
Glyceryl Trinitrate: History, Mystery, and Alcohol Intolerance


Abstract
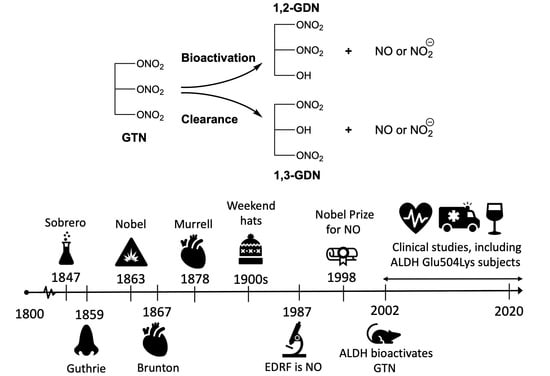
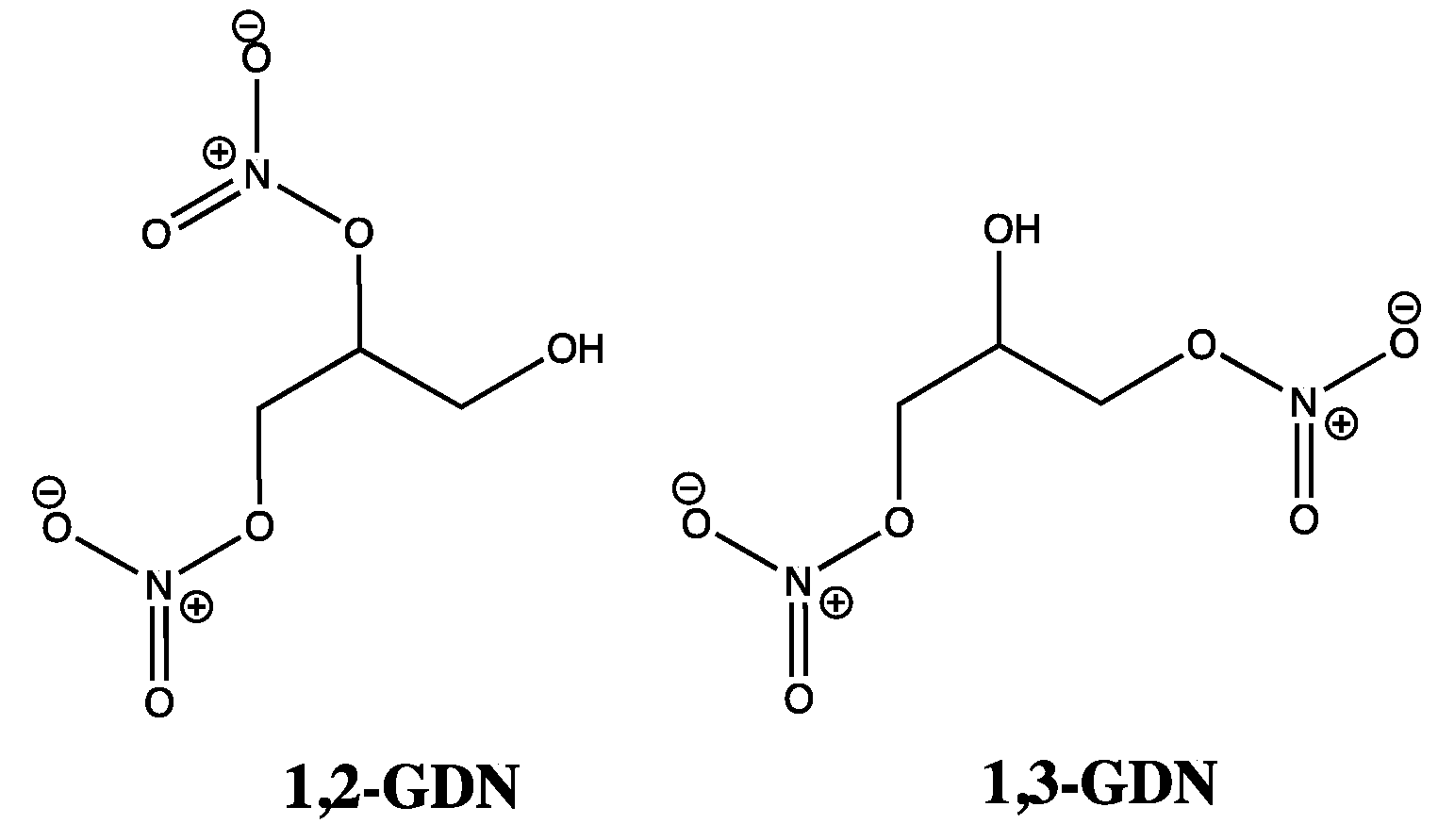
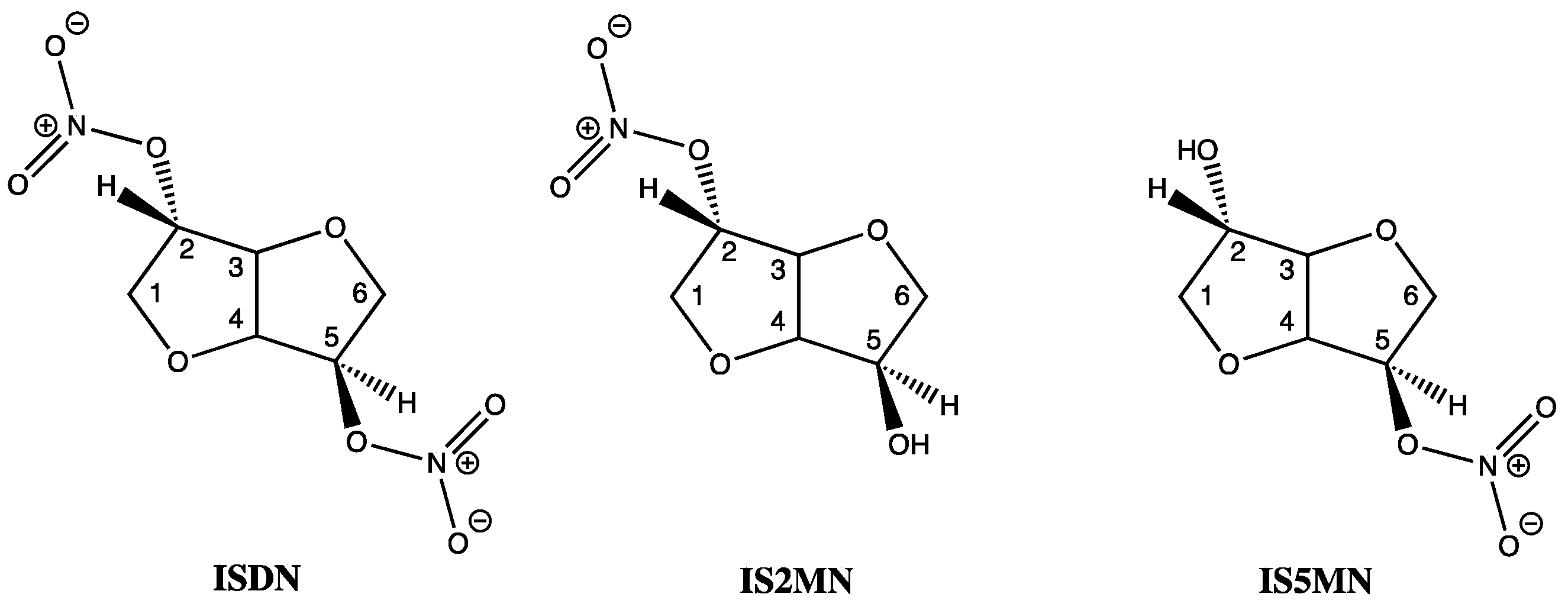
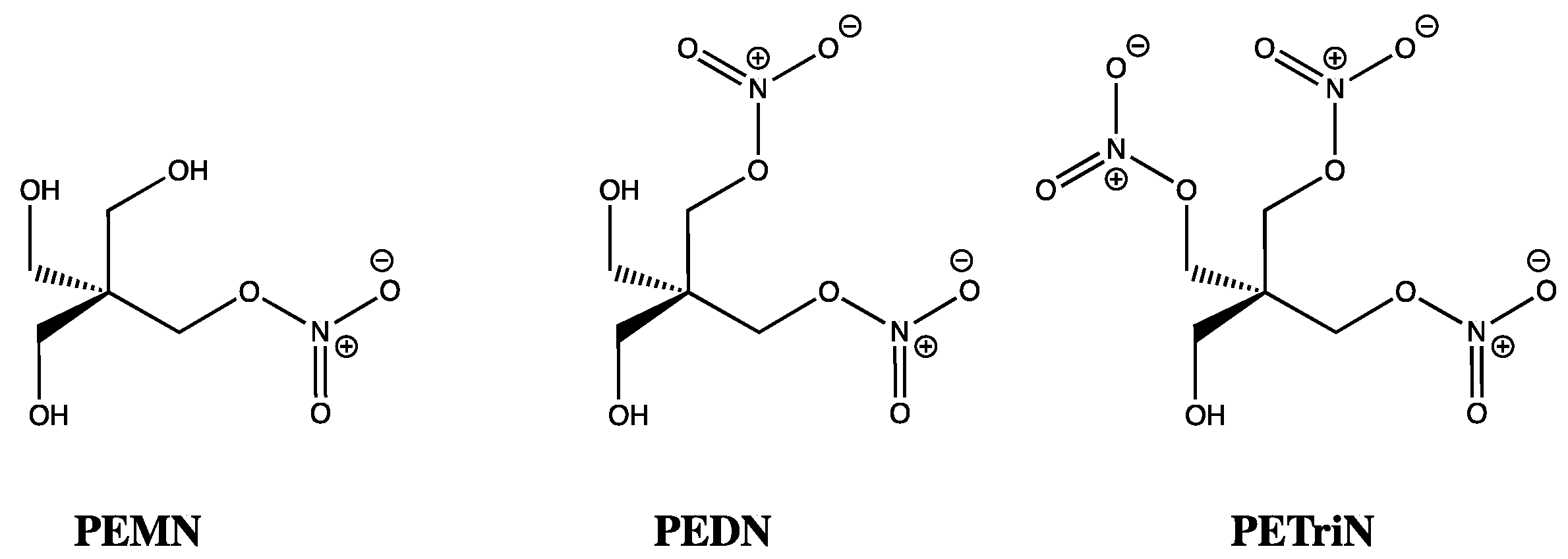
:
1. Introduction
2. Invention of Glyceryl Trinitrate
3. Dynamite
4. Biological Action of GTN
5. GTN as a Muscle Relaxant
6. The Coming of GTN
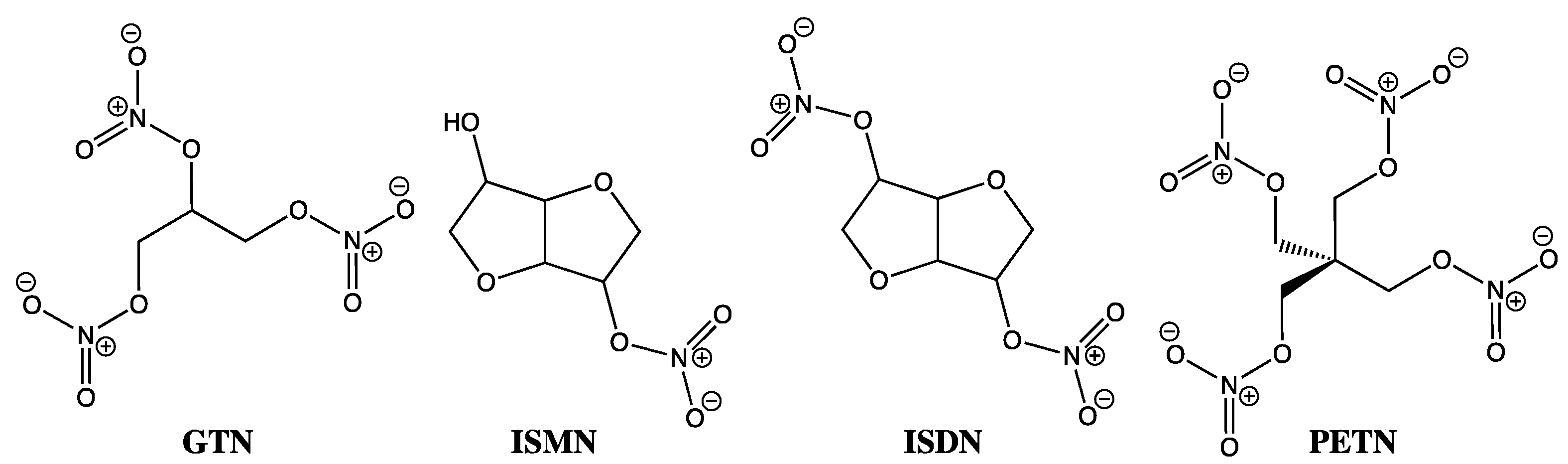
7. The GTN Conundrum
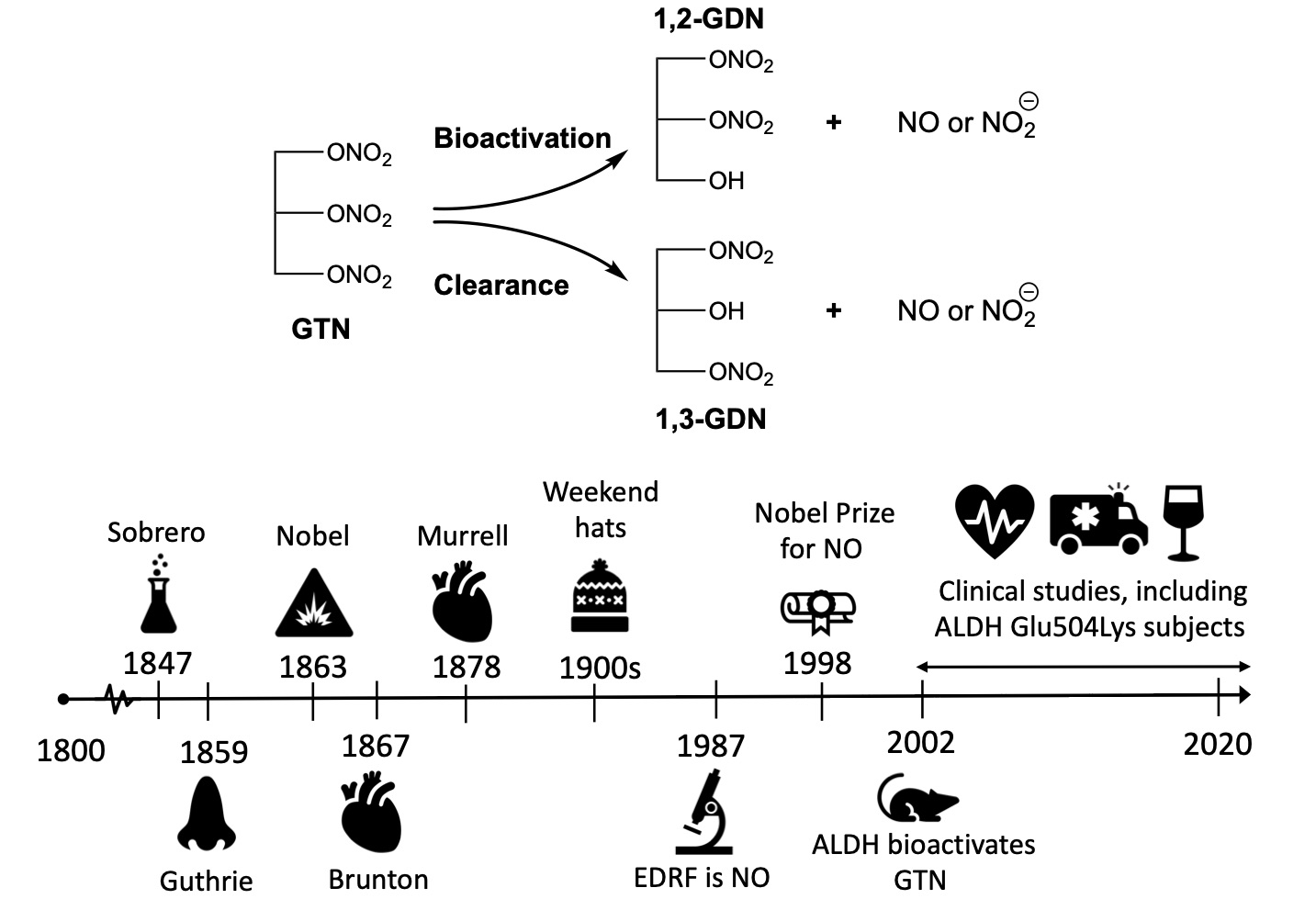
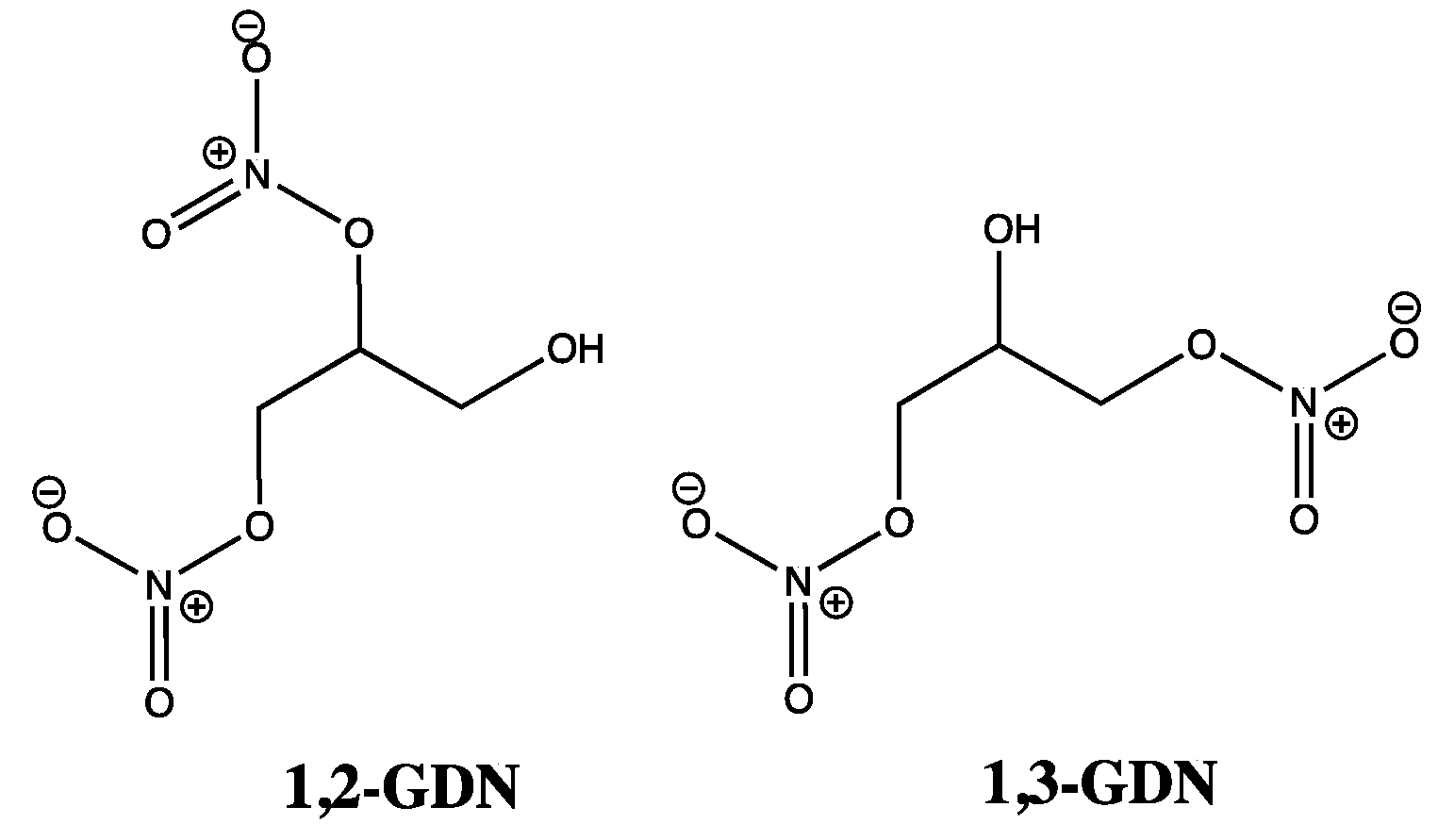
8. GTN Clearance versus Bioactivation
9. Non-Enzymatic Reactions
10. Mechanistic Evidence from Observations Outside of the Lab
11. Aldehyde Dehydrogenase (ALDH)
12. The Use of ALDH Inhibitors and ALDH Knockout Models
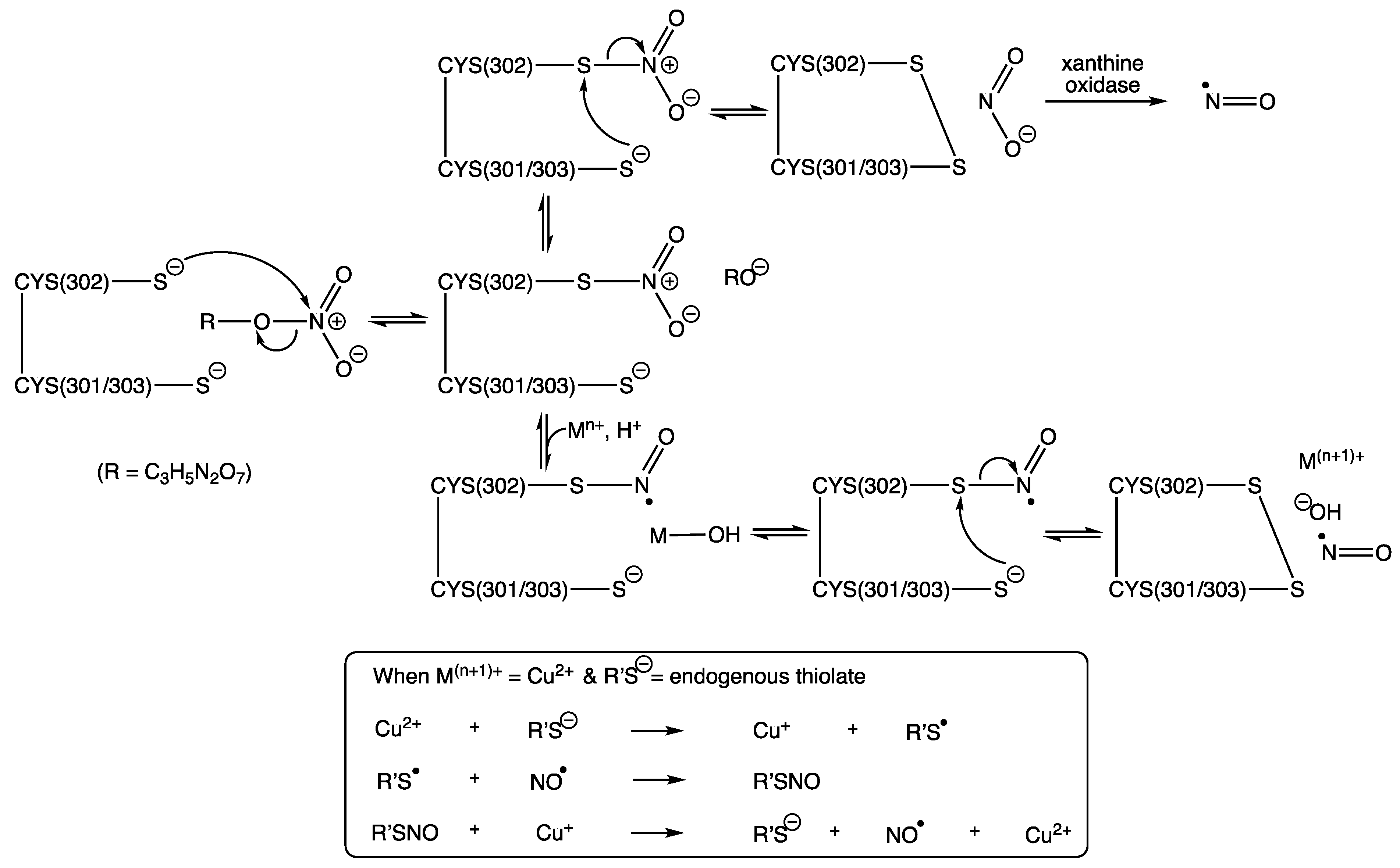
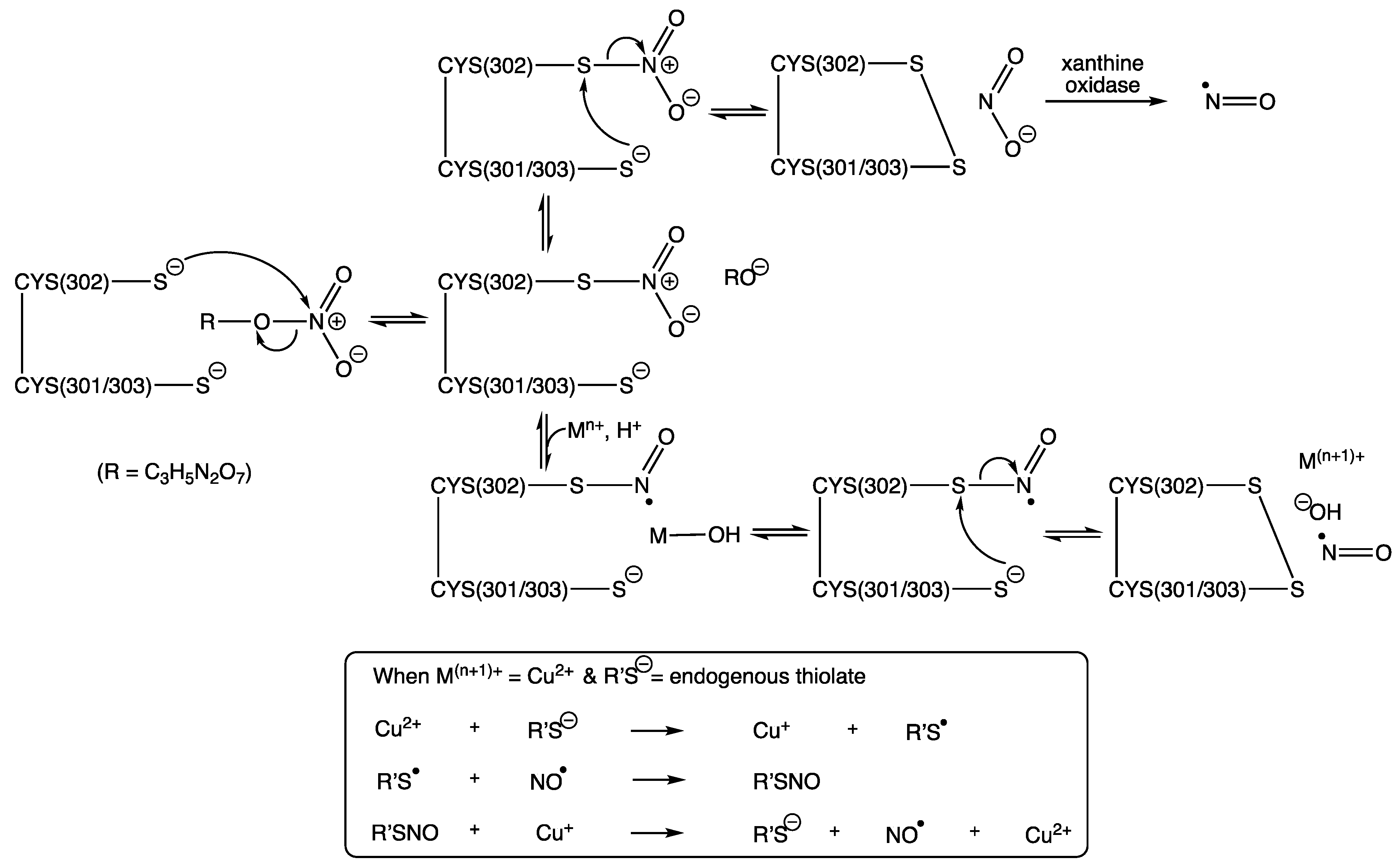
13. The Active Site of Mitochondrial Aldehyde Dehydrogenase
14. Nitrate Tolerance
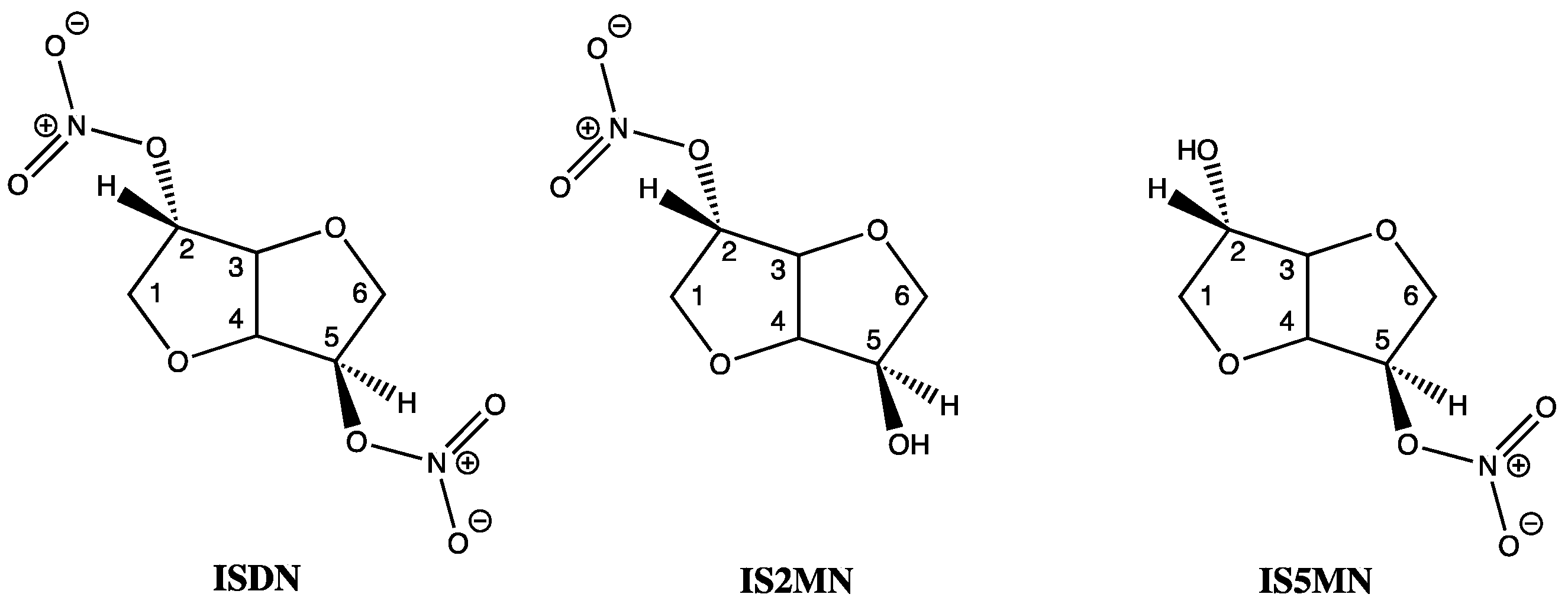
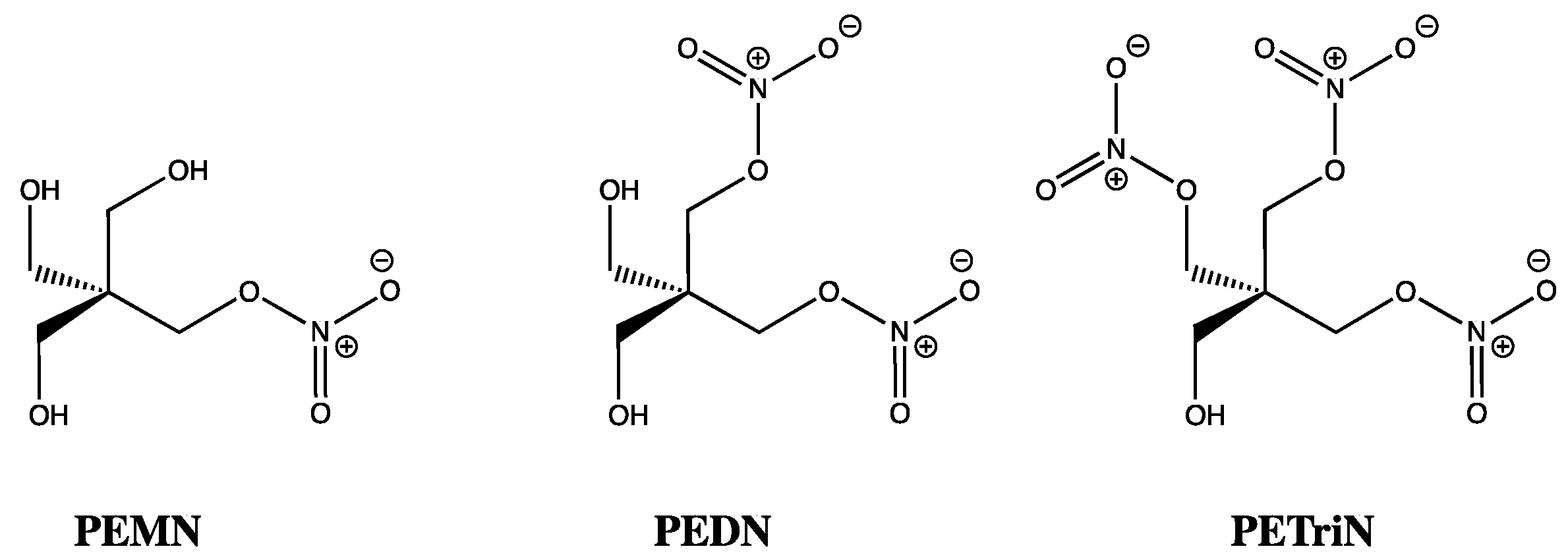
15. GTN in Individuals with Lowered Alcohol Tolerance
16. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Palmer, R.M.J.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci. USA 1987, 84, 9265–9269. [Google Scholar] [CrossRef] [Green Version]
- Furchgott, R.F.; Zawadzi, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Murad, F.; Arnold, W.P.; Mittal, C.K.; Braughler, J.M. Properties and regulation of guanylate cyclase and some proposed functions for cyclic GMP. Adv. Cyclic Nucleotide Res. 1979, 11, 175–204. [Google Scholar]
- Thatcher, G.R.J.; Nicolescu, A.C.; Bennett, B.M.; Toader, V. Nitrates and NO release: Contemporary aspects in biological and medicinal chemistry. Free Rad. Biol. Med. 2004, 37, 1122–1143. [Google Scholar] [CrossRef] [PubMed]
- McAninly, J.; Williams, L.H.; Askew, S.C.; Butler, A.R.; Russell, C. Metal ion catalysis in nitrosothiol (RSNO) decomposition. J. Chem. Soc. Chem. Comm. 1993, 1758–1759. [Google Scholar] [CrossRef]
- Fant, K. Alfred Nobel: A Biography; Arcade Publishing: New York, NY, USA, 1993. [Google Scholar]
- Guthrie, F. Contribution to the knowledge of the amyl group. Q. J. Chem. Soc. 1859, 11, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Richardson, B.W. Report on the physiological action of nitrite of amyl. Rep. Brit. Assoc. Adv. Sci. 1864, 34, 120–129. [Google Scholar]
- Marsh, N.; Marsh, A. A short history of nitroglycerine and nitric oxide in pharmacology and physiology. Clin. Exp. Pharmacol. Physiol. 2000, 27, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Molina, C.R.; Andresen, J.W.; Rapoport, R.M.; Waldman, S.; Murad, F. Effect of in vivo nitroglycerin therapy on endothelium-dependent and independent vascular relaxation and cyclic GMP accumulation in rat aorta. J. Cardiovasc. Pharmacol. 1987, 10, 371–378. [Google Scholar] [CrossRef]
- Ignarro, L.J. Biological actions and properties of endothelium-derived nitric oxide formed and released from artery and vein. Circ. Res. 1989, 65, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Mayer, B.; Beretta, M. The enigma of nitroglycerin bioactivation and nitrate tolerance: News, views and troubles. Brit. J. Pharmacol. 2008, 155, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Fung, H.-L. Biochemical mechanism of nitroglycerin action and tolerance: Is this old mystery solved? Annu. Rev. Pharmacol. Toxicol. 2004, 44, 67–85. [Google Scholar] [CrossRef]
- Hashimoto, S.; Kobayashi, A. Clinical pharmacokinetics and pharmacodynamics of glyceryl trinitrate and its metabolites. Clin. Pharmacokinet. 2003, 42, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Jung, P.; Sanders, S.W.; Lin, E.T.; Benet, L.Z. Pharmacokinetics of nitroglycerin and its four metabolites during nitroglycerin transdermal administration. Biopharm. Drug Dispos. 1994, 15, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Christodoulatos, C.; Bhaumik, S.; Brodman, B.W. Anaerobic biodegradation of nitroglycerin. Wat. Res. 1997, 31, 1462–1470. [Google Scholar] [CrossRef]
- Needleman, P.; Jakschik, B.; Johnson, E.M., Jr. Sulfhydryl requirement for relaxation of vascular smooth muscle. J. Pharmacol. Exper. Therapeut. 1973, 187, 324–331. [Google Scholar]
- Ignarro, L.J.; Gruetter, C.S. Requirement of thiols for activation of coronary arterial guanylate cyclase by glyceryl trinitrate and sodium nitrite: Possible involvement of S-nitrosothiols. Biochim. Biophys. Acta 1980, 631, 221–231. [Google Scholar] [CrossRef]
- Ignarro, L.J.; Lippton, H.; Edwards, J.C.; Baricos, W.H.; Hyman, A.L.; Kadowitz, P.J.; Gruetter, C.A. Mechanism of vascular smooth muscle relaxation by organic nitrates, nitrites, nitroprusside and nitric oxide: Evidence for the involvement of S-nitrosothiols as active intermediates. J. Pharmacol. Exper. Therapeut. 1981, 218, 739–749. [Google Scholar]
- Mukerjee, N.; Pietruszko, R. Inactivation of human aldehyde dehydrogenase by isosorbide dinitrate. J. Biol. Chem. 1994, 269, 21664–21669. [Google Scholar] [CrossRef]
- Pietruszko, R.; Mukerjee, N.; Blatter, E.E.; Lehmann, T. Nitrate esters as inhibitors and substrates of aldehyde dehydrogenase. In Enzymology and Molecular Biology of Carbonyl Metabolism 5; Weiner, H., Holmes, R.S., Wermuth, B., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 25–34. [Google Scholar]
- Yeates, R.A.; Laufen, H.; Leitold, M. The reaction between organic nitrates and sulhydryl compounds. A possible model system for the activation of organic nitrates. Mol. Pharmacol. 1985, 28, 555–559. [Google Scholar]
- Butler, A.R.; Ridd, J.H. Formation of nitric oxide from nitrous acid in ischemic tissue and skin. Nitric Oxide 2004, 10, 20–24. [Google Scholar] [CrossRef]
- Dicks, A.P.; Swift, H.R.; Williams, D.L.H.; Butler, A.R.; Al-Sa’doni, H.H.; Cox, B.G. Identification of Cu+ as the effective reagent in nitric oxide formation from S-nitrosothiols (RSNO). J. Chem. Soc. Perkin Trans. 2 1996, 481–487. [Google Scholar] [CrossRef]
- Mirvish, S.S.; Wallcave, L.; Eagen, M.; Shubik, P. Ascorbate-nitrite reaction: Possible means of blocking the formation of carcinogenic N-nitroso compounds. Science 1972, 177, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Archer, M.C.; Tannenbaum, S.R.; Fan, T.-Y.; Weisman, M. Reaction of nitrite with ascorbate and its relation to nitrosamine formation. J. Nation. Cancer Instit. 1975, 54, 1203–1205. [Google Scholar] [CrossRef]
- Kollau, A.; Baretta, M.; Gorren, A.C.F.; Russwurm, M.; Koesling, D.; Schmidt, K.; Mayer, B. Bioactivation of nitroglycerin by ascorbate. Mol. Pharmacol. 2007, 72, 191–196. [Google Scholar] [CrossRef]
- Feelisch, M.; Noack, E.A. Correlation between nitric oxide formation during degradation of organic nitrates and activation of guanylate cyclase. Eur. J. Pharmacol. 1987, 139, 19–30. [Google Scholar] [CrossRef]
- Bennett, B.M.; Kobus, S.M.; Brien, J.F.; Nakatsu, K.; Marks, G.S. Requirement for reduced, unliganded hemoprotein for the hemoglobin- and myoglobin-mediated biotransformation of glyceryl trinitrate. J. Pharmacol. Exp. Therapeut. 1986, 237, 629–635. [Google Scholar]
- Ignarro, L.J. After 130 years, the molecular mechanism of action of nitroglycerin is revealed. Proc. Nat. Acad. Sci. USA 2002, 99, 7816–7817. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.R. The heart less bounding: Treating angina pectoris. J. R. Coll. Physians Edinb. 2006, 36, 185–189. [Google Scholar]
- Nash, G. GTN headaches and reduction of natural banding? Colorectal Dis. 2008, 10, 96. [Google Scholar] [CrossRef]
- Tjandra, J.J.; Tan, J.J.F.; Lim, J.F.; Murray-Green, C.; Kennedy, M.L.; Lubowski, D.Z. Rectogesic (glyceryl trinitrate 0.2%) ointment relieved symptoms of haemorrhoids associated with high resting anal canal pressures. Colorectal Dis. 2007, 9, 457–463. [Google Scholar] [CrossRef]
- Loder, P.B.; Kamm, M.A.; Nicholls, R.J.; Phillips, R.K.S. Reversible chemical sphincterotomy by local application of glyceryl trinitrate. Brit. J. Surg. 1994, 9, 1386–1389. [Google Scholar] [CrossRef] [PubMed]
- Dipalma, J.F. The nitrites and nitrates. Am. Fam. Physician 1982, 25, 216–218. [Google Scholar] [PubMed]
- Chen, Z.; Zhang, J.; Stamler, J.S. Identification of the enzymatic mechanism of nitroglycerin bioactivation. Proc. Nat. Acad. Sci. USA 2002, 99, 8306–8311. [Google Scholar] [CrossRef] [Green Version]
- Murad, F.; Mittal, C.K.; Arnold, W.P.; Katsuki, S.; Kimura, H. Guanylate cyclase: Activation by azide, nitro compounds, nitric oxide, and hydroxyl radical and inhibition by hemoglobin and myoglobin. Adv. Cyclic Nucleotide Res. 1978, 9, 145–158. [Google Scholar]
- Needleman, P.; Hunter, F.E. The transformation of glyceryl trinitrate and other nitrates by glutathione-organic nitrate reductase. Mol. Pharmacol. 1965, 1, 77–86. [Google Scholar]
- Tsuchida, S.; Maki, T.; Sato, K. Purification and characterization of glutathione transferases with an activity toward nitroglycerin from human aorta and heart. J. Biol. Chem. 1990, 265, 7150–7157. [Google Scholar] [CrossRef]
- Nigam, R.; Anderson, D.J.; Lee, S.F.; Bennett, B.M. Isoform-specific biotransformation of glyceryl trinitrate by rat aortic glutathione S-transferases. J. Pharmacol. Exper. Therapeut. 1996, 279, 1527–1534. [Google Scholar]
- Ji, Y.; Anderson, D.J.; Bennett, B.M. Role of microsomal glutathione transferase 1 in the mechanism-based biotransformation of glyceryl trinitrate in LLC-PK1 cells. Biochem. Pharmacol. 2009, 77, 1702–1708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-J.; Chang, L.; Zhang, Y.-M.; Deng, S.; Li, Y.-J.; Peng, J. Comparing the role of glutathione-S-transferase and mitochondrial aldehyde dehydrogenase in nitroglycerin biotransformation and the correlation with calcitonin gene-related peptide. Euro. J. Pharmacol. 2009, 617, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Servent, D.; Delaforge, M.; Ducrocq, C.; Mansuy, D.; Lenfant, M. Nitric oxide formation during microsomal hepatic denitration of glyceryl trinitrate: Involvement of cytochrome P-450. Biochem. Biophys. Res. Commun. 1989, 163, 1210–1216. [Google Scholar] [CrossRef]
- McDonald, B.J.; Bennett, B.M. Cytochrome P-450-mediated biotransformation of organic nitrates. Can. J. Physiol. Pharmacol. 1990, 68, 1552–1557. [Google Scholar] [CrossRef]
- McDonald, B.J.; Bennett, B.M. Biotransformation of glyceryl trinitrate by rat aortic cytochrome P450. Biochem. Pharmacol. 1993, 45, 268–270. [Google Scholar] [CrossRef]
- Lopez, M.; Malacarne, P.F.; Gajos-Draus, A.; Ding, X.; Daiber, A.; Lundberg, J.O.; Offermanns, S.; Brandes, R.P.; Rezende, F. Vascular biotransformation of organic nitrates is independent of cytochrome P450 monooxygenases. Br. J. Pharmacol. 2021, 178, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, X.; Cui, H.; Chen, Y.-R.; Cardounel, A.J.; Zweier, J.L. Characterization of the mechanism of cytochrome P450 reductase-cytochrome P450-mediated nitric oxide and nitrosothiol generation from organic nitrates. J. Biol. Chem. 2006, 281, 12546–12554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, T.M.; Stevens, C.R.; Benjamin, N.; Eisenthal, R.; Harrison, R.; Blake, D.R. Xanthine oxidoreductase catalyses the reduction of nitrates and nitrite to nitroc oxide under hypoxic conditions. FEBS Lett. 1998, 427, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Doel, J.J.; Godber, B.L.J.; Eisenthal, R.; Harrison, R. Reduction of organic nitrates catalysed by xanthine oxidoreductase under anaerobic conditions. Biochim. Biophys. Acta 2001, 1527, 81–87. [Google Scholar] [CrossRef]
- Li, H.; Samouilov, A.; Liu, X.; Zweier, J.L. Characterization of the magnitude and kinetics of xanthine oxidase-catalysed nitrite reduction. J. Biol. Chem. 2001, 276, 24482–24489. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cui, H.; Kundu, T.K.; Alzawahra, W.; Zweier, J.L. Nitric oxide production from nitrite occurs primarily in tissues not in the blood. J. Biol. Chem. 2008, 283, 17855–17863. [Google Scholar] [CrossRef] [Green Version]
- Bernardino-Paula, R.; Carvalho-Galvão, A.; Cavalcanti, A.L.M.; Rocha, P.K.L.; Carvalho, L.R.R.A.; Brandão, M.C.R.; Athayde-Filho, P.F.; Diniz, T.F.; Lemos, V.S.; França-Silva, M.S.; et al. The new organic nitrate 2-nitrate-1,3-diocthanoxy propan (NDOP) induces nitric oxide production and vasorelaxation via activation of inward-rectifier potassium channels (KIR). Nitric Oxide 2020, 104–105, 61–69. [Google Scholar] [CrossRef]
- Axton, E.R.; Cristobal, E.; Choi, J.; Miranda, C.L.; Stevens, J.F. Metabolomics-driven elucidation of cellular nitrate tolerance reveals ascorbic acid prevents nitroglycerin-induced inactivation of xanthine oxidase. Front. Pharmacol. 2018, 9, 1085. [Google Scholar] [CrossRef] [PubMed]
- Golwala, N.H.; Hodenette, C.; Murthy, S.N.; Nossaman, B.D.; Kadowitz, P.J. Vascular responses to nitrite are mediated by xanthine oxidoreductase and mitochondrial aldehyde dehydrogenase in the rat. Can. J. Physiol. Pharmacol. 2009, 87, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Kitson, T.M. The disulfiram-ethanol reaction. J. Stud. Alc. 1977, 38, 96–113. [Google Scholar] [CrossRef]
- Koppaka, V.; Thompson, D.C.; Chen, Y.; Ellermann, M.; Nicolaou, K.C.; Juvonen, R.O.; Petersen, D.; Deitrich, R.A.; Hurley, T.D.; Vasiliou, V. Aldehyde dehydrogenase inhibitors: A comprehensive review of the pharmacology, mechanism of action, substrate specificity, and clinical application. Pharmacol. Rev. 2012, 64, 520–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, P.; Hink, U.; Oelze, M.; Seeling, A.; Isse, T.; Bruns, K.; Steinhoff, L.; Brandt, M.; Kleschyov, A.L.; Schulz, E.; et al. Number of nitrate groups determines reactivity and potency of organic nitrates: A proof of concept study in ALDH-2-/- mice. Brit. J. Pharmacol. 2007, 150, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Foster, M.W.; Zhang, J.; Mao, L.; Rockman, H.A.; Kawamoto, T.; Kitagawa, K.; Nakayama, K.I.; Hess, D.T.; Stamler, J.S. An essential role for mitochondria aldehyde dehydrogenase in nitroglycerin bioactivation. Proc. Nat. Acad. Sci. USA 2005, 102, 12159–12164. [Google Scholar] [CrossRef] [Green Version]
- Tsou, P.-S.; Page, N.A.; Lee, S.G.; Fung, S.M.; Keung, W.M.; Fung, H.-L. Differential metabolism of organic nitrates by aldehyde dehydrogenase 1a1 and 2: Substrate selectivity, enzyme inactivation, and active cysteine sites. Amer. Assoc. Pharmaceut. Sci. 2011, 13, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Page, N.A.; Fung, S.M.; Fung, H.-L. In vitro organic nitrate bioactivation to nitric oxide by recombinant aldehyde dehydrogenase 3A1. Nitric Oxide 2013, 35, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Beretta, M.; Wolkart, G.; Schernthaner, M.; Griesberger, M.; Neubauer, R.; Schmidt, K.; Sacherer, M.; Heinzel, F.R.; Kohlwein, S.D.; Mayer, B. Vascular bioactivation of nitroglycerin is catalyzed by cytosolic aldehyde dehydrogenase-2. Circ. Res. 2012, 110, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baretta, M.; Gruber, K.; Kollau, A.; Russwurm, M.; Koesling, D.; Goessler, W.; Keung, W.M.; Schmidt, K.; Mayer, B. Bioactivation of nitroglycerin by purified mitochondrial and cytosolic aldehyde dehydrogenases. J. Biol. Chem. 2008, 283, 17873–17880. [Google Scholar] [CrossRef] [Green Version]
- Kitson, T.M.; Hill, J.P.; Midwinter, G.G. Identification of a catalytically essential nucleophilic residue in sheep liver cytoplasmic aldehyde dehydrogenase. Biochem. J. 1991, 275, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzl, M.V.; Beretta, M.; Gorren, A.C.F.; Zeller, A.; Baral, P.K.; Gruber, K.; Russwurm, M.; Koesling, D.; Schmidt, K.; Mayer, B. Role of the general base Glu-268 in nitroglycerin bioactivation and superoxide formation by aldehyde dehydrogenase-2. J. Biol. Chem. 2009, 284, 19878–19886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Stamler, J.S. Bioactivation of nitroglycerin by mitochondrial aldehyde dehydrogenase. Trends Cardiovasc. Med. 2006, 16, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.L.; Lipsky, J.J.; Naylor, S. Role of disulfiram in the in vitro inhibition of rat liver mitochondrial aldehyde dehydrogenase. Biochem. Pharmacol. 2000, 60, 947–953. [Google Scholar] [CrossRef]
- Münzel, T.; Daiber, A.; Mülsch, A. Explaining the phenomenon of nitrate tolerance. Circ. Res. 2005, 97, 618–628. [Google Scholar] [CrossRef]
- Wenzl, V.M.; Beretta, M.; Griesberger, M.; Russwurm, M.; Koesling, D.; Schmidt, K.; Mayer, B.; Gorren, A.C.F. Site-directed mutagenesis of aldehyde dehydrogenase-2 suggests three distinct pathways of nitroglycerin biotransformation. Mol. Pharmacol. 2011, 80, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Lang, B.S.; Gorren, A.C.F.; Oberdorfer, G.; Wenzl, M.V.; Furdui, C.M.; Poole, L.B.; Mayer, B.; Gruber, K. Vascular bioactivation of nitroglycerin by aldehyde dehydrogenase-2. J. Biol. Chem. 2012, 287, 38124–38134. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.M.; Libedinsky, A.; Elorza, A.A. Role of copper on mitochondrial function and metabolism. Front. Mol. Biosci. 2021, 8, 711227. [Google Scholar] [CrossRef]
- Needleman, P.; Johnson, E.M., Jr. Mechanism of tolerance development to organic nitrates. J. Pharmacol. Exper. Therapeut. 1973, 184, 709–715. [Google Scholar]
- Opelt, M.; Wölkart, G. Sustained formation of nitroglycerin-derived nitric oxide by aldehyde dehydrogenase-2 in vascular smooth muscle without added reductants: Implications for the development of nitrate tolerance. Mol. Pharmacol. 2018, 93, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Hink, U.; Oelze, M.; Schuppan, S.; Schaeuble, K.; Schildknecht, S.; Ho, K.K.; Weiner, H.; Bachschmid, M.; Münzel, T.; et al. Role of reduced lipoic acid in the redox regulation of mitochondrial aldehyde dehydrogenase (ALDH-2) activity. J. Biol. Chem. 2007, 282, 792–799. [Google Scholar] [CrossRef] [Green Version]
- Bilska-Wilkosz, A.; Kotanska, M.; Górny, M.; Filipek, B.; Iciek, M. Is the mechanism of nitroglycerin tolerance associated with aldehyde dehydrogenase activity? A contribution to the ongoing discussion. Acta Biochim. Polon. 2019, 66, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Abrams, J. How to use nitrates. Cardiovasc. Drug Ther. 2002, 16, 511–514. [Google Scholar] [CrossRef]
- Daiber, A.; Wenzel, P.; Oelze, M.; Münzel, T. New insights into bioactivation of organic nitrates, nitrate tolerance and cross tolerance. Clin. Res. Cardiol. 2008, 97, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Sayed, N.; Kim, D.D.; Fioramonti, X.; Iwahashi, T.; Durán, W.N.; Beuve, A. Nitroglycerin-induced S-nitrosylation and desensitization of soluble guanylyl cyclase contribute to nitrate tolerance. Circ. Res. 2008, 103, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Stamler, J.S. Nitroglycerin-mediated S-nitrosylation of proteins: A field comes full cycle. Circ. Res. 2008, 103, 557–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laursen, J.B.; MüIsch, A.; Boesgaard, S.; Mordvintcev, P.; Trautner, S.; Gruhn, N.; Nielsen-Kudsk, J.E.; Busse, R.; Aldershvile, J. In vivo nitrate tolerance is not associated with reduced bioconversion of nitroglycerin to nitric oxide. Circulation 1996, 94, 2241–2247. [Google Scholar] [CrossRef]
- Münzel, T.; Sayegh, H.; Freeman, B.A.; Tarpey, M.M.; Harrison, D.G. Evidence for enhanced vascular superoxide anion production in nitrate tolerance. J. Clin. Investig. 1995, 95, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Mollace, V.; Musoli, C.; Dagostino, C.; Giancotti, L.A.; Gliozzi, M.; Sacco, I.; Visalli, V.; Gratteri, S.; Palma, E.; Malara, N.; et al. The effect of peroxynitrite decomposition catalyst MnTBAP on aldehyde dehydrogenase-2 nitration by organic nitrates: Role in nitrate tolerance. Pharmacol. Res. 2014, 89, 29–35. [Google Scholar] [CrossRef]
- Garcia-Bou, R.; Rocha, M.; Apostolova, N.; Herance, R.; Hernandez-Mijares, A.; Victor, V.M. Evidence for a relationship between mitochondrial complex 1 activity and mitochondrial aldehyde dehydrogenase during nitroglycerin tolerance: Effects of mitochondrial antioxidants. Biochim. Biophys. Acta 2012, 1817, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Klemenska, E.; Beresewicz, A. Bioactivation of organic nitrates and the mechanism of nitrate tolerance. Cardiol. J. 2009, 16, 11–19. [Google Scholar]
- Watanabe, H.; Kakihana, M.; Ohtsuka, S.; Sugishita, Y. Randomized, double-blind, placebo-controlled study of supplemental vitamin E on attenuation of the development of nitrate tolerance. Circulation 1997, 96, 2545–2550. [Google Scholar] [CrossRef] [PubMed]
- Fink, B.; Dikalov, S.; Bassenge, E. A new approach for extracellular spin trapping of nitroglycerin-induced superoxide radicals both in vitro and in vivo. Free Rad. Biol. Med. 2000, 28, 121–128. [Google Scholar] [CrossRef]
- Coskun, B.; Soylemez, S.; Parlar, A.I.; Ulus, A.T.; Katircioglu, S.F.; Akar, F. Effect of resveratrol on nitrate tolerance in isolated human internal mammary artery. J. Cardiovasc. Pharmacol. 2006, 47, 437–445. [Google Scholar] [PubMed]
- D’Souza, Y.; Kawamoto, T.; Bennett, B.M. Role of lipid peroxidation product, 4-hydroxynonenal, in the development of nitrate tolerance. Chem. Res. Toxicol. 2014, 27, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Larson, H.N.; Zhou, J.; Chen, Z.; Stamler, J.S.; Weiner, H.; Hurley, T.D. Structural and functional consequences of coenzyme binding to the inactive Asian variant of mitochondrial aldehyde dehydrogenase. J. Biol. Chem. 2007, 282, 12940–12950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, D.; Jin, W.; Shao, C.; Yan, P.; Xu, C.; Sheng, H.; Liu, Y.; Yu, J.; Xie, Y.; et al. Mitochondrial aldehyde dehydrogenase-2 (ALDH2) Glu504Lys polymorphism contributes to the variation in efficacy of sublingual nitroglycerin. J. Clin. Investig. 2006, 116, 506–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedde, H.W.; Agarwal, D.P.; Harada, S.; Meier-Tackmann, D.; Ruofu, D.; Bienzle, U.; Kroeger, A.; Hussein, L. Population genetic studies on aldehyde dehydrogenase isozyme deficiency and alcohol sensitivity. Am. J. Hum. Genet. 1983, 35, 769–772. [Google Scholar] [PubMed]
- Mackenzie, I.S.; Maki-Petaja, K.M.; McEniery, C.M.; Bao, Y.P.; Wallace, S.M.; Cheriyan, J.; Monteith, S.; Brown, M.J.; Wilkinson, I.B. Aldehyde dehydrogenase 2 plays a role in the bioactivation of nitroglycerin in humans. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1891–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Fritze, G.; Fang, B.; Harada, S.; Paik, Y.K.; Eckey, R.; Agarwal, D.P.; Goedde, H.W. Inheritance of mitochondrial aldehyde dehydrogenase: Genotyping in Chinese, Japanese and South Korean families reveals dominance of the mutant allele. Hum. Gen. 1989, 83, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T.; Ushijima, K.; Taga, N.; Takeuchi, M.; Kawada, M.; Aizawa, K.; Imai, Y.; Fujimura, A. Influence of the aldehyde dehydrogenase 2 polymorphism on the vasodilatory effect of nitroglycerin in infants with congenital heart disease and pulmonary arterial hypertension. Euro. J. Clin. Pharmacol. 2019, 75, 1361–1367. [Google Scholar] [CrossRef]
- Miura, T.; Nishinaka, T.; Terada, T.; Yonezawa, K. Vasodilatory effect of nitroglycerin in Japanese subjects with different aldehyde dehydrogenase 2 (ALDH2) genotypes. Chem.-Biol. Interact. 2017, 276, 40–45. [Google Scholar] [CrossRef] [PubMed]
- He, J.D.; Lytvyn, Y.; Zhou, K.B.; Parker, J.D. Role of mitochondrial aldehyde dehydrogenase in nitroglycerin-mediated vasodilation: Observations concerning the dose-response relationship. J. Cardiovasc. Pharmacol. 2019, 73, 359–364. [Google Scholar] [CrossRef]
- Xia, J.-Q.; Song, J.; Zhang, Y.; An, N.-N.; Ding, L.; Zhang, Z. Effect of aldehyde dehydrogenase 2 gene polymorphism on hemodynamics after nitroglycerin intervention in Northern Chinese Han population. Chin. Med. J. 2015, 128, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Farrés, J.; Wang, X.; Takahashi, K.; Cunningham, S.J.; Wang, T.T.; Weiner, H. Effects of changing glutamate 487 to lysine in rat and human liver mitochondrial aldehyde dehydrogenase. J. Biol. Chem. 1994, 269, 13854–13860. [Google Scholar] [CrossRef]
- Arif, S.; Borgognone, A.; Lai-Sze Lin, E.; O’Sullivan, A.G.; Sharma, V.; Drury, N.E.; Menon, A.; Nightingale, P.; Mascaro, J.; Bonser, R.S.; et al. Role of aldehyde dehydrogenase in hypoxic vasodilator effects of nitrite in rats and humans. Br. J. Pharmacol. 2015, 172, 3341–3352. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, Y.; Ji, Y.; Bennett, B.M. Effect of overexpression of human aldehyde dehydrogenase 2 in LL-PK1 cells on glyceryl trinitrate biotransformation and cGMP accumulation. Br. J. Pharmacol. 2013, 168, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Seabra, A.B.; Ouellet, M.; Antonic, M.; Chrétien, M.N.; English, A.M. Catalysis of nitrite generation from nitroglycerin by glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Nitric Oxide 2013, 35, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Y.-G.; Xu, F.; Xue, L.; Jiang, C.-X.; Zhang, Y. The relationship between aldehyde dehydrogenase-2 gene polymorphisms and efficacy of nitroglycerin. Zhonghua Nei Ke Za Zhi 2007, 46, 629–632. [Google Scholar] [PubMed]
- Ferreira, J.C.B.; Mochly-Rosen, D. Nitroglycerin use in myocardial infarction patients: Risks and benefits. Circ. J. 2012, 76, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellon, D.M.; He, Z.; Khambata, R.; Ahluwalia, A.; Davidson, S.M. The GTN patch: A simple and effective new approach to cardioprotection? Basic Res. Cardiol. 2018, 113, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pearson, R.; Butler, A. Glyceryl Trinitrate: History, Mystery, and Alcohol Intolerance. Molecules 2021, 26, 6581. https://doi.org/10.3390/molecules26216581
Pearson R, Butler A. Glyceryl Trinitrate: History, Mystery, and Alcohol Intolerance. Molecules. 2021; 26(21):6581. https://doi.org/10.3390/molecules26216581
Chicago/Turabian StylePearson, Russell, and Anthony Butler. 2021. "Glyceryl Trinitrate: History, Mystery, and Alcohol Intolerance" Molecules 26, no. 21: 6581. https://doi.org/10.3390/molecules26216581
APA StylePearson, R., & Butler, A. (2021). Glyceryl Trinitrate: History, Mystery, and Alcohol Intolerance. Molecules, 26(21), 6581. https://doi.org/10.3390/molecules26216581