
Extraction and Purification of R-Phycoerythrin Alpha Subunit from the Marine Red Algae Pyropia Yezoensis and Its Biological Activities


Abstract
1. Introduction
2. Results and Discussion
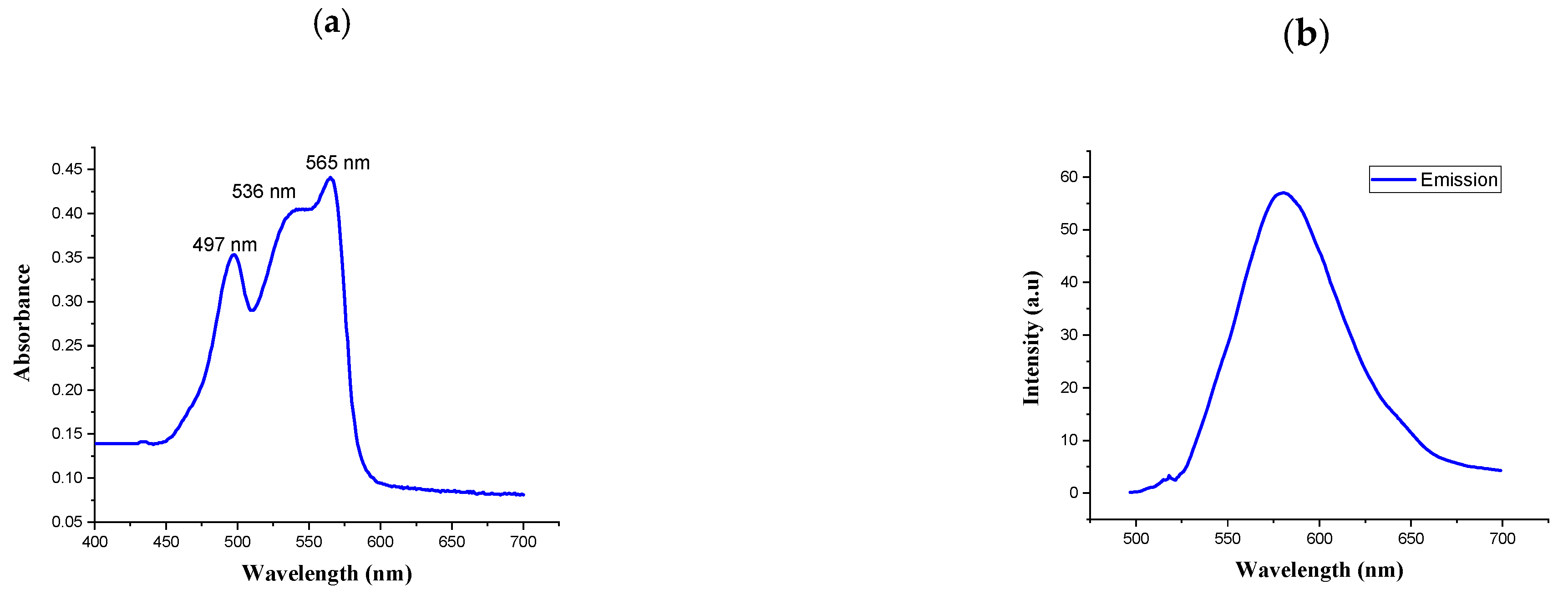
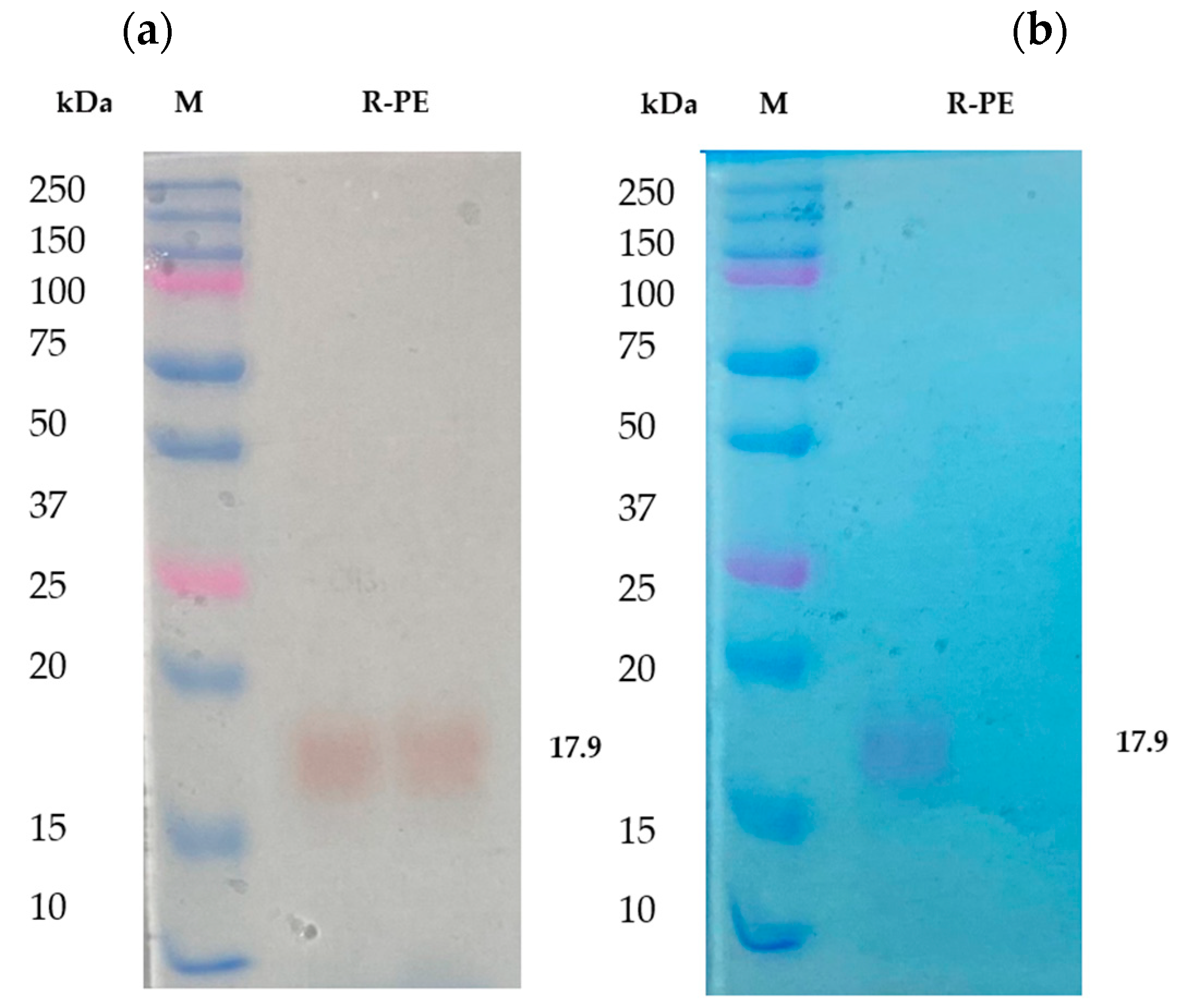
2.1. Isolation and Purification of R-Phycoerythrin Alpha Subunit from Pyropia Yezoensis
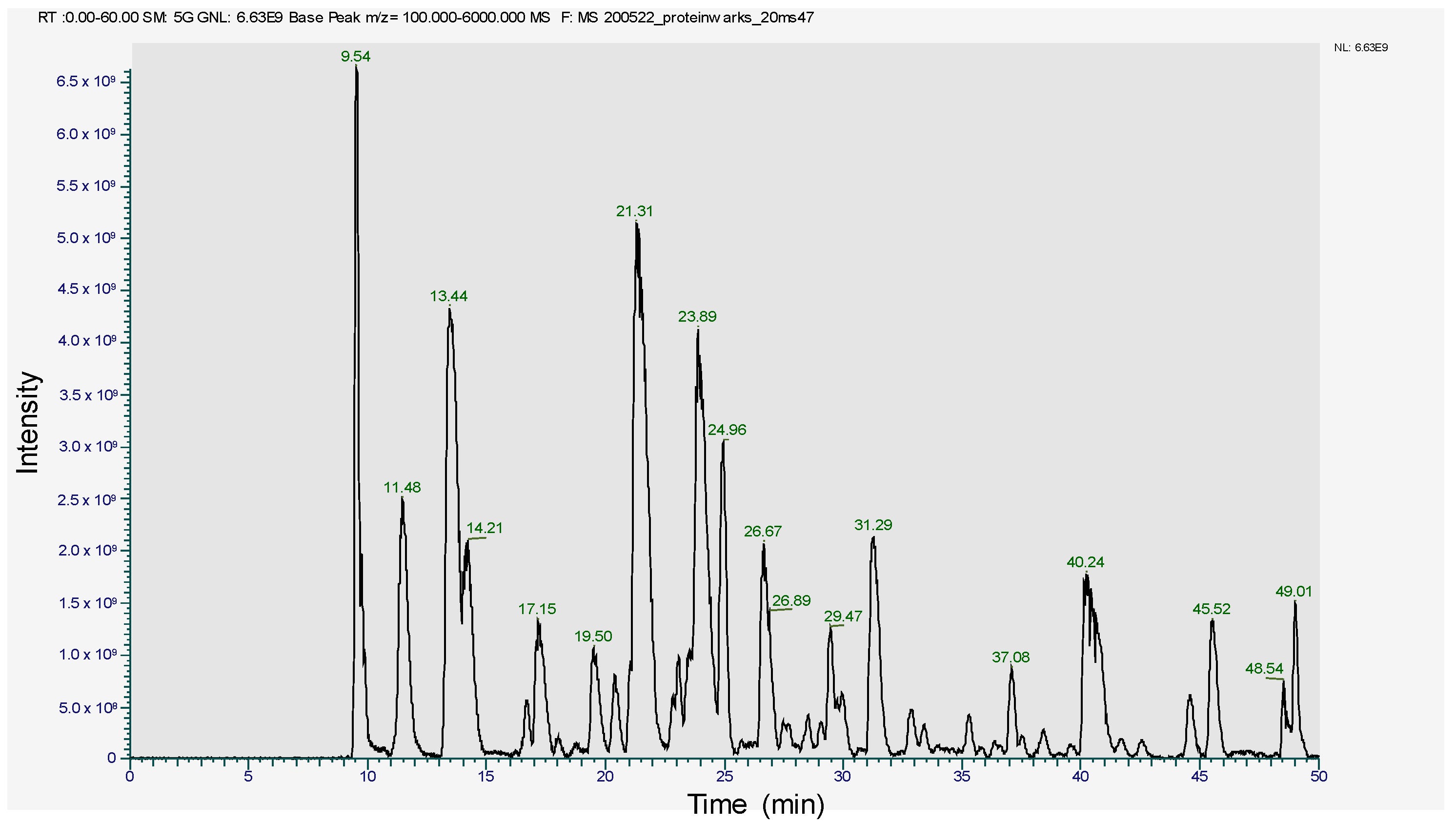
2.2. MS2 In-Gel Digestion and Protein ID by LC-MS/MS
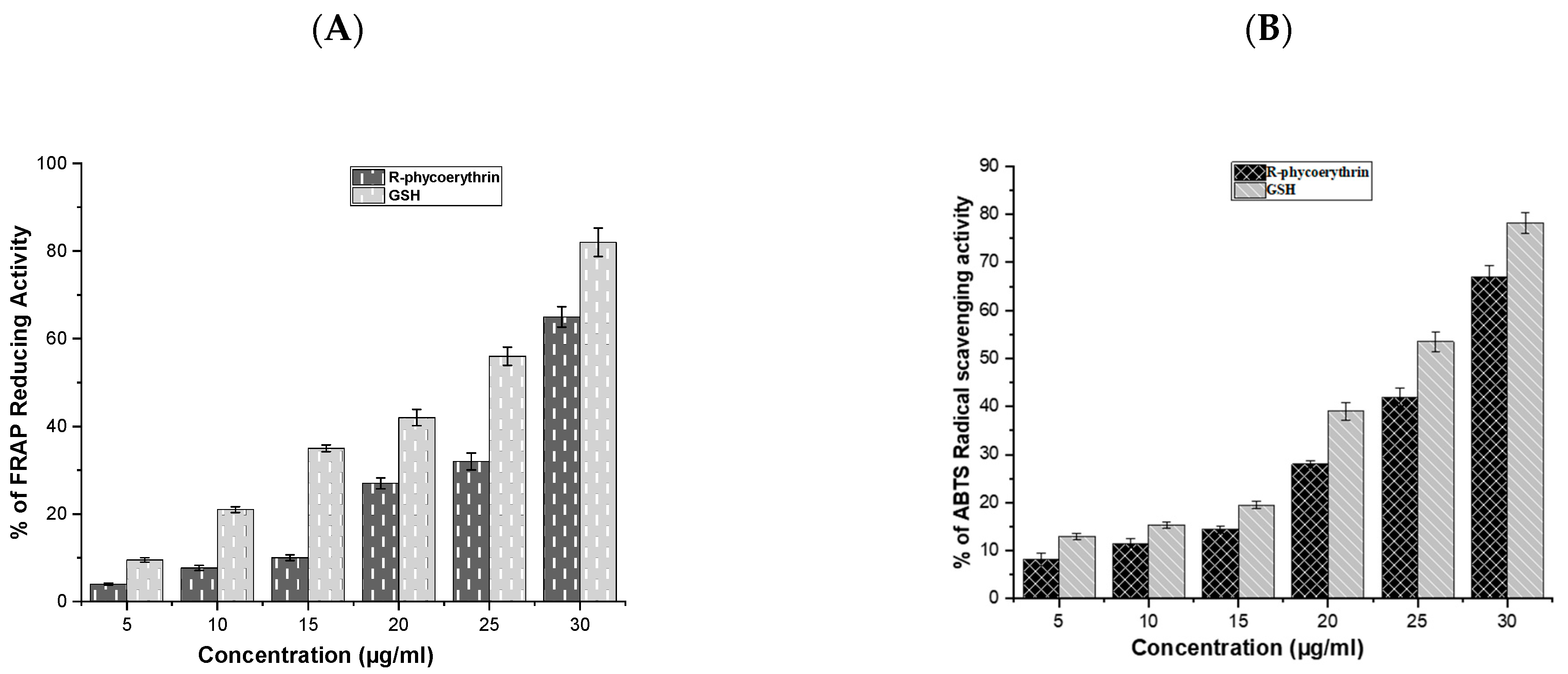
2.3. Antioxidant Activity
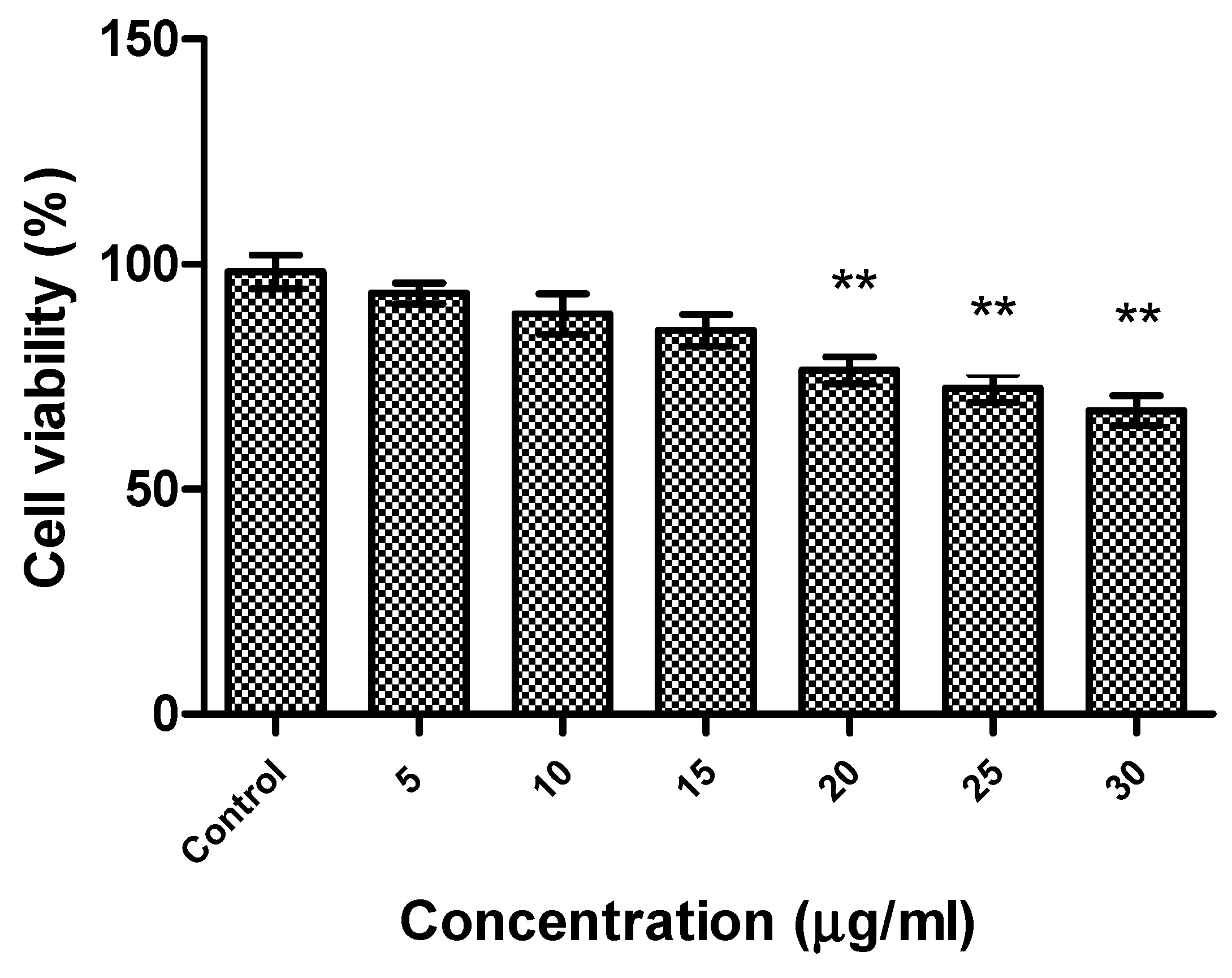
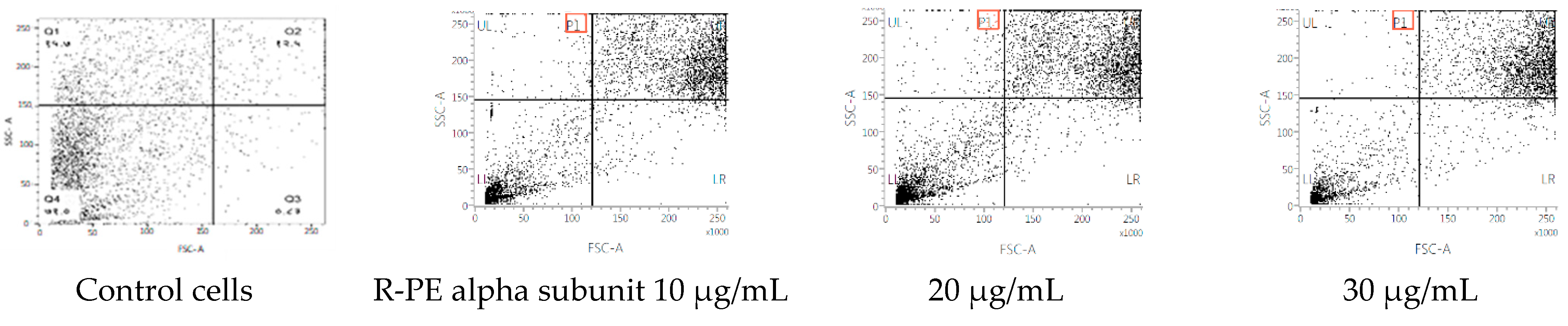
2.4. Cytotoxicity of R-Phycoerythrin Alpha Subunit
3. Materials and Methods
3.1. Chemicals
3.2. Preparation of Pyropia Yezoensis Extract
3.3. Purification
3.4. MS2 In-Gel Digestion and Protein Identification by Automatic LC-MS/MS
3.5. Antioxidant Activity
3.5.1. ABTS Assay
3.5.2. Ferric-Reducing Antioxidant Power (FRAP) Assay
3.6. Cell Culture
3.7. MTS Assay
3.8. Flow Cytometry Analyses
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F.; Global Cancer Observatory: Cancer Today. Lyon: International Agency for Research on Cancer 2020. Available online: https://gco.iarc.fr/today (accessed on 21 February 2021).
- Shaaban, S.; Negm, A.; Ibrahim, E.E.; Elrazak, A.A. Chemotherapeutic agents for the treatment of hepatocellular carcinoma: Efficacy and mode of action. Oncol. Rev. 2014, 8, 246. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pangestuti, R.; Kim, S. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed]
- Gantar, M.; Dhandayuthapani, S.; Rathinavelu, A. Phycocyanin Induces Apoptosis and Enhances the Effect of Topotecan on Prostate Cell Line LNCaP 1. J. Med. Food 2012, 15, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Qian, Z.-J.; Byun, H.-G.; Kim, S.-K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, L.; Kokot, S. Voltammetric determination of butylated hydroxyanisole, butylated hydroxytoluene, propyl gallate and tert -butylhydroquinone by use of chemometric approaches. Anal. Chim. Acta 2000, 412, 185–193. [Google Scholar] [CrossRef]
- Cornish, M.L.; Garbary, D.J. Antioxidants from macroalgae: Potential applications in human health and nutrition. Algae 2010, 25, 155–171. [Google Scholar] [CrossRef]
- Borzelleca, J.F.; Capen, C.C.; Hallagan, J.B. Lifetime toxicity/carcinogenicity study of fd & c red no. 3 (erythrosine) in rats. Food Chem. Toxic. 1987, 25, 723–733. [Google Scholar]
- Mahmoud, N.H. Toxic effects of the synthetic food dye brilliant blue on liver, kidney and testes functions in rats. J. Egypt. Soc. Toxicol. 2006, 34, 77–84. [Google Scholar]
- Hashem, M.M.; Abd-elhakim, Y.M.; Abo-el-sooud, K.; Eleiwa, M.M.E. Embryotoxic and Teratogenic Effects of Tartrazine in Rats. Toxicol. Res. 2019, 35, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Shetty, M.J.; Chandran, M. Natural Pigments as Potential Food Colourants: A Review Natural Pigments as Potential Food Colourants: A Review. Biosci. Trends 2017, 10, 4057–4064. [Google Scholar]
- Selvam, R.M.; Athinarayanan, G.; Raja, A.U.; Singh, A.J.A.; Kalirajan, K.; Selvakumar, P.M. Extraction of natural dyes from Curcuma longa, Trigonella foenum graecum and Nerium oleander, plants and their application in antimicrobial fabric. Ind. Crop. Prod. 2015, 70, 84–90. [Google Scholar] [CrossRef]
- Mirjalili, M.; Nazarpoor, K.; Karimi, L. Eco-friendly dyeing of wool using natural dye from weld as co-partner with synthetic dye. J. Clean. Prod. 2011, 19, 1045–1051. [Google Scholar] [CrossRef]
- Frick, D. The coloration of food. Color. Technol. 2003, 33, 15–32. [Google Scholar] [CrossRef]
- Dweck, A.C. Natural ingredients for colouring and styling. Int. J. Cosmet. Sci 2002, 24, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Shabbir, M.; Mohammad, F. Natural Colorants: Historical, Processing and Sustainable Prospects. Nat. Prod. Bioprospect. 2017, 7, 123–145. [Google Scholar] [CrossRef] [PubMed]
- Gamal, A.A. El Biological importance of marine algae. Saudi Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Kuddus, M.; Singh, P.; Thomas, G.; Al-hazimi, A. Recent Developments in Production and Biotechnological Applications of C-Phycocyanin. BioMed Res. Int. 2013, 2013, 742859. [Google Scholar] [CrossRef]
- Dumay, J.; Morançais, M.; Munier, M.; Cecile, G.; Fleurence, J. Phycoerythrins: Valuable Proteinic Pigments in Red Seaweeds. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, pp. 321–343. [Google Scholar]
- Maccoll, R. Cyanobacterial Phycobilisomes. J. Struct. Biol. 1998, 334, 311–334. [Google Scholar] [CrossRef]
- Sidler, W. Phycobilisome and Phycobiliprotein Structures. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; Volume 2, ISBN 978-0-7923-3273-2. [Google Scholar]
- Fernandez, M.; Bermejo, R.; Acien, F.G.; Molina, E. Preparative purification of B-phycoerythrin from the microalga Porphyridium cruentum by expanded-bed adsorption chromatography. J. Chromatogr. B 2003, 790, 317–325. [Google Scholar]
- Sathuvan, M.; Sakthivel, M.; Babu, G.V.; Palani, P.; Rengasamy, R. Qualitative and quantitative determination of R-Phycoerythrin from Halymenia floresia (Clemente) C. Agardh by Polyacrylamide Gel Using Electrophoretic elution technique. J. Chromatogr. A 2016, 1454, 120–126. [Google Scholar]
- Rossano, R.; Liuzzi, G.M.; Riccio, P. Extracting and purifying R-phycoerythrin from Mediterranean red Extracting and purifying R-phycoerythrin from Mediterranean red algae Corallina elongata Ellis & Solander. J. Biotechnol. 2003, 1656, 2–6. [Google Scholar]
- Wang, L.; Wang, S.; Fu, X.; Sun, L. Characteristics of an R-Phycoerythrin with Two γ Subunits Prepared from Red Macroalga Polysiphonia urceolata. PLoS ONE 2015, 10, e0120333. [Google Scholar] [CrossRef]
- Blouin, N.A.; Brodie, J.A.; Grossman, A.C.; Xu, P.; Brawley, S.H. Porphyra: A marine crop shaped by stress. Trends Plant Sci. 2011, 16, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Tae, J.C.; Min, S.R. Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed. Mar. Drugs 2020, 18, 14. [Google Scholar] [CrossRef]
- Ismail, M.M.; Alotaibi, B.S.; EL-Sheekh, M.M. Therapeutic Uses of Red Macroalgae. Molecules 2020, 25, 4411. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Gaigalas, A.; Gallagher, T.; Cole, K.D.; Singh, T.; Wang, L.; Zhang, Y.Z. A multistate model for the fluorescence response of r-phycoerythrin. Photochem. Photobiol. 2006, 82, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Punampalam, R.; Soo, K.; Weng, N. Evaluation of antioxidant properties of phycobiliproteins and phenolic compounds extracted from Bangia atropurpurea. Malays. J. Fundam. Appl. Sci. 2018, 14, 289–297. [Google Scholar] [CrossRef]
- Niu, J.; Wang, G.; Tseng, C. Method for large-scale isolation and purification of R-phycoerythrin from red alga Polysiphonia urceolata Grev. Protein Expr. Purif. 2006, 49, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, N.; Kurinjimalar, C.; Thangam, R. Further studies and biological activities of macromolecular protein R-Phycoerythrin from Portieria hornemannii. Int. J. Biol. Macromol. 2013, 62, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Tajbakhsh, S.; Nabipour, I.; Rastian, Z.; Yousefi, F.; Sharafian, S.; Sartavi, K. In vitro antitumor activity of Gracilaria corticata (a red alga) against Jurkat and molt-4 human cancer cell lines. Afr. J. Biotechnol. 2010, 9, 6787–6790. [Google Scholar]
- Yifeng, X.; Quanfu, W.; Yanhua, H. Efficient Purification of R-phycoerythrin from Marine Algae (Porphyra yezoensis) Based on a Deep Eutectic Solvents Aqueous Two-Phase System. Mar. Drugs 2020, 18, 618. [Google Scholar] [CrossRef]
- Denis, C.; Massé, A.; Fleurence, J.; Jaouen, P. Concentration and pre-purification with ultrafiltration of a R-phycoerythrin solution extracted from macro-algae Grateloupia turuturu: Process definition and up-scaling. Sep. Purif. Technol. 2009, 69, 37–42. [Google Scholar] [CrossRef]
- Munier, M.; Jubeau, S.; Wijaya, A.; Morançais, M.; Dumay, J.; Marchal, L.; Jaouen, P.; Fleurence, J. Physicochemical factors affecting the stability of two pigments: R-phycoerythrin of Grateloupia turuturu and B-phycoerythrin of Porphyridium cruentum. Food Chem. 2014, 150, 400–407. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.; Krishna, J.G.; Panikkar, M.V.N.; Nair, B.G.; Pai, J.G.; Nair, S.S. Identification, purification, biochemical and mass spectrometric characterization of novel phycobiliproteins from a marine red alga, Centroceras clavulatum. Int. J. Biol. Macromol. 2018, 114, 679–691. [Google Scholar] [CrossRef]
- Kelman, D.; Posner, E.K.; Mcdermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant Activity of Hawaiian Marine Algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef]
- Ngo, D.H.; Wijesekara, I.; Vo, T.S.; Van Ta, Q.; Kim, S.K. Marine food derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int. 2010, 44, 523–529. [Google Scholar] [CrossRef]
- Gülçin, I. Fe3+–Fe2+ transformation method: An important antioxidant assay. Methods Mol. Biol. 2015, 1208, 233–246. [Google Scholar] [CrossRef]
- Chu, W.-L.; Lim, Y.-W.; Radhakrishnan, A.K.; Lim, P.-E. Protective effect of aqueous extract from Spirulina platensis against cell death induced by free radicals. BMC Complement. Altern. Med. 2010, 10, 53. [Google Scholar] [CrossRef]
- Yasin, D.; Zafaryab, M.; Ansari, S.; Ahmad, N.; Khana, N.F.; Zaki, A.; Rizvi, M.M.A.; Fatma, T. Evaluation of antioxidant and anti-proliferative efficacy of Nostoc muscorum NCCU-442. Biocatal. Agric. Biotechnol. 2019, 17, 284–293. [Google Scholar] [CrossRef]
- Gholamreza, M.; Mohammad, F.; Habibollah, N. Survey of Antioxidant and Cytotoxic Activities of Gracilaria corticata (a Red Seaweed), Against RKO, AGS and HepG2 Human Cancer Cell Lines. J. Biol. Today’s World 2017, 6, 269–273. [Google Scholar]
- Shalaby, E.A.; Shanab, S.M.M. Antioxidant compounds, assays of determination and mode of action. Afr. J. Pharm. Pharmacol. 2013, 7, 528–539. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Roberta, R.E.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Koopman, G.; Reutelingsperger, C.P.M.; Kuijten, G.A.M.; Keehnen, R.M.J.; Pals, S.T.; Oers, M.H.J. Van Annexin V for Flow Cytometric Detection of Phosphatidylserine Expression on B Cells Undergoing Apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Sl.No | Identified Peptides | Enzyme Used | Charge State | Observe Mass (Da) | Calculated Mass (Da) | Software Used For Peptide Identification |
|---|---|---|---|---|---|---|---|
| R-Phycoerythrin alpha subunit | 1 | VNKCYR | Trypsin | 2 | 838.4112 | 838.4119 | Mascot |
| 2 | DRLCVPR | Trypsin | 2 | 914.4727 | 914.4756 | Mascot | |
| 3 | DVDHYMR | Trypsin | 2 | 934.3965 | 934.3967 | Mascot | |
| 4 | EAGDACFAK | Trypsin | 2 | 967.4037 | 967.4069 | Mascot | |
| 5 | AAARLEAAEK | Trypsin | 2 | 1028.558 | 1028.561 | Mascot | |
| 6 | LASNHEAVVK | Trypsin | 3 | 1066.574 | 1066.577 | Mascot | |
| 7 | NPGEAGDSQEK | Trypsin | 2 | 1130.484 | 1130.484 | Mascot | |
| 8 | CYRDVDHYMR | Trypsin | 4 | 1413.589 | 1413.592 | Mascot | |
| 9 | SVITTTISAADAAGR | Trypsin | 2 | 1432.753 | 1432.752 | Mascot | |
| 10 | NPGEAGDSQEKVNK | Trypsin | 3 | 1471.689 | 1471.69 | Mascot | |
| 11 | EAGDACFAKYSYLK | Trypsin | 2 | 1621.732 | 1621.745 | Mascot | |
| 12 | MKSVITTTISAADAAGR | Trypsin | 3 | 1691.889 | 1691.888 | Mascot | |
| 13 | LEAAEKLASNHEAVVK | Trypsin | 3 | 1707.914 | 1707.916 | Mascot | |
| 14 | FPSSSDLESVQGNIQR | Trypsin | 3 | 1762.848 | 1762.849 | Mascot | |
| 15 | VNKCYRDVDHYMR | Trypsin | 5 | 1770.792 | 1770.793 | Mascot | |
| 16 | Y*SYLKNPGEAGDSQEK | Trypsin | 2 | 1784.818 | 1784.822 | Mascot | |
| 17 | TLNLPTSAYVASFAFAR | Trypsin | 3 | 1827.945 | 1827.952 | Mascot | |
| 18 | NPGEAGDSQEKVNKCYR | Trypsin | 4 | 1950.876 | 1950.885 | Mascot | |
| 19 | LASNHEAVVKEAGDACFAK | Trypsin | 3 | 2015.969 | 2015.974 | Mascot | |
| 20 | AAARLEAAEKLASNHEAVVK | Trypsin | 4 | 2077.131 | 2077.128 | Mascot | |
| 21 | TLNLPTSAYVASFAFARDR | Trypsin | 3 | 2099.083 | 2099.08 | Mascot | |
| 22 | YSYLKNPGEAGDSQEKVNK | Trypsin | 3 | 2126.024 | 2126.028 | Mascot | |
| 23 | EVYRTLNLPTSAYVASFAFAR | Trypsin | 4 | 2375.234 | 2375.227 | Mascot | |
| 24 | LVNYCLVVGGTGPVDEWGIAGAR | Trypsin | 3 | 2402.2 | 2402.205 | Mascot | |
| 25 | DMSAQAGVEYAGNLDYIINSLC | Trypsin | 2 | 2403.068 | 2403.072 | Mascot | |
| 26 | LEAAEKLASNHEAVVKEAGDACFAK | Trypsin | 3 | 2657.287 | 2657.312 | Mascot | |
| 27 | TLNLPTSAYVASFAFARDRLCVPR | Trypsin | 3 | 2724.424 | 2724.417 | Mascot | |
| 28 | EAGDACFAKYSYLKNPGEAGDSQEK | Trypsin | 3 | 2734.216 | 2734.218 | Mascot | |
| 29 | LCVPRDMSQAGVEYAGNLDYIINSLC | Trypsin | 3 | 3044.407 | 3044.404 | Mascot | |
| 30 | S*VITTTISAADAAGRFPSSSDLESVQGNIQR | Trypsin | 4 | 3177.58 | 3177.59 | Mascot | |
| 31 | KSVITTTISAADAAGRFPSSSDLESVQGNIQR | Trypsin | 3 | 3305.695 | 3305.685 | Mascot | |
| 32 | DRLCVPRDMSAQAGVEYAGNLDYIINSLC | Trypsin | 3 | 3315.529 | 3315.532 | Mascot | |
| 33 | D*VDHYMRLVNYCLVVGGTGPVDEWGIAGAR | Trypsin | 4 | 3319.586 | 3318.591 | Mascot | |
| 34 | MKSVITTTISAADAAGRFPSSSDLESVQGNIQR | Trypsin | 4 | 3436.702 | 3436.726 | Mascot |
| Accessions | Description | Organism | Prot_Score | MW | PI | Coverage |
|---|---|---|---|---|---|---|
| D6QTA5 | Phycoerythrin alpha subunit | Pyropia yezoensis | 30,804 | 17,972 | 5.40 | 100.0 |
| D6QTA4 | Phycoerythrin beta subunit | Pyropia yezoensis | 24,573 | 18,810 | 6.23 | 77.4 |
| Q1XDB0 | C-phycocyanin beta chain | Pyropia yezoensis | 3196 | 18,360 | 4.94 | 84.9 |
| M4QTP4 | Allophycocyanin alpha subunit | Pyropia yezoensis | 179 | 17,669 | 5.06 | 49.7 |
| Q1XDA9 | C-phycocyanin alpha chain | Pyropia yezoensis | 82 | 17,567 | 7.71 | 24.1 |
| M4QTY0 | Ribosomal protein L12 | Pyropia yezoensis | 46 | 13,752 | 4.61 | 15.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulagesan, S.; Nam, T.-J.; Choi, Y.-H. Extraction and Purification of R-Phycoerythrin Alpha Subunit from the Marine Red Algae Pyropia Yezoensis and Its Biological Activities. Molecules 2021, 26, 6479. https://doi.org/10.3390/molecules26216479
Ulagesan S, Nam T-J, Choi Y-H. Extraction and Purification of R-Phycoerythrin Alpha Subunit from the Marine Red Algae Pyropia Yezoensis and Its Biological Activities. Molecules. 2021; 26(21):6479. https://doi.org/10.3390/molecules26216479
Chicago/Turabian StyleUlagesan, Selvakumari, Taek-Jeong Nam, and Youn-Hee Choi. 2021. "Extraction and Purification of R-Phycoerythrin Alpha Subunit from the Marine Red Algae Pyropia Yezoensis and Its Biological Activities" Molecules 26, no. 21: 6479. https://doi.org/10.3390/molecules26216479
APA StyleUlagesan, S., Nam, T.-J., & Choi, Y.-H. (2021). Extraction and Purification of R-Phycoerythrin Alpha Subunit from the Marine Red Algae Pyropia Yezoensis and Its Biological Activities. Molecules, 26(21), 6479. https://doi.org/10.3390/molecules26216479