron, A.; Guyot, S.; Drilleau, J.F. Interactions between apple cell walls and native apple polyphenols: Quantification and some consequences. Int. J. Biol. Macromol. 2001, 29, 115–125. [Google Scholar] [CrossRef]
- Bindon, K.; Kassandra, S.; Smith, P. Retention of proanthocyanidin in wine like solution is conferred by a dynamic interaction between soluble and insoluble grape cell wall components. J. Agric. Food Chem. 2016, 64, 8406–8419. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P. Analysis of proanthocyanidin cleavage products following acid-catalysis in the presence of excess phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef]
- Smith, P.A. Precipitation of tannin with methyl cellulose allows tannin quantification in grape and wine samples. Tech. Rev. ARWI 2005, 158, 3–7. [Google Scholar]
- De Vries, J.A.; Voragen, A.G.J.; Rombouts, F.M.; Pilnik, W. Extraction and purification of pectins from alcohol insoluble solids from ripe and unripe apples. Carbohydr. Polym. 1981, 1, 117–127. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Moller, I.; Sørensen, I.; Bernal, A.J.; Blaukopf, C.; Lee, K.; Øbro, J.; Pettolino, F.; Roberts, A.; Mikkelsen, J.D.; Knox, J.P.; et al. High-throughput mapping of cell-wall polymers within and between plants using novel microarrays. Plant. J. 2007, 50, 1118–1128. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Samples | TT (mg/L) | % Adsorption | mDP | %Gal |
|---|---|---|---|---|
| Tannin | 1788.4 c * | 2.70 b | 14.5 d | |
| CW + Tan | 1040.3 a | 41.8 | 2.49 a | 12.3 a |
| CW + Tan + CEL | 1274.5 b | 28.7 | 2.48 a | 12.8 bc |
| CW + Tan + PME | 1179.2 ab | 34.1 | 2.49 a | 12.6 b |
| CW + Tan + PEC | 1272.1 b | 28.9 | 2.47 a | 12.9 c |
| CW + Tan + XYL | 1278.4 b | 28.5 | 2.47 a | 12.8 bc |
| Samples | TT (mg/L) | % Released | mDP | %Gal |
|---|---|---|---|---|
| CW-Tan | 186.0 a * | 24.9 | 3.05 ab | 15.3 ab |
| CW-Tan + CEL | 185.6 a | 24.8 | 3.00 ab | 14.9 a |
| CW-Tan + PME | 227.4 b | 30.4 | 2.99 a | 14.9 a |
| CW-Tan + PEC | 166.7 a | 22.3 | 3.24 c | 16.3 c |
| CW-Tan + XYL | 219.7 b | 29.4 | 3.09 b | 15.4 b |
| Fractions | Samples | HG partially/de-esterifies (mAb JIM5) | HG partially esterifies (mAb JIM7) | HG partially/de-esterifies (mAb LM8) | HG partially/de-esterifies (mAb LM19) | HG partially esterifies (mAb LM20) | HG Ca2+ crosslinked (mAb 2F4) | Xylogalacturonan (mAb LM8) | Backbone of RG I (mAb INRA-RU1) | Backbone of RG I (mAb INRA-RU2) | (1→4)-β-D-galactan (mAb LM5) | Feruloylated (1→4)-β-D-galactan (mAb LM9) | Linearised (1→5)-α-L-arabinan (mAb LM13) | (1→4)-β-D-mannan (mAb LM21) | (1→4)-β-D-mannan/galactomannan (mAb LM22) | (1→3)-β-D-glucan (mAb BS-400-2) | Xyloglucan (XXXG motif) (mAb LM15) | Xyloglucan (mAb LM25) | (1→4)-β-D-xylan (mAb LM10) | (1→4)-β-D-xylan/arabinoxylan (mAb LM11) | Cellulose (Cristalline < 9 (mAb CBM3a) | Extensin (mAb LM1) | Extensin (mAb JIM11) | Extensin (mAb JIM20) | AGP (mAb JIM8) | AGP (mAb JIM13) | AGP (mAb JIM14) | AGP, β-linked GlcA (mAb LM2) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CDTA | CW’ | 31 | 49 | 9 | 0 | 47 | 0 | 0 | 10 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| CW | 43 | 59 | 15 | 8 | 51 | 0 | 0 | 17 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan | 46 | 65 | 18 | 10 | 64 | 0 | 0 | 19 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + CEL | 36 | 47 | 14 | 7 | 42 | 0 | 0 | 16 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + PME | 58 | 73 | 21 | 9 | 66 | 0 | 0 | 20 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + PEC | 37 | 26 | 20 | 7 | 13 | 0 | 0 | 12 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + XYL | 39 | 63 | 15 | 8 | 56 | 0 | 0 | 16 | 22 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| NaOH | CW’ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 90 | 87 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| CW | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 9 | 6 | 0 | 0 | 2 | 0 | 0 | 81 | 81 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 84 | 84 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + CEL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 8 | 2 | 0 | 0 | 0 | 0 | 0 | 80 | 80 | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + PME | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 83 | 81 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + PEC | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 22 | 8 | 0 | 0 | 0 | 0 | 0 | 79 | 77 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 0 | 0 | |
| CW + Tan + XYL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 76 | 74 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 |
| Fractions | Samples | HG partially/de-esterifies (mAb JIM5) | HG partially esterifies (mAb JIM7) | HG partially/de-esterifies (mAb LM8) | HG partially/de-esterifies (mAb LM19) | HG partially esterifies (mAb LM20) | HG Ca2+ crosslinked (mAb 2F4) | Xylogalacturonan (mAb LM8) | Backbone of RG I (mAb INRA-RU1) | Backbone of RG I (mAb INRA-RU2) | (1→4)-β-D-galactan (mAb LM5) | Feruloylated (1→4)-β-D-galactan (mAb LM9) | Linearised (1→5)-α-L-arabinan (mAb LM13) | (1→4)-β-D-mannan (mAb LM21) | (1→4)-β-D-mannan/galactomannan (mAb LM22) | (1→3)-β-D-glucan (mAb BS-400-2) | Xyloglucan (XXXG motif) (mAb LM15) | Xyloglucan (mAb LM25) | (1→4)-β-D-xylan (mAb LM10) | (1→4)-β-D-xylan/arabinoxylan (mAb LM11) | Cellulose (Cristalline < 9 (mAb CBM3a) | Extensin (mAb LM1) | Extensin (mAb JIM11) | Extensin (mAb JIM20) | AGP (mAb JIM8) | AGP (mAb JIM13) | AGP (mAb JIM14) | AGP, β-linked GlcA (mAb LM2) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CDTA | CW’’ | 31 | 54 | 11 | 0 | 46 | 0 | 0 | 12 | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| CW-Tan | 36 | 56 | 11 | 0 | 49 | 0 | 0 | 11 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + CEL | 27 | 46 | 8 | 0 | 37 | 0 | 0 | 10 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + PME | 40 | 58 | 13 | 0 | 53 | 0 | 0 | 14 | 22 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + PEC | 37 | 20 | 13 | 0 | 4 | 0 | 0 | 7 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + XYL | 40 | 51 | 11 | 0 | 47 | 0 | 0 | 15 | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| NaOH | CW’’ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 76 | 71 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 |
| CW-Tan | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 77 | 76 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + CEL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 70 | 69 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + PME | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 87 | 84 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + PEC | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 87 | 82 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | |
| CW-Tan + XYL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 85 | 82 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Samples | Area (12–22.5 min) | Area (12–18 min) | Area (18.01–22.5 min) |
|---|---|---|---|
| CW | 75.04 | 53.91 | 21.03 |
| Tan | 18.51 | −5.12 | 23.64 |
| CW + Tan | 79.30 | 39.53 | 39.68 |
| CW + CEL | 168.93 | 75.08 | 93.53 |
| CW + Tan + CEL | 164.65 | 56.92 | 107.61 |
| CW + PME | 91.50 | 63.37 | 27.93 |
| CW + Tan + PME | 87.41 | 41.44 | 45.87 |
| CW + PEC | 675.30 | 59.24 | 614.49 |
| CW + Tan + PEC | 461.69 | 17.65 | 443.09 |
| CW + XYL | 91.87 | 69.66 | 22.03 |
| CW + Tan + XYL | 92.71 | 44.50 | 48.11 |
| Samples | Area (12–22.5 min) | Area (12–18 min) | Area (18.01–22.5 min) |
|---|---|---|---|
| CW-Tan | 13.91 | 11.78 | 2.11 |
| CW’’ + CEL | 99.07 | 29.24 | 69.76 |
| CW-Tan + CEL | 67.39 | 13.00 | 54.36 |
| CW’’ + PME | 19.15 | 13.63 | 5.50 |
| CW-Tan + PME | 13.23 | 5.04 | 8.19 |
| CW’’ + PEC | 450.60 | 31.10 | 418.39 |
| CW-Tan + PEC | 337.09 | 6.06 | 330.45 |
| CW’’ + XYL | 17.15 | 14.20 | 2.93 |
| CW-Tan + XYL | 16.44 | 8.77 | 7.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osete-Alcaraz, A.; Gómez-Plaza, E.; Martínez-Pérez, P.; Weiller, F.; Schückel, J.; Willats, W.G.T.; Moore, J.P.; Ros-García, J.M.; Bautista-Ortín, A.B. The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix. Molecules 2021, 26, 770. https://doi.org/10.3390/molecules26030770
Osete-Alcaraz A, Gómez-Plaza E, Martínez-Pérez P, Weiller F, Schückel J, Willats WGT, Moore JP, Ros-García JM, Bautista-Ortín AB. The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix. Molecules. 2021; 26(3):770. https://doi.org/10.3390/molecules26030770
Chicago/Turabian StyleOsete-Alcaraz, Andrea, Encarna Gómez-Plaza, Pilar Martínez-Pérez, Florent Weiller, Julia Schückel, William G.T. Willats, John P. Moore, José M. Ros-García, and Ana B. Bautista-Ortín. 2021. "The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix" Molecules 26, no. 3: 770. https://doi.org/10.3390/molecules26030770
APA StyleOsete-Alcaraz, A., Gómez-Plaza, E., Martínez-Pérez, P., Weiller, F., Schückel, J., Willats, W. G. T., Moore, J. P., Ros-García, J. M., & Bautista-Ortín, A. B. (2021). The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix. Molecules, 26(3), 770. https://doi.org/10.3390/molecules26030770