
Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods

 ,
,  , ,
, ,  ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
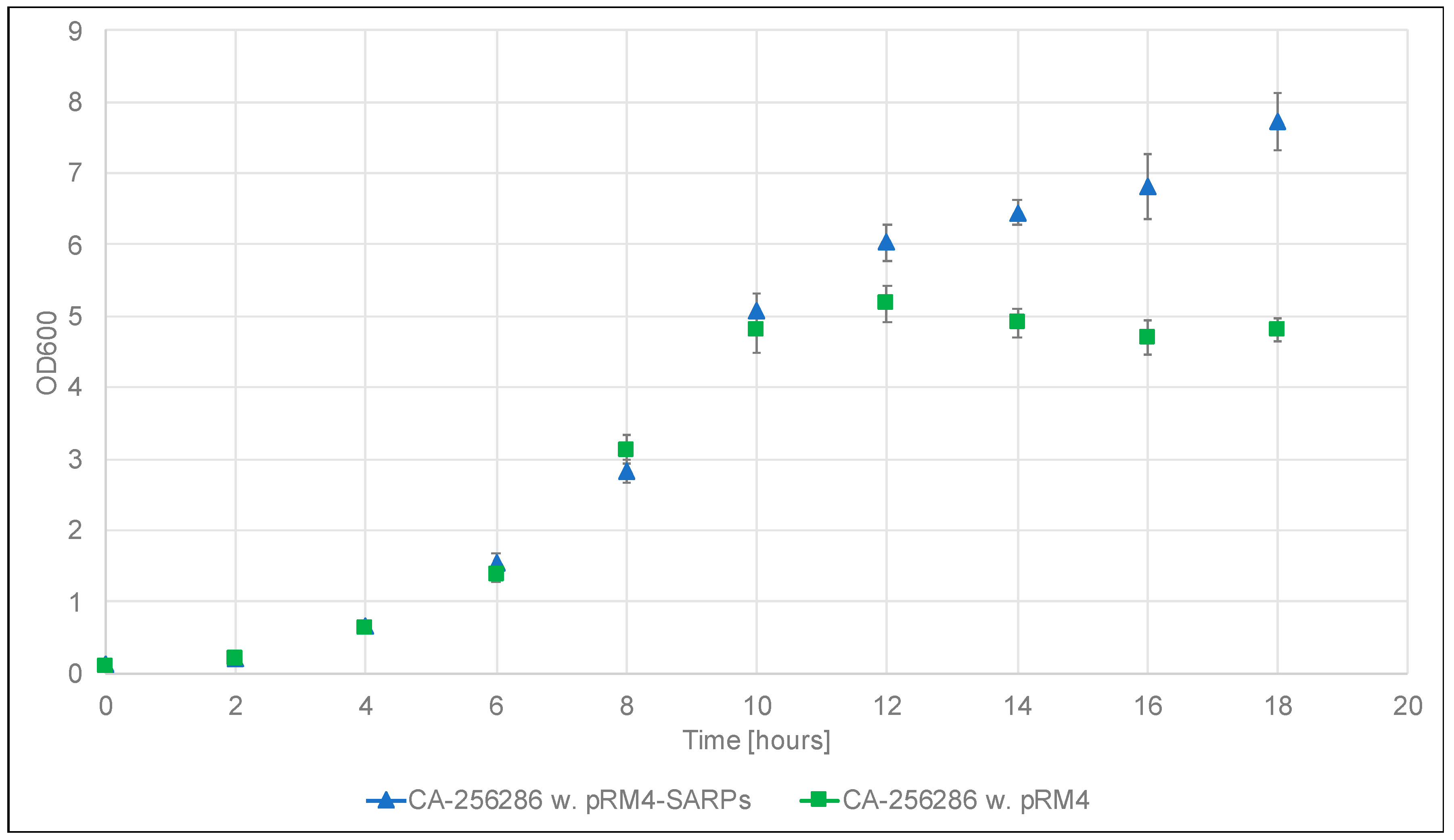
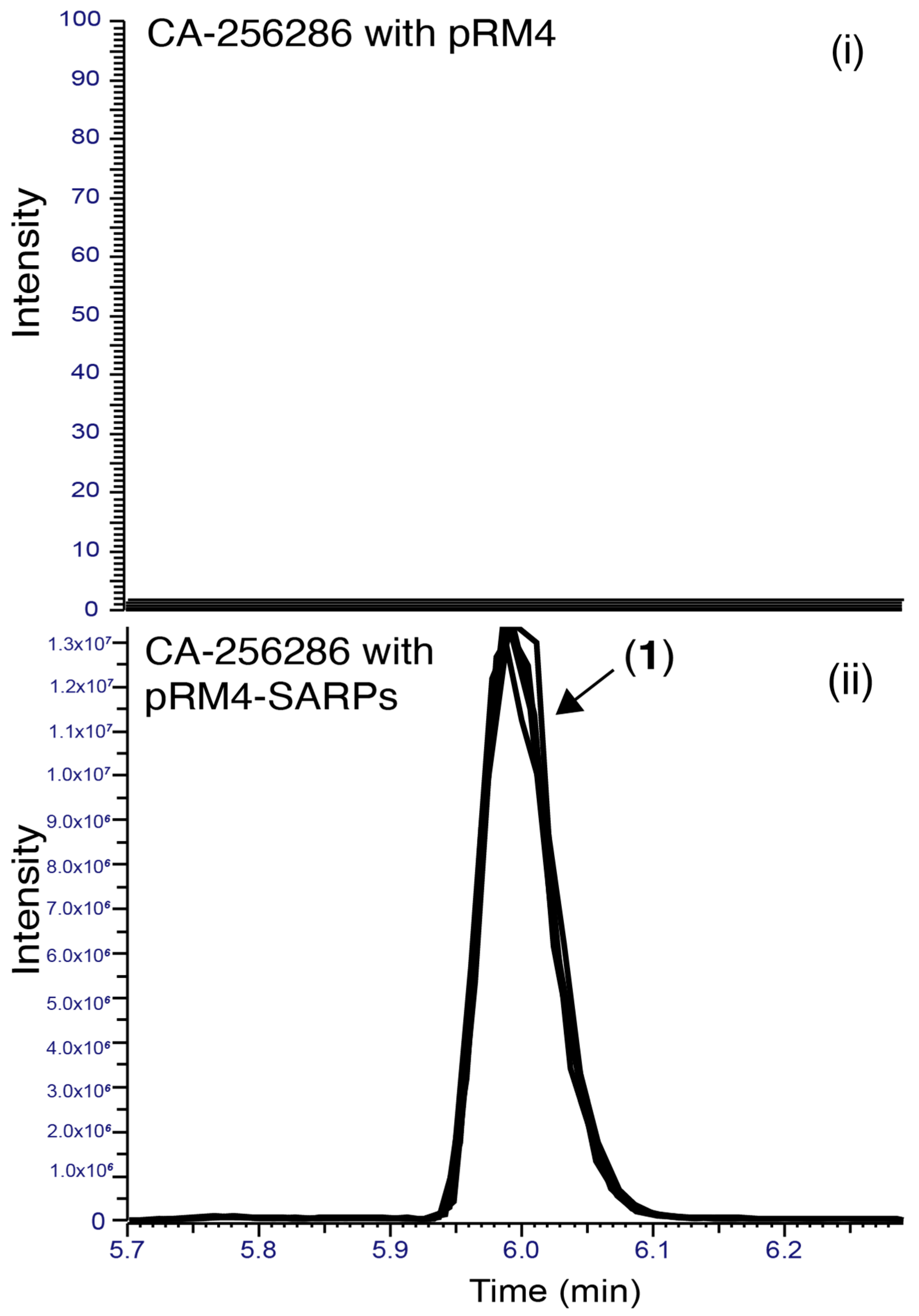
2.1. Transfer of Plasmids Encoding Transcriptional Regulators, High Throughput Cultivation and Metabolomics
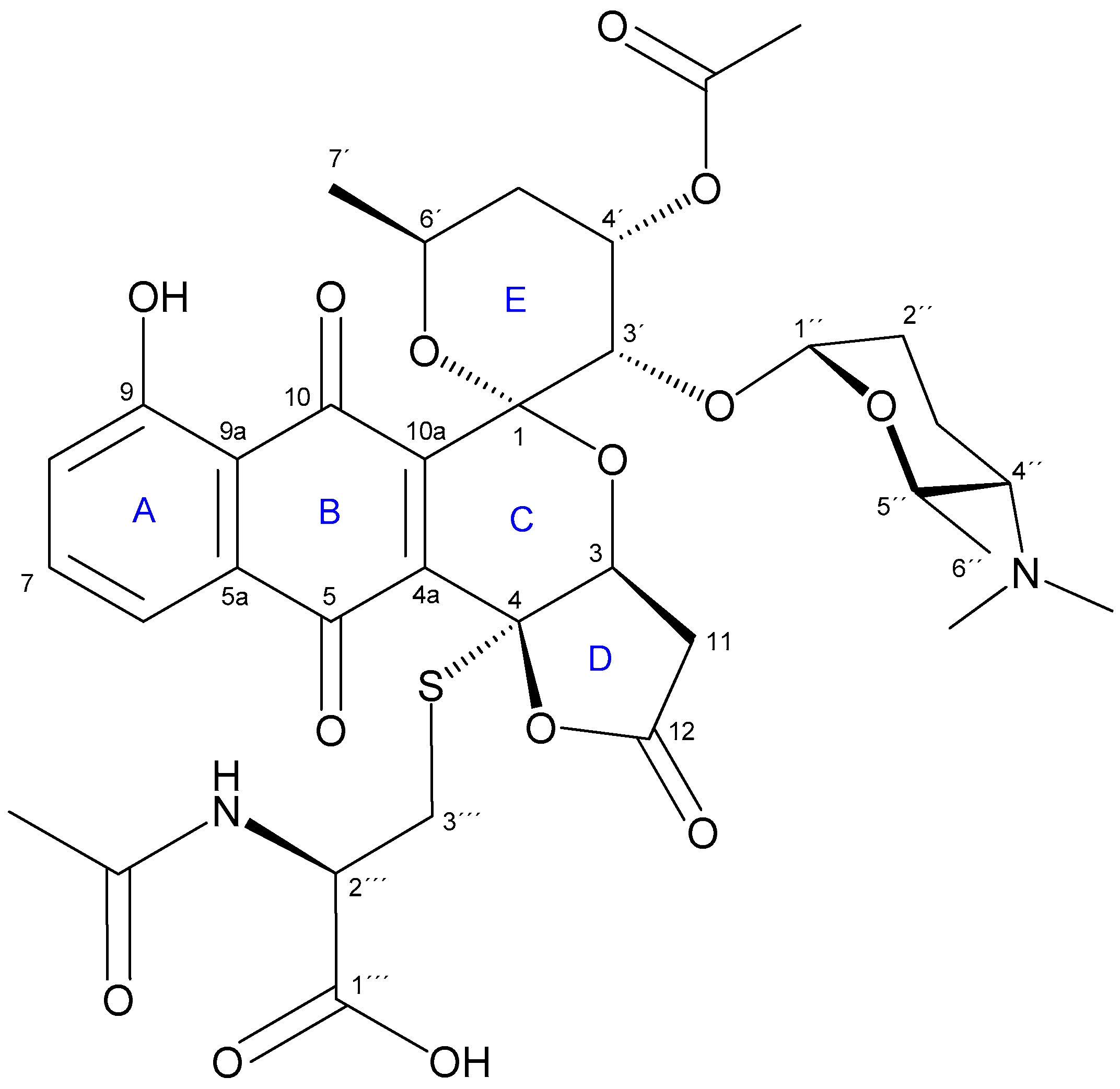
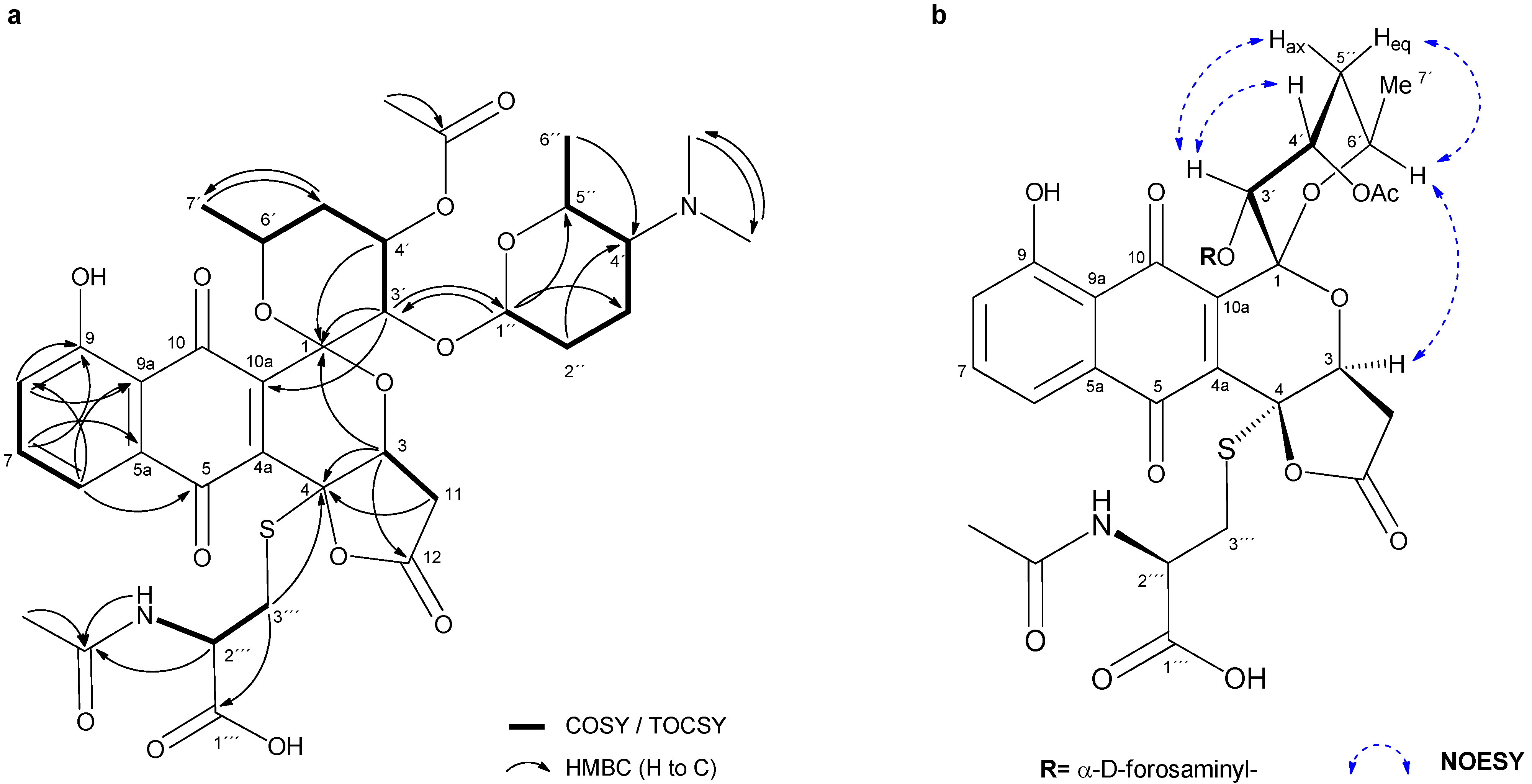
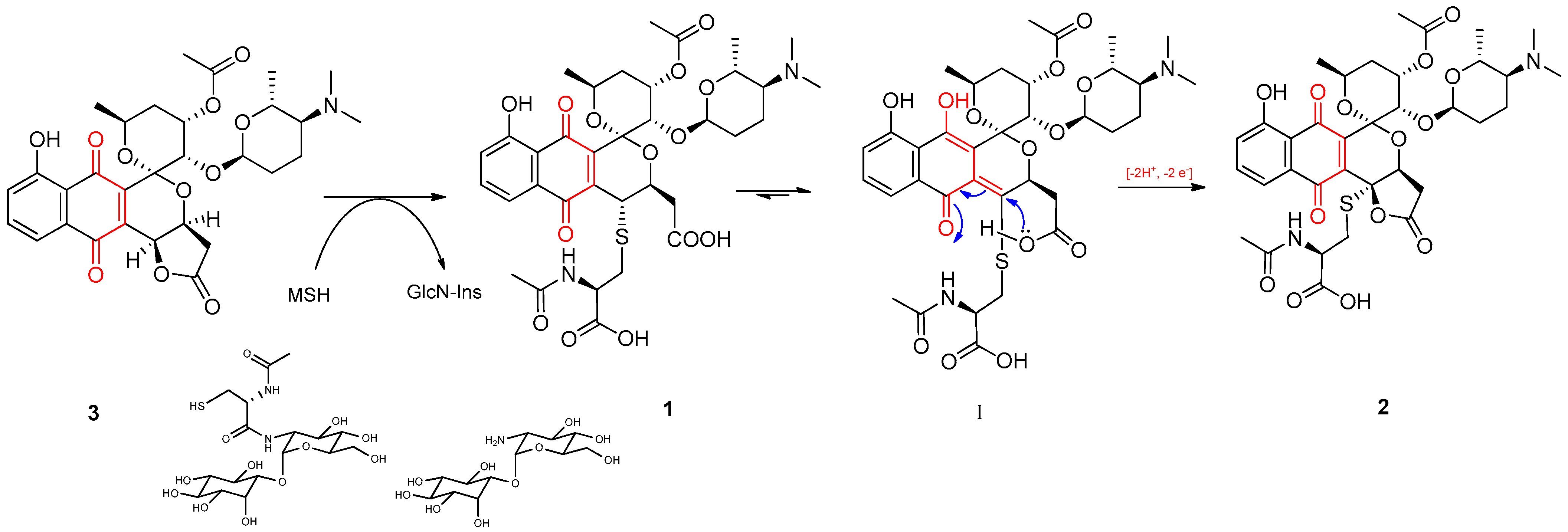
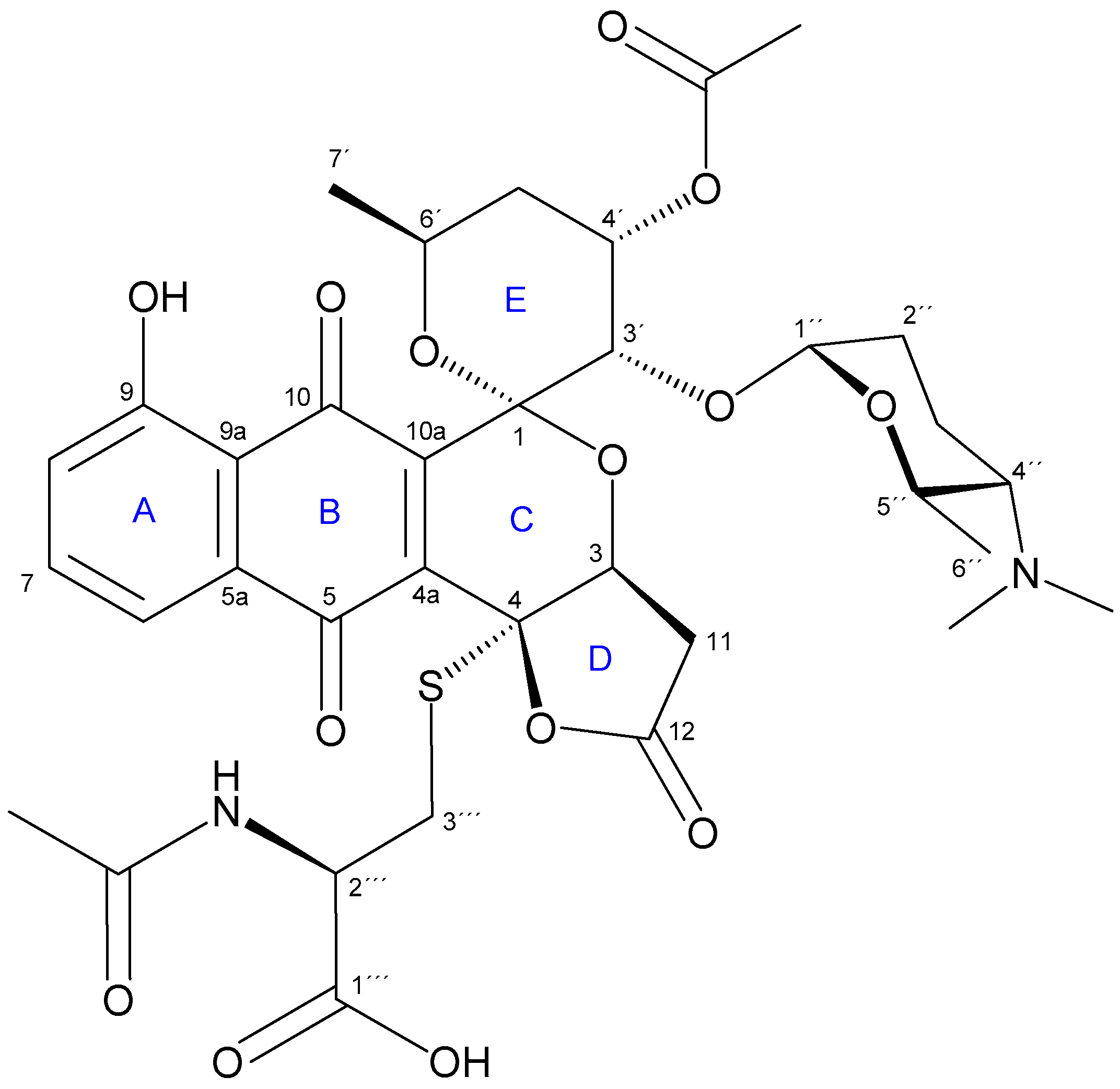
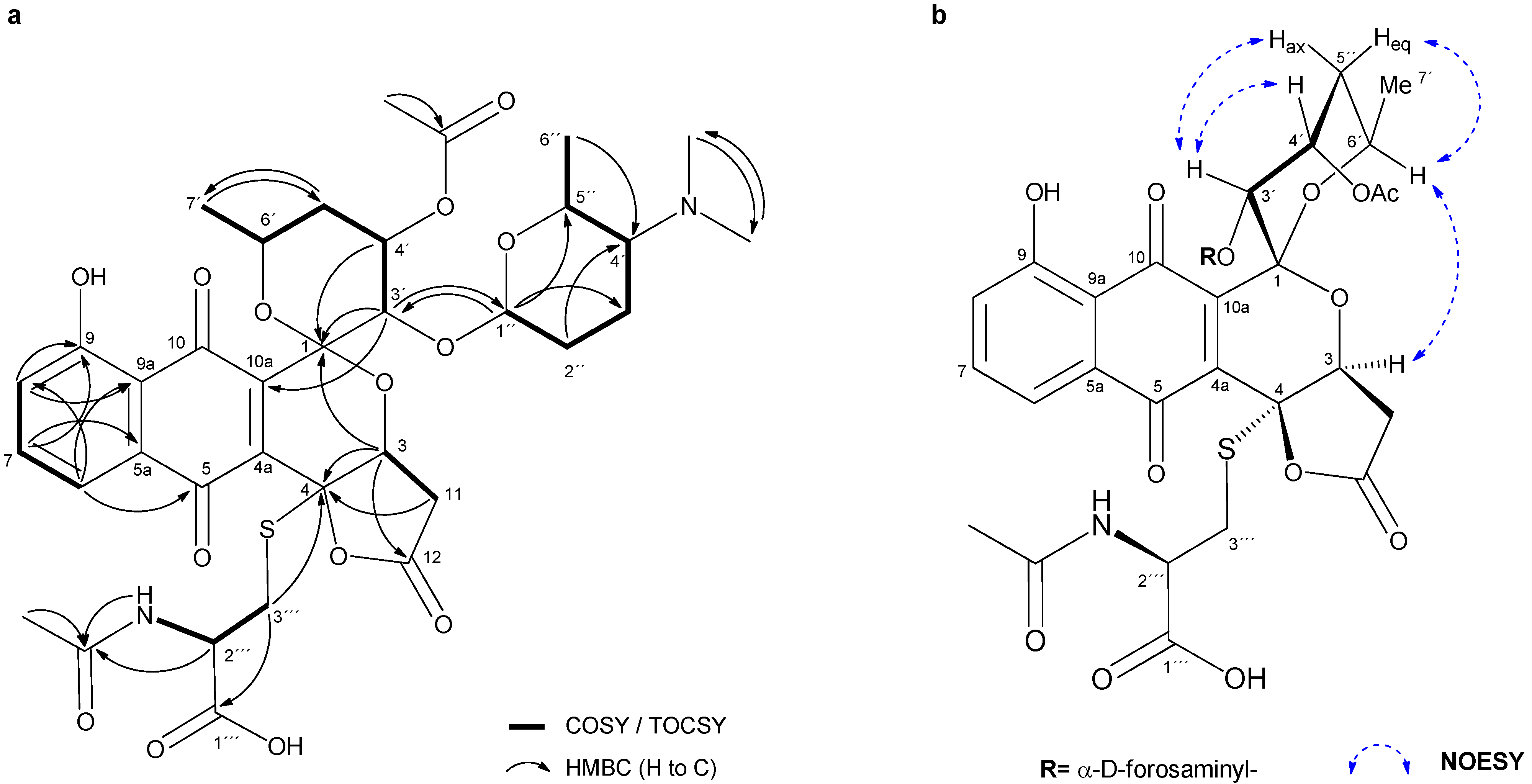
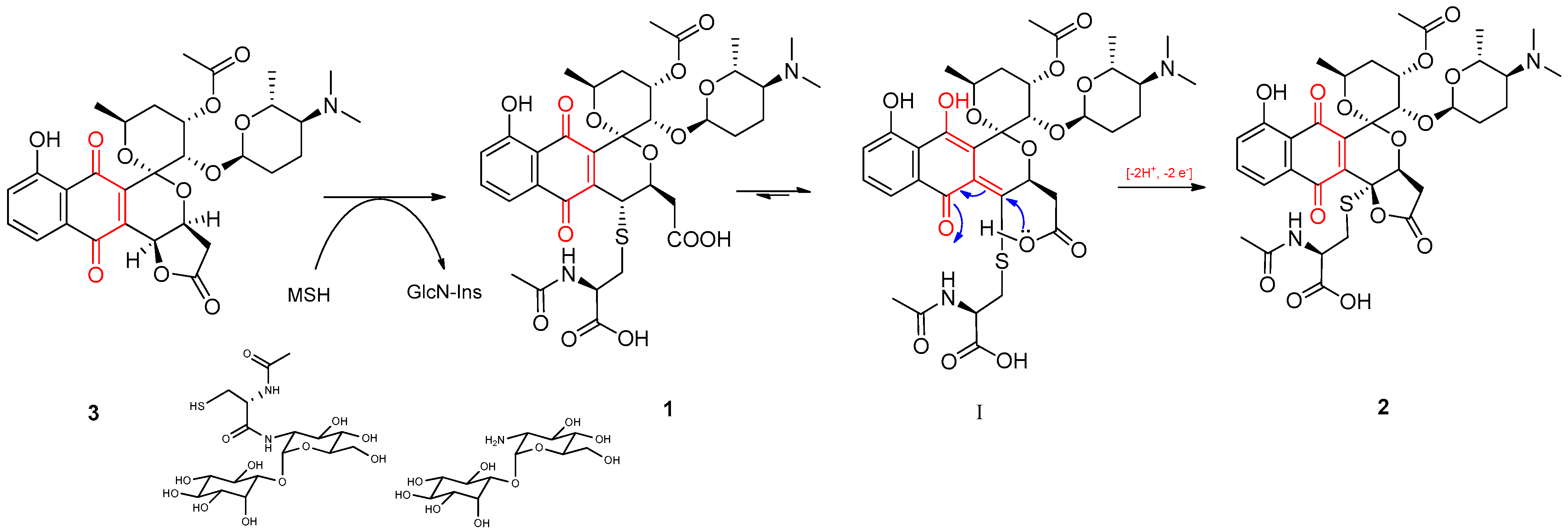
2.2. Isolation and Structure Elucidation of Griseusin-Derived N-Acetyl Cysteine Adducts 1 and 2
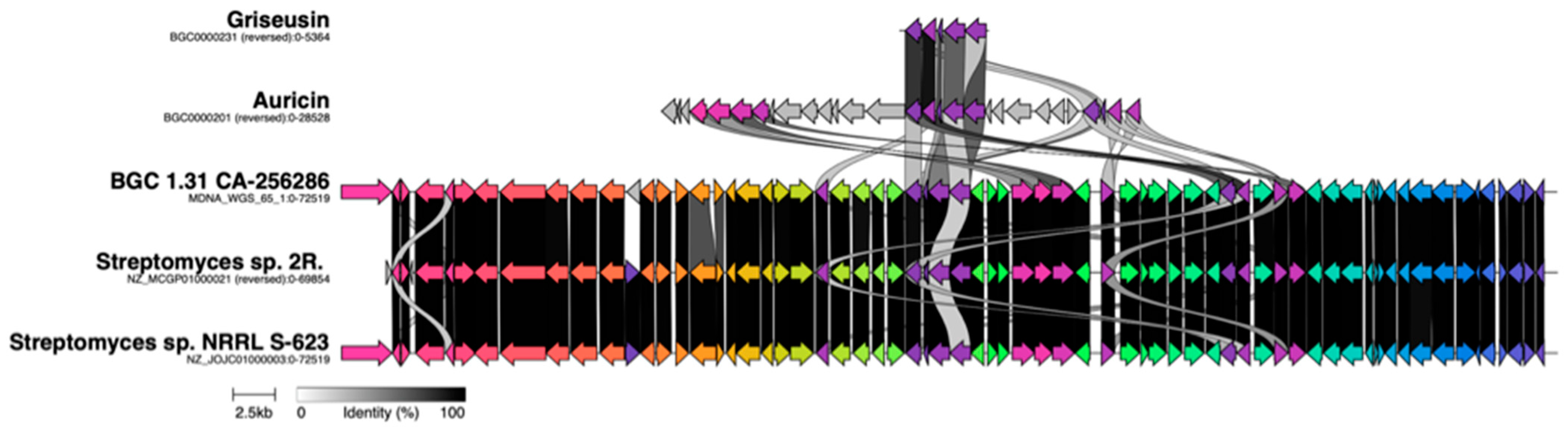
2.3. Sequencing of CA-256286, Genome Mining and Identification of Candidate BGCs
2.4. Cultivation and Sampling for Omics Studies
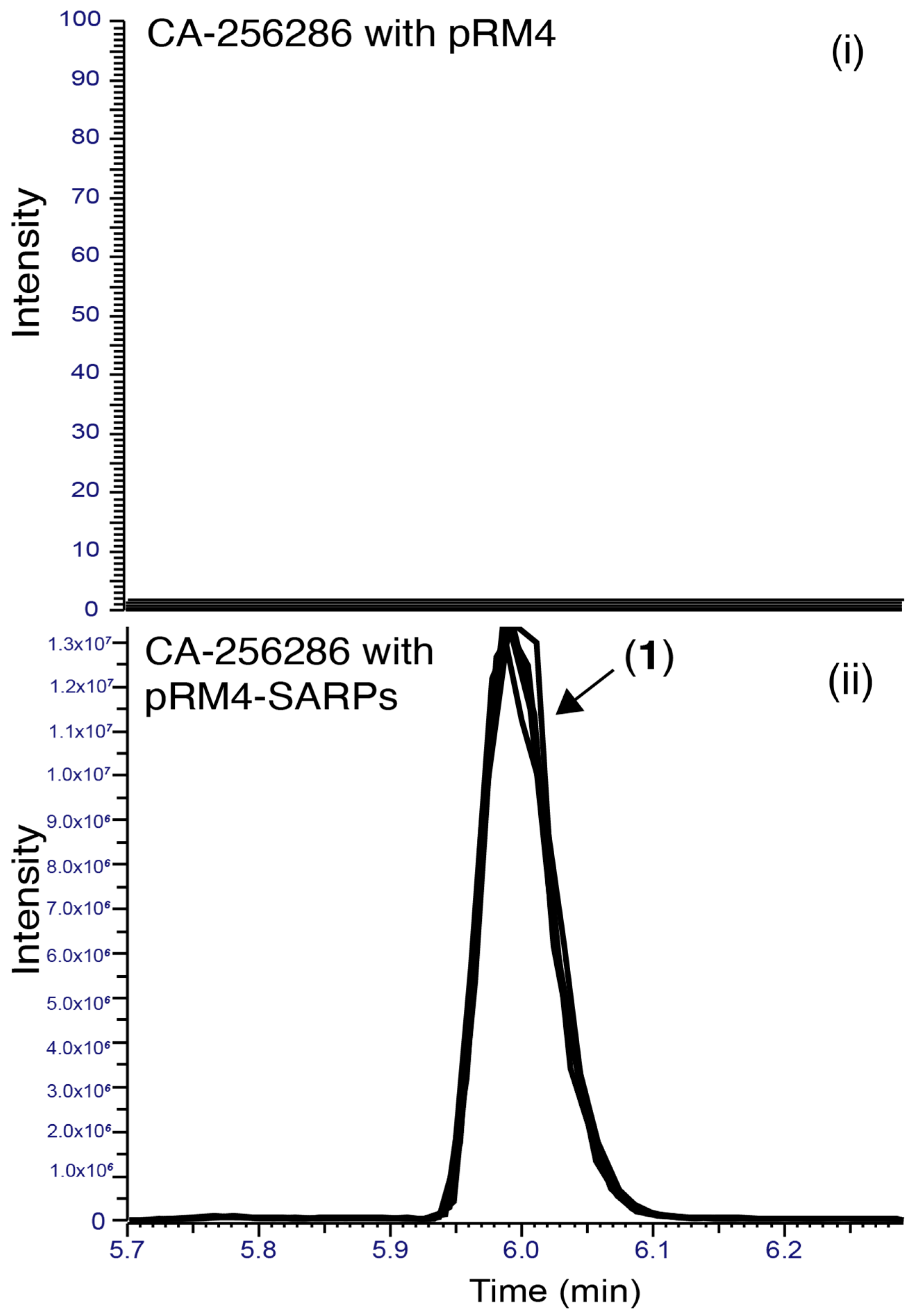
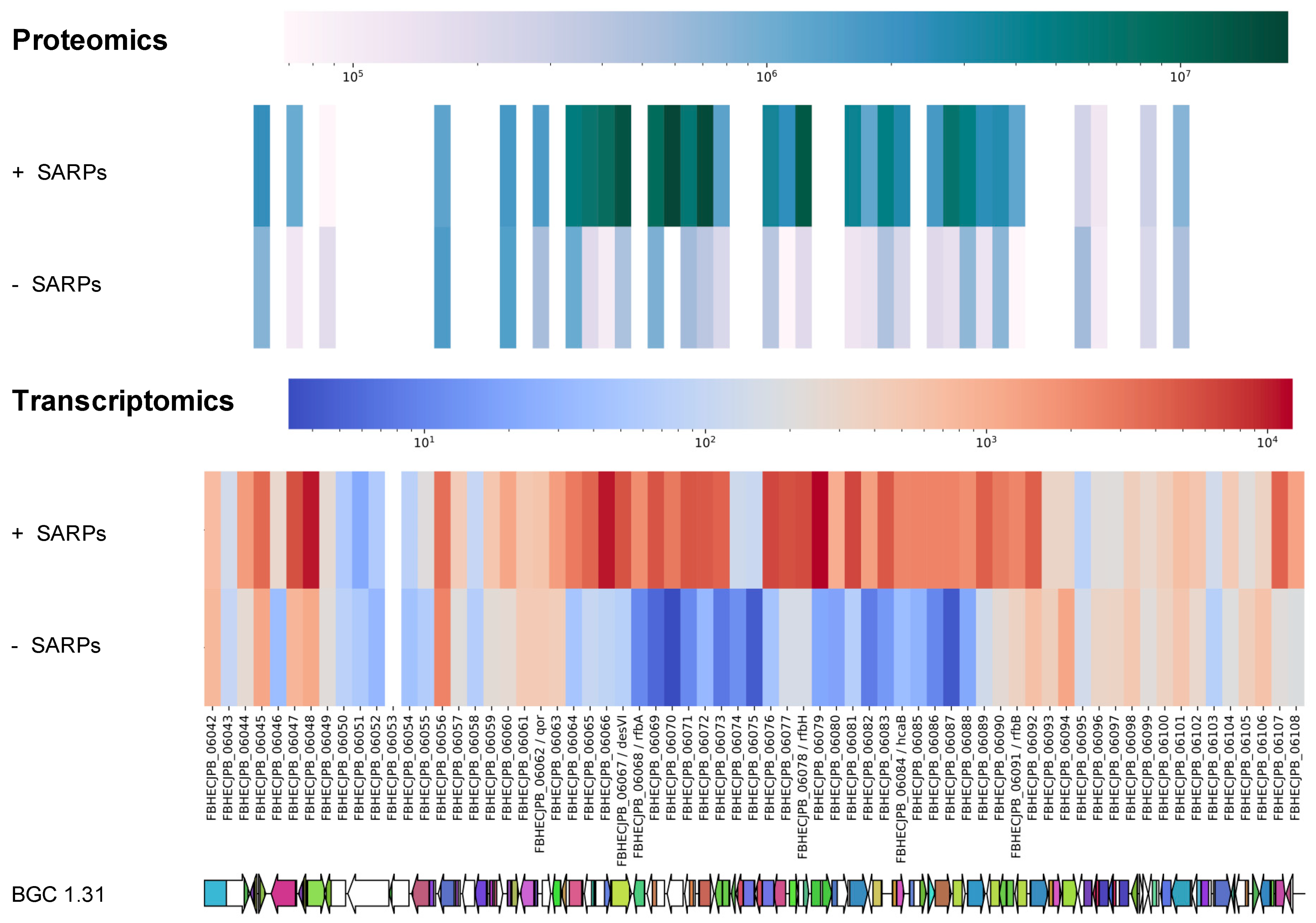
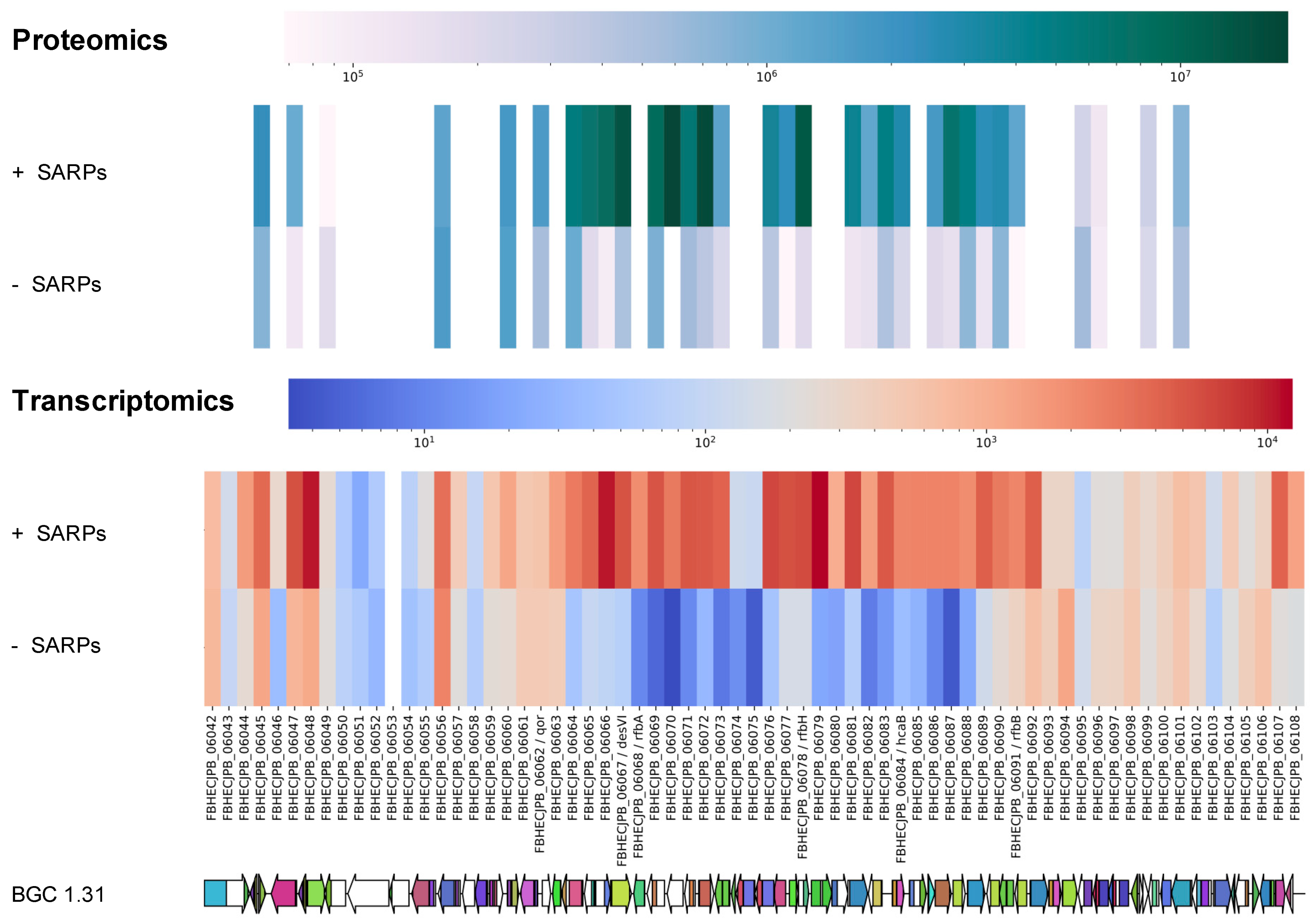
2.5. Proteomics and Transcriptomics Analyses Confirms Identity of the Griseusin BGC
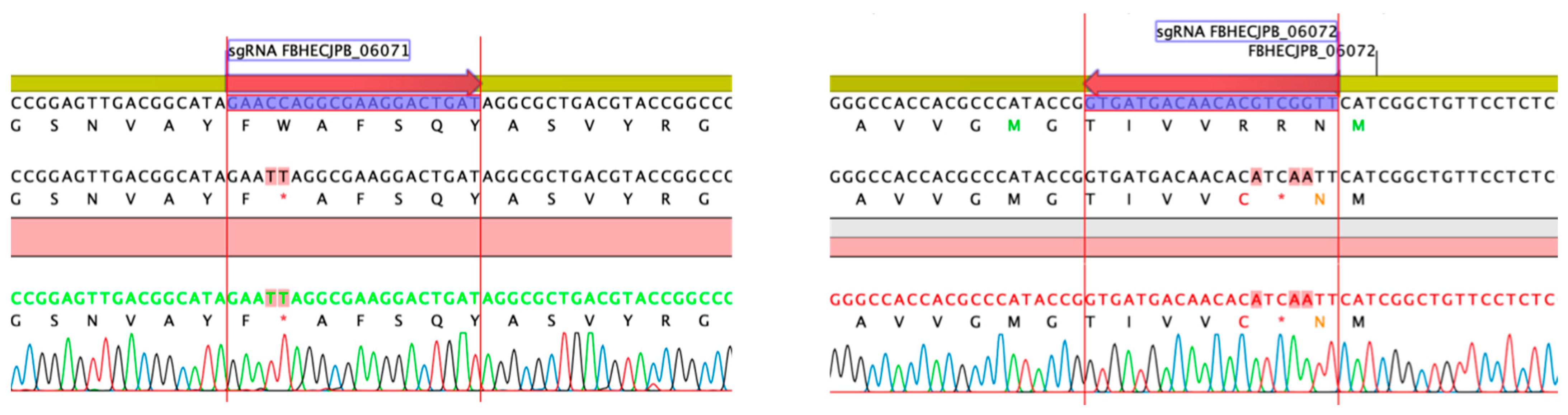
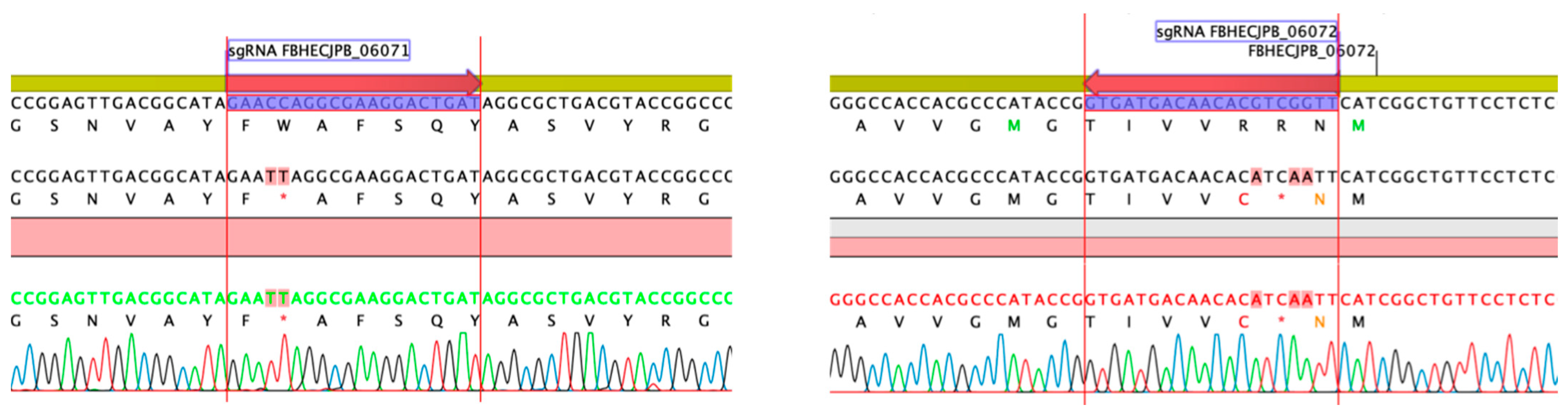
2.6. Gene Inactivation of the Griseusin PKS KS/CLF by CRISPR-cBEST Base Editing
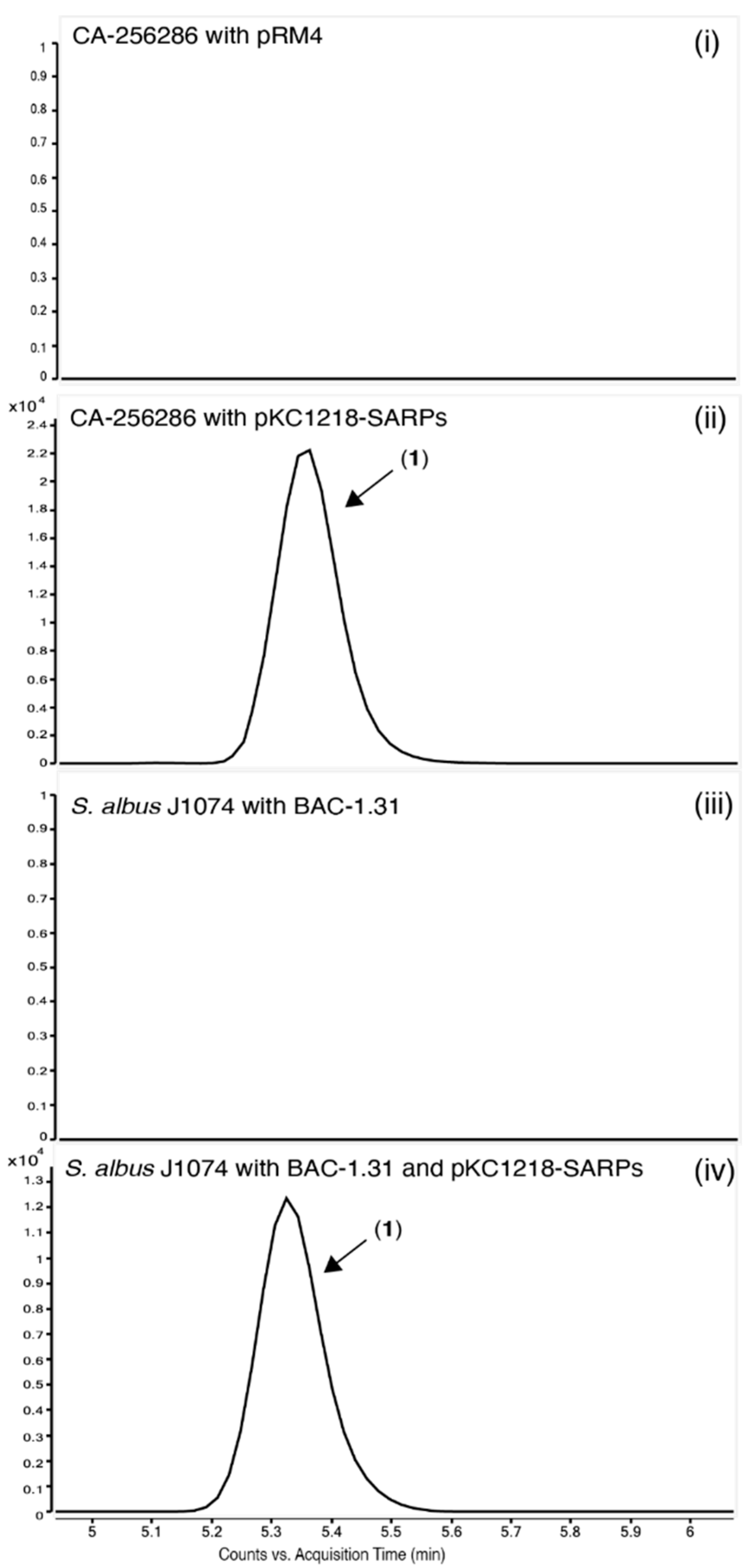
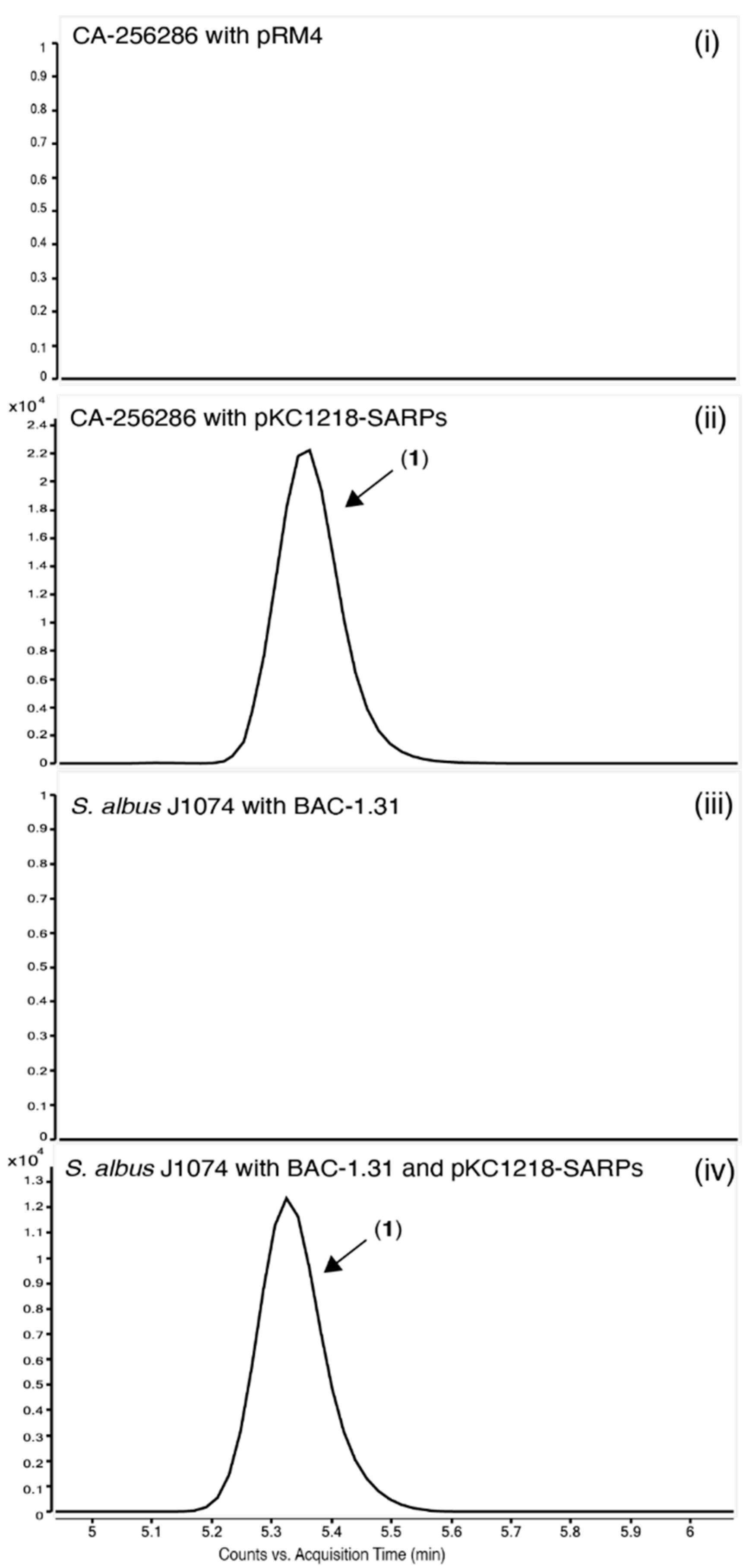
2.7. Heterologous Expression of the Putative Griseusin BGC in the Heterologous Host S. albus J1074
2.8. Identification of the Responsible Activator
2.9. The Griseusin Biosynthesis Pathway
3. Conclusions
4. Materials and Methods
4.1. Strains and Cultivations
4.2. General Molecular Biology Techniques
4.3. Plasmids
4.4. BAC Library and Heterologous Expression
4.5. Comparative Metabolomics with LC-MS
4.6. General Experimental Procedures
4.7. Isolation and Characterization Data of N-AcCys Adduct 2
4.8. RNA seq. and Transcriptomics Analysis
4.9. Proteomics
4.10. CRISPR-cBEST Inactivations and Verification
4.11. Antimicrobial Sensitivity Testing
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Donadio, S.; Staver, M.J.; McAlpine, J.B.; Swanson, S.J.; Katz, L. Modular organization of genes required for complex polyketide biosynthesis. Science 1991, 252, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Marahiel, M.A. A structural model for multimodular NRPS assembly lines. Nat. Prod. Rep. 2016, 33, 136–140. [Google Scholar] [CrossRef]
- Walsh, C.T. Insights into the chemical logic and enzymatic machinery of NRPS assembly lines. Nat. Prod. Rep. 2016, 33, 127–135. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Hoskisson, P.A.; Seipke, R.F. Cryptic or Silent? The known unknowns, unknown knowns, and unknown unknowns of secondary metabolism. mBio 2020, 11, e02642-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent advances in silent gene cluster activation in Streptomyces. Front. Bioeng. Biotechnol. 2021, 9, 632230. [Google Scholar] [CrossRef] [PubMed]
- Mingyar, E.; Mühling, L.; Kulik, A.; Winkler, A.; Wibberg, D.; Kalinowski, J.; Blin, K.; Weber, T.; Wohlleben, W.; Stegmann, E. A regulator based “semi-targeted” approach to activate silent biosynthetic gene clusters. Int. J. Mol. Sci. 2021, 22, 7567. [Google Scholar] [CrossRef] [PubMed]
- Bibb, M.J. Regulation of secondary metabolism in streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef]
- Krause, J.; Handayani, I.; Blin, K.; Kulik, A.; Mast, Y. Disclosing the potential of the sARP-type regulator PapR2 for the activation of antibiotic gene clusters in Streptomycetes. Front. Microbiol. 2020, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Menges, R.; Muth, G.; Wohlleben, W.; Stegmann, E. The ABC transporter Tba of Amycolatopsis balhimycina is required for efficient export of the glycopeptide antibiotic balhimycin. Appl. Microbiol. Biotechnol. 2007, 77, 125–134. [Google Scholar] [CrossRef]
- Bibb, M.J.; Janssen, G.R.; Ward, J.M. Cloning and analysis of the promoter region of the erythromycin resistance gene (ermE) of Streptomyces erythraeus. Gene 1985, 38, 215–226. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000; ISBN 9780708406236. [Google Scholar]
- Myronovskyi, M.; Welle, E.; Fedorenko, V.; Luzhetskyy, A. Beta-glucuronidase as a sensitive and versatile reporter in actinomycetes. Appl. Environ. Microbiol. 2011, 77, 5370–5383. [Google Scholar] [CrossRef] [Green Version]
- Martínez, G.; González-Menéndez, V.; Martín, J.; Reyes, F.; Genilloud, O.; Tormo, J.R. MASS Studio: A novel software utility to simplify LC-MS analyses of large sets of samples for metabolomics. Lect. Notes Comput. Sci. 2017, 10208, 230–244. [Google Scholar]
- Naysmith, B.J.; Hume, P.A.; Sperry, J.; Brimble, M.A. Pyranonaphthoquinones—Isolation, biology and synthesis: An update. Nat. Prod. Rep. 2017, 34, 25–61. [Google Scholar] [CrossRef]
- Metsä-Ketelä, M.; Oja, T.; Taguchi, T.; Okamoto, S.; Ichinose, K. Biosynthesis of pyranonaphthoquinone polyketides reveals diverse strategies for enzymatic carbon-carbon bond formation. Curr. Opin. Chem. Biol. 2013, 17, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Nakagawa, A.; Neszmélyi, A.; Gero, S.D.; Sepulchre, A.M.; Piriou, F.; Lukacs, G. Carbon-13 nuclear magnetic resonance spectral analysis of 16-membered macrolide antibiotics. J. Am. Chem. Soc. 1975, 97, 4001–4009. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.-G.; Zhao, J.-Y.; Li, M.-G.; Huang, R.; Li, Q.-M.; Cui, X.-L.; Zhu, H.-J.; Wen, M.-L. Griseusins F and G, Spiro-naphthoquinones from a tin mine tailings-derived alkalophilic Nocardiopsis species. J. Nat. Prod. 2012, 75, 1994–1998. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Nishida, C.; Takahashi, Y.; Naganawa, H.; Hamada, M.; Takeuchi, T. 3’-O-alpha-D-forosaminyl-(+)-griseusin A from Streptomyces griseus. J. Antibiot. 1994, 47, 952–954. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, T.; Yabe, M.; Odaki, H.; Shinozaki, M.; Metsä-Ketelä, M.; Arai, T.; Okamoto, S.; Ichinose, K. Biosynthetic conclusions from the functional dissection of oxygenases for biosynthesis of actinorhodin and related Streptomyces antibiotics. Chem. Biol. 2013, 20, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, T.; Maruyama, T.; Sawa, R.; Igarashi, M.; Okamoto, S.; Ichinose, K. Structure and biosynthetic implication of 5R-(N-acetyl-L-cysteinyl)-14S-hydroxy-dihydrokalafungin from a mutant of the actVA-ORF4 gene for actinorhodin biosynthesis in Streptomyces coelicolor A3(2). J. Antibiot. 2015, 68, 481–483. [Google Scholar] [CrossRef]
- He, J.; Roemer, E.; Lange, C.; Huang, X.; Maier, A.; Kelter, G.; Jiang, Y.; Xu, L.-H.; Menzel, K.-D.; Grabley, S.; et al. Structure, derivatization, and antitumor activity of new griseusins from Nocardiopsis sp. J. Med. Chem. 2007, 50, 5168–5175. [Google Scholar] [CrossRef]
- Tatsuta, K.; Akimoto, K.; Annaka, M.; Ohno, Y.; Kinoshita, M. Enantiodivergent total syntheses of nanaomycins and their enantiomers, kalafungins. Bull. Chem. Soc. Jpn. 1985, 58, 1699–1706. [Google Scholar] [CrossRef]
- Fernandes, R.A.; Brueckner, R. Efficient synthesis of (+)-kalafungin and (-)-nanaomycin D by asymmetric dihydroxylation, Oxa-Pictet—Spengler cyclization, and H2SO4-mediated isomerization. ChemInform 2005, 36, 1281–1285. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, Q.; Ponomareva, L.V.; Cao, Y.; Liu, Y.; Cui, Z.; Van Lanen, S.G.; Voss, S.R.; She, Q.-B.; Thorson, J.S. Total synthesis of griseusins and elucidation of the griseusin mechanism of action. Chem. Sci. 2019, 10, 7641–7648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, N.; Kobayashi, M.; Terui, Y.; Tori, K. The structures of griseusins A and B, new isochromanquinone antibiotics. Tetrahedron 1976, 32, 2207–2210. [Google Scholar] [CrossRef]
- Kometani, T.; Takeuchi, Y.; Yoshii, E. Pyranonaphthoquinone antibiotics. 4. Total synthesis of (+)-griseusin A, an enantiomer of the naturally occurring griseusin A. J. Org. Chem. 1983, 48, 2311–2314. [Google Scholar] [CrossRef]
- Franco, R.; Schoneveld, O.J.; Pappa, A.; Panayiotidis, M.I. The central role of glutathione in the pathophysiology of human diseases. Arch. Physiol. Biochem. 2007, 113, 234–258. [Google Scholar] [CrossRef]
- Newton, G.; Fahey, R. Mycothiol biochemistry. Arch. Microbiol. 2002, 178, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Jothivasan, V.K.; Hamilton, C.J. Mycothiol: Synthesis, biosynthesis and biological functions of the major low molecular weight thiol in actinomycetes. Nat. Prod. Rep. 2008, 25, 1091–1117. [Google Scholar] [CrossRef]
- Beck, C.; Gren, T.; Jørgensen, T.S.; González, I.; Román-Hurtado, F.; Oves-Costales, D.; Genilloud, O.; Weber, T. Complete genome sequence of sp. strain CA-256286. Microbiol. Resour. Announc. 2021, 10, e0029021. [Google Scholar] [CrossRef] [PubMed]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef] [PubMed]
- Alanjary, M.; Steinke, K.; Ziemert, N. AutoMLST: An automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res. 2019, 47, W276–W282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.W.; Bibb, M.J.; Revill, W.P.; Hopwood, D.A. Cloning, sequencing, and analysis of the griseusin polyketide synthase gene cluster from Streptomyces griseus. J. Bacteriol. 1994, 176, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [Green Version]
- Palazzotto, E.; Weber, T. Omics and multi-omics approaches to study the biosynthesis of secondary metabolites in microorganisms. Curr. Opin. Microbiol. 2018, 45, 109–116. [Google Scholar] [CrossRef]
- Hilker, R.; Stadermann, K.B.; Doppmeier, D.; Kalinowski, J.; Stoye, J.; Straube, J.; Winnebald, J.; Goesmann, A. ReadXplorer—Visualization and analysis of mapped sequences. Bioinformatics 2014, 30, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilker, R.; Stadermann, K.B.; Schwengers, O.; Anisiforov, E.; Jaenicke, S.; Weisshaar, B.; Zimmermann, T.; Goesmann, A. ReadXplorer 2-detailed read mapping analysis and visualization from one single source. Bioinformatics 2016, 32, 3702–3708. [Google Scholar] [CrossRef]
- Tong, Y.; Whitford, C.M.; Blin, K.; Jørgensen, T.S.; Weber, T.; Lee, S.Y. CRISPR-Cas9, CRISPRi and CRISPR-BEST-mediated genetic manipulation in streptomycetes. Nat. Protoc. 2020, 15, 2470–2502. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Tong, Y.; Weber, T. Designing sgRNAs for CRISPR-BEST base editing applications with CRISPy-web 2.0. Synth. Syst. Biotechnol. 2020, 5, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Rao, R.N.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef]
- Sosio, M.; Giusino, F.; Cappellano, C.; Bossi, E.; Puglia, A.M.; Donadio, S. Artificial chromosomes for antibiotic-producing actinomycetes. Nat. Biotechnol. 2000, 18, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.C.; Gust, B.; Kulik, A.; Heide, L.; Buttner, M.J.; Bibb, M.J. Phage p1-derived artificial chromosomes facilitate heterologous expression of the FK506 gene cluster. PLoS ONE 2013, 8, e69319. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-H.; Cha, C.-J.; Roe, J.-H. Identification of genes for mycothiol biosynthesis in Streptomyces coelicolor A3(2). J. Microbiol. 2006, 44, 121–125. [Google Scholar] [PubMed]
- Park, J.-H.; Roe, J.-H. Mycothiol regulates and is regulated by a thiol-specific antisigma factor RsrA and σRin Streptomyces coelicolor. Mol. Microbiol. 2008, 68, 861–870. [Google Scholar] [CrossRef]
- Hopwood, D.A. Genetic contributions to understanding polyketide synthases. Chem. Rev. 1997, 97, 2465–2498. [Google Scholar] [CrossRef]
- Hertweck, C.; Luzhetskyy, A.; Rebets, Y.; Bechthold, A. Type II polyketide synthases: Gaining a deeper insight into enzymatic teamwork. Nat. Prod. Rep. 2007, 24, 162–190. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T.; Maltby, D.A.; Medzihradszky, K.F.; Khosla, C.; Stroud, R.M. An antibiotic factory caught in action. Nat. Struct. Mol. Biol. 2004, 11, 888–893. [Google Scholar] [CrossRef]
- Hong, L.; Zhao, Z.; Melançon, C.E.; Zhang, H.; Liu, H.-W. In vitro characterization of the enzymes involved in TDP-d-forosamine biosynthesis in the spinosyn pathway of Saccharopolyspora spinosa. J. Am. Chem. Soc. 2008, 130, 4954–4967. [Google Scholar] [CrossRef] [Green Version]
- Kormanec, J.; Novakova, R.; Mingyar, E.; Feckova, L. Intriguing properties of the angucycline antibiotic auricin and complex regulation of its biosynthesis. Appl. Microbiol. Biotechnol. 2014, 98, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Novakova, R.; Rehakova, A.; Kutas, P.; Feckova, L.; Kormanec, J. The role of two SARP family transcriptional regulators in regulation of the auricin gene cluster in Streptomyces aureofaciens CCM 3239. Microbiology 2011, 157, 1629–1639. [Google Scholar] [CrossRef] [Green Version]
- Bekeova, C.; Rehakova, A.; Feckova, L.; Vlckova, S.; Novakova, R.; Mingyar, E.; Kormanec, J. Characterisation of the genes involved in the biosynthesis and attachment of the aminodeoxysugar D-forosamine in the auricin gene cluster of Streptomyces aureofaciens CCM3239. Appl. Microbiol. Biotechnol. 2016, 100, 3177–3195. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.L.M.; Chooi, Y.-H. clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar]
- Medema, M.H.; Kottmann, R.; Yilmaz, P.; Cummings, M.; Biggins, J.B.; Blin, K.; de Bruijn, I.; Chooi, Y.H.; Claesen, J.; Coates, R.C.; et al. Minimum information about a biosynthetic gene cluster. Nat. Chem. Biol. 2015, 11, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework to explore large-scale biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Weber, T.; Medema, M.H. BiG-FAM: The biosynthetic gene cluster families database. Nucleic Acids Res. 2021, 49, D490–D497. [Google Scholar] [CrossRef]
- Bilyk, B.; Luzhetskyy, A. Unusual site-specific DNA integration into the highly active pseudo-attB of the Streptomyces albus J1074 genome. Appl. Microbiol. Biotechnol. 2014, 98, 5095–5104. [Google Scholar] [CrossRef]
- Martín, J.; Crespo, G.; González-Menéndez, V.; Pérez-Moreno, G.; Sánchez-Carrasco, P.; Pérez-Victoria, I.; Ruiz-Pérez, L.M.; González-Pacanowska, D.; Vicente, F.; Genilloud, O.; et al. MDN-0104, an antiplasmodial betaine lipid from Heterospora chenopodii. J. Nat. Prod. 2014, 77, 2118–2123. [Google Scholar] [CrossRef]
- Martín, J.; da S Sousa, T.; Crespo, G.; Palomo, S.; González, I.; Tormo, J.R.; de la Cruz, M.; Anderson, M.; Hill, R.T.; Vicente, F.; et al. Kocurin, the true structure of PM181104, an anti-methicillin-resistant Staphylococcus aureus (MRSA) thiazolyl peptide from the marine-derived bacterium Kocuria palustris. Mar. Drugs 2013, 11, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
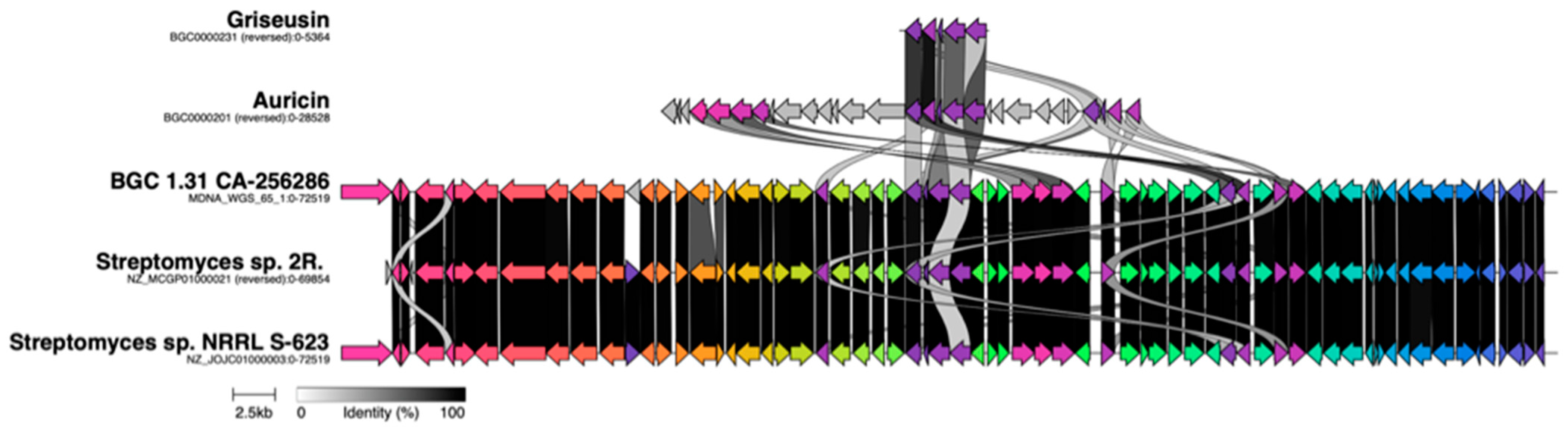{kind=link}
| Class of Transcriptional Regulator | Transcriptional Regulator Genes | Plasmid Name |
|---|---|---|
| Cluster specific regulators | aur1P-pntR-strR-fkbN | pRM4-CSRs/pEM1 [7] |
| Streptomyces antibiotic regulatory proteins | actIIORF4-griR-aur1PR3-papR2-redD | pRM4-SARPs/pEM2 [7] |
| Gamma butyrolactone synthases | scbA-afsA | pRM4-GBLs |
| Global regulators | afsR-adpA-crp-absB-dasR | pRM4-GRs |
| Position No. | δC, Mult | δH (Mult *, J in Hz) |
|---|---|---|
| 1 | 95.6, C | |
| 3 | 69.8, CH | 4.74, d (5.0) |
| 4 | 81.9, C | |
| 4a | 141.2, C | |
| 5 | 181.3, C | |
| 5a | 131.4, C | |
| 6 | 118.9, CH | 7.60, dd (7.4, 1.5) |
| 7 | 137.4, CH | 7.82, dd (8.6, 7.4) |
| 8 | 125.3, CH | 7.43, dd (8.6, 1.5) |
| 9 | 160.8, C | |
| 9-OH | 11.72, br s | |
| 9a | 114.9, C | |
| 10 | 186.3, C | |
| 10a | 140.2, C | |
| 11 | 34.4, CH2 | 3.61, dd (18.1, 5.0)/2.59, d (18.1) |
| 12 | 174.1, C | |
| 3′ | 70.9, CH | 4.78 a, d (4.2) |
| 4′ | 65.1 a, CH | 5.49, q (3.3) |
| 4′-OCOCH3 | 169.9, C | |
| 4′-OCOCH3 | 20.8, CH3 | 2.01, s |
| 5′ | 35.5, CH2 | H-5′eq:1.89, m/H-5′ax: 1.84, m |
| 6′ | 62.8, CH | 4.22, m |
| 7′ | 20.2, CH3 | 1.18, d (6.3) |
| 1″ | 93,7, CH | 4.76 a, br s |
| 2″ | 28.7, CH2 | 1.53, m; 1.40, m |
| 3″ | 14.3 | 1.61, m; 1.38, m |
| 4″ | 64.4 | 2.57 a, m |
| 4″-N(CH3)2 | 40.4, 2 × CH3 | 2.54 a, s |
| 5″ | 65.1 a, CH | 3.19, m |
| 6″ | 17.3, CH3 | 0.47, d (6.2) |
| 1″′ | 171.6, C | |
| 2″′ | 51.5, CH | 4.37, ddd (8.5, 8.2, 5.0) |
| 3″′ | 32.6, CH2 | 3.33, dd (13.5, 5.0)/2.94, dd (13.5, 8.5) |
| 2″′-NH | 8.22, d (8.2) | |
| 2″′-NHCOCH3 | 169.3, C | |
| 2″′-NHCOCH3 | 22.1, CH3 | 1.67, s |
| Gene Name | Origin of the Gene | Best BLAST Hit Score | Pairwise Identity, % | E Value | Position, nt | BGC and Gene Locus Tag |
|---|---|---|---|---|---|---|
| Ketosynthase CDS from X77865.1: CAA54858.1 | S. griseus K-63 | 1061.67 682.96 | 78.8% 73.4% | 0 0 | 3,472,656–3,471,404 6,876,549–6,875,389 | BGC 1.20 FBHECJPB_03028 BGC 1.31 FBHECJPB_06072 |
| Putative chain length determination factor CDS from X77865.1: CAA54859.1 | S. griseus K-63 | 590.988 365.567 | 72.4% 69.1% | 1.17 × 10168 8.44 × 10101 | 3,471,265–3,470,151 6,875,245–6,874,135 | BGC 1.20 FBHECJPB_03027 BGC 1.31 FBHECJPB_06071 |
| Acyl carrier protein CDS from X77865.1: CAA54860.1 | S. griseus K-63 | 206.87 62.6008 | 82.4% 68.8% | 1.01 × 1053 2.72 × 1010 | 3,470,076–3,469,873 6,874,057–6,873,904 | BGC 1.20 FBHECJPB_03026 BGC 1.31 FBHECJPB_06070 |
| Ketoreductase CDS from X77865.1: CAA54861.1 | S. griseus K-63 | 668.533 780.342 | 79.2% 82.6% | 0 0 | 6,893,267–6,892,495 3,469,673–3,468,908 | BGC 1.31 FBHECJPB_06088 BGC 1.20 FBHECJPB_03025 |
| Putative cyclase/dehydratase CDS from X77865.1: CAA54862.1 | S. griseus K-63 | 746.078 625.252 | 78.2% 75.4% | 0 4.37 × 10179 | 3,468,839–3,467,912 6,892,412–6,891,493 | BGC 1.20 FBHECJPB_03024 BGC 1.31 FBHECJPB_06087 |
| Gene Name | Origin of the Gene | Best BLAST Hit Score | Pairwise Identity, % | E Value | Position, nt | BGC and Locus Tags |
|---|---|---|---|---|---|---|
| mshA (SCO4204) | S. coelicolor A3(2) | 1296.62 | 85.2% | 0 | 3,445,621–3,444,352 No other good hits | BGC 1.20 FBHECJPB_03001 |
| mshB (SCO5126) | S. coelicolor A3(2) | 431.39 75.224 | 76.5% 73.4% | 9.22 × 10121 1.51 × 1013 | 2,457,013–2,456,142 2,642,174–2,642,051 | Close to BGC 1.18 FBHECJPB_02090 Close to BGC 1.18 FBHECJPB_02245 |
| mshC (SCO1663) | S. coelicolor A3(2) | 1362.1 118.505 | 86.7% 70.4% | 0 1.98 × 1026 | 6,462,580–6,463,808 3,428,519–3,428,260 | Not part of any BGC FBHECJPB_05696 Close to BGC 1.20 FBHECJPB_02982 |
| mshD (SCO4151) | S. coelicolor A3(2) | 780.342 | 79.8% | 0 | 3,532,239–3,533,119 No other good hits | Close to BGC 1.20 FBHECJPB_03096 |
| Mca (SCO4967) | S. coelicolor A3(2) | 1126.59 | 88.3% | 0 | 2,642,188–2,641,307 | Close to BGC 1.18 FBHECJPB_02245 |
| Protein Name | Origin of the Protein | Best BLAST Hit Score | Pairwise Identity, % | E Value | Position, nt | BGC and Locus Tag |
|---|---|---|---|---|---|---|
| SpnR | Saccharopolyspora spinosa NRRL 18537 | 503.056 168.318 | 63.4% 33.4% | 1.43 × 10160 3.58 × 1045 | 6,893,551–6,894,681 5,334,768–5,335,814 | BGC 1.31 FBHECJPB_06089 Not part of any BGC |
| SpnQ | Saccharopolyspora spinosa NRRL 18537 | 625.165 141.739 | 68.6% 30.3% | 0 6.48 × 1036 | 6,881,515–6,882,825 5,334,753–5,335,826 | BGC 1.31 FBHECJPB_06078 Not part of any BGC |
| SpnO | Saccharopolyspora spinosa NRRL 18537 | 409.068 | 48.2% | 2.11 × 10126 | 6,879,120–6,880,454 | BGC 1.31 FBHECJPB_06076 |
| SpnN | Saccharopolyspora spinosa NRRL 18537 | 265.003 | 50.8% | 7.04 × 1079 | 6,880,460–6,881,404 | BGC 1. 31 FBHECJPB_06077 |
| SpnS | Saccharopolyspora spinosa NRRL 18537 | 219.164 | 46.5% | 3.11 × 1064 | 6,871,449–6,870,763 | BGC 1.31 FBHECJPB_06067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beck, C.; Gren, T.; Ortiz-López, F.J.; Jørgensen, T.S.; Carretero-Molina, D.; Martín Serrano, J.; Tormo, J.R.; Oves-Costales, D.; Kontou, E.E.; Mohite, O.S.; et al. Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods. Molecules 2021, 26, 6580. https://doi.org/10.3390/molecules26216580
Beck C, Gren T, Ortiz-López FJ, Jørgensen TS, Carretero-Molina D, Martín Serrano J, Tormo JR, Oves-Costales D, Kontou EE, Mohite OS, et al. Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods. Molecules. 2021; 26(21):6580. https://doi.org/10.3390/molecules26216580
Chicago/Turabian StyleBeck, Charlotte, Tetiana Gren, Francisco Javier Ortiz-López, Tue Sparholt Jørgensen, Daniel Carretero-Molina, Jesús Martín Serrano, José R. Tormo, Daniel Oves-Costales, Eftychia E. Kontou, Omkar S. Mohite, and et al. 2021. "Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods" Molecules 26, no. 21: 6580. https://doi.org/10.3390/molecules26216580
APA StyleBeck, C., Gren, T., Ortiz-López, F. J., Jørgensen, T. S., Carretero-Molina, D., Martín Serrano, J., Tormo, J. R., Oves-Costales, D., Kontou, E. E., Mohite, O. S., Mingyar, E., Stegmann, E., Genilloud, O., & Weber, T. (2021). Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods. Molecules, 26(21), 6580. https://doi.org/10.3390/molecules26216580