
Starch Granule Size and Morphology of Arabidopsis thaliana Starch-Related Mutants Analyzed during Diurnal Rhythm and Development


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
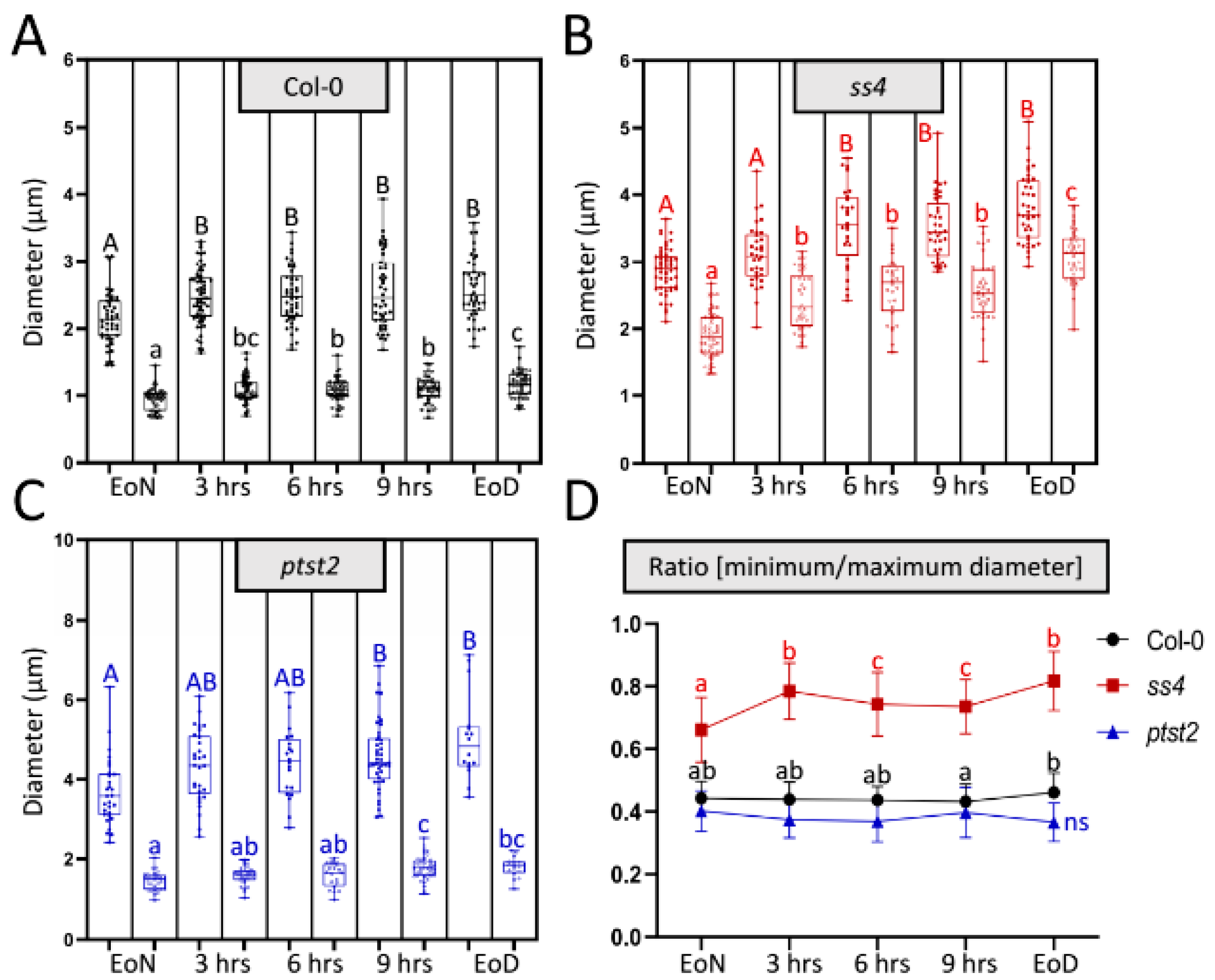
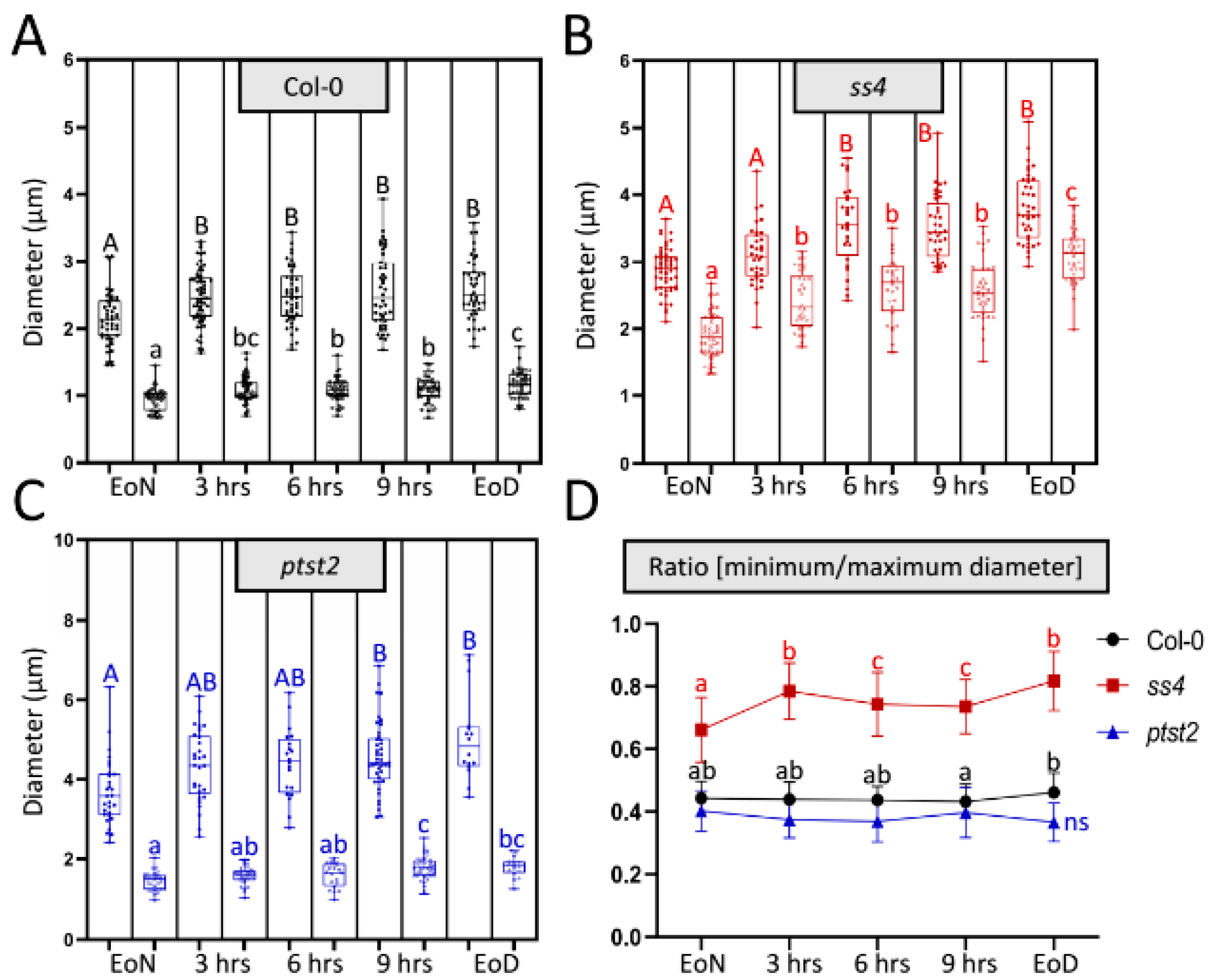
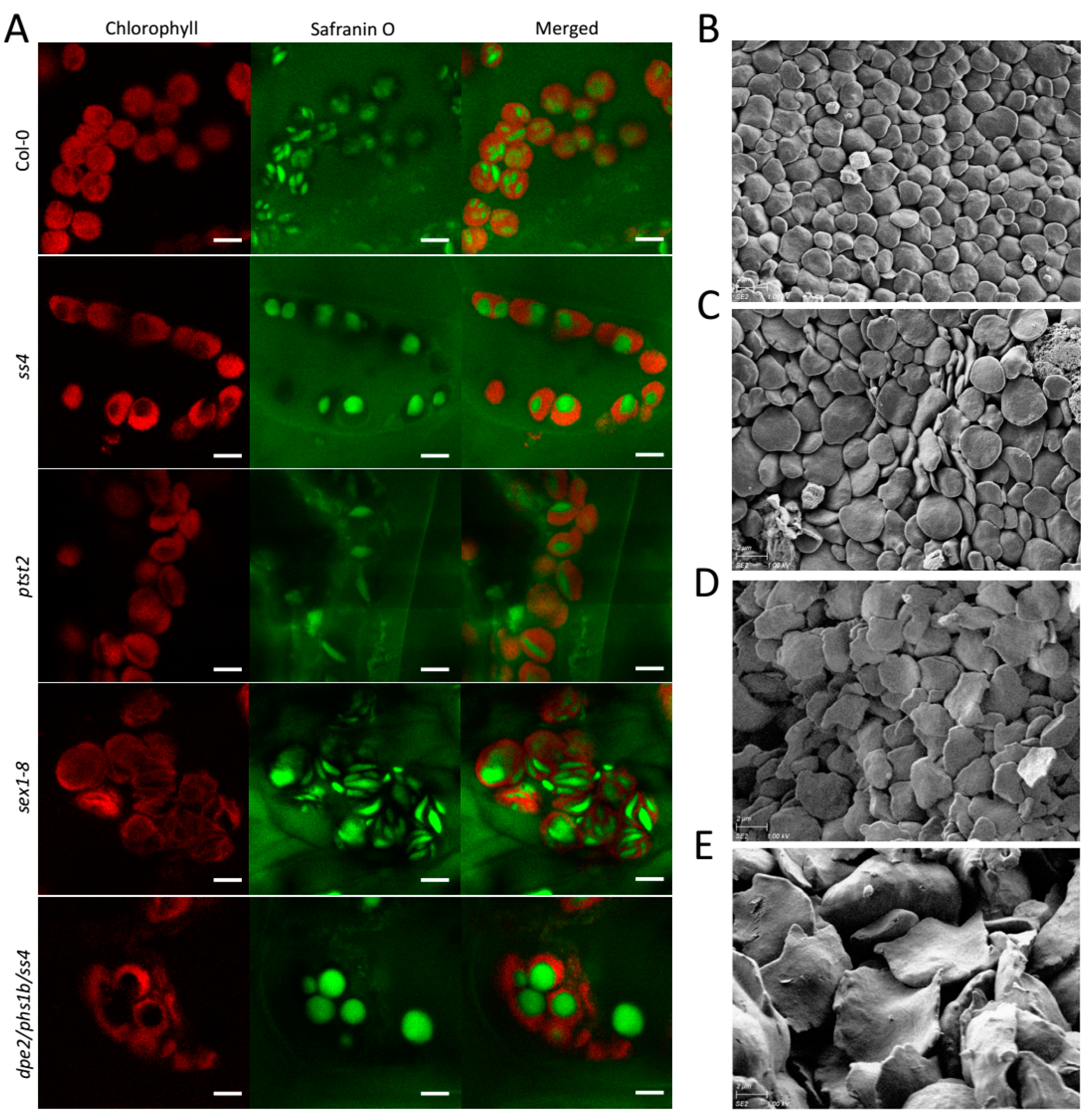
2.1. In Vivo Imaging by LCSM Can Track and Estimate Diurnal Starch Granule Size and Morphological Alterations
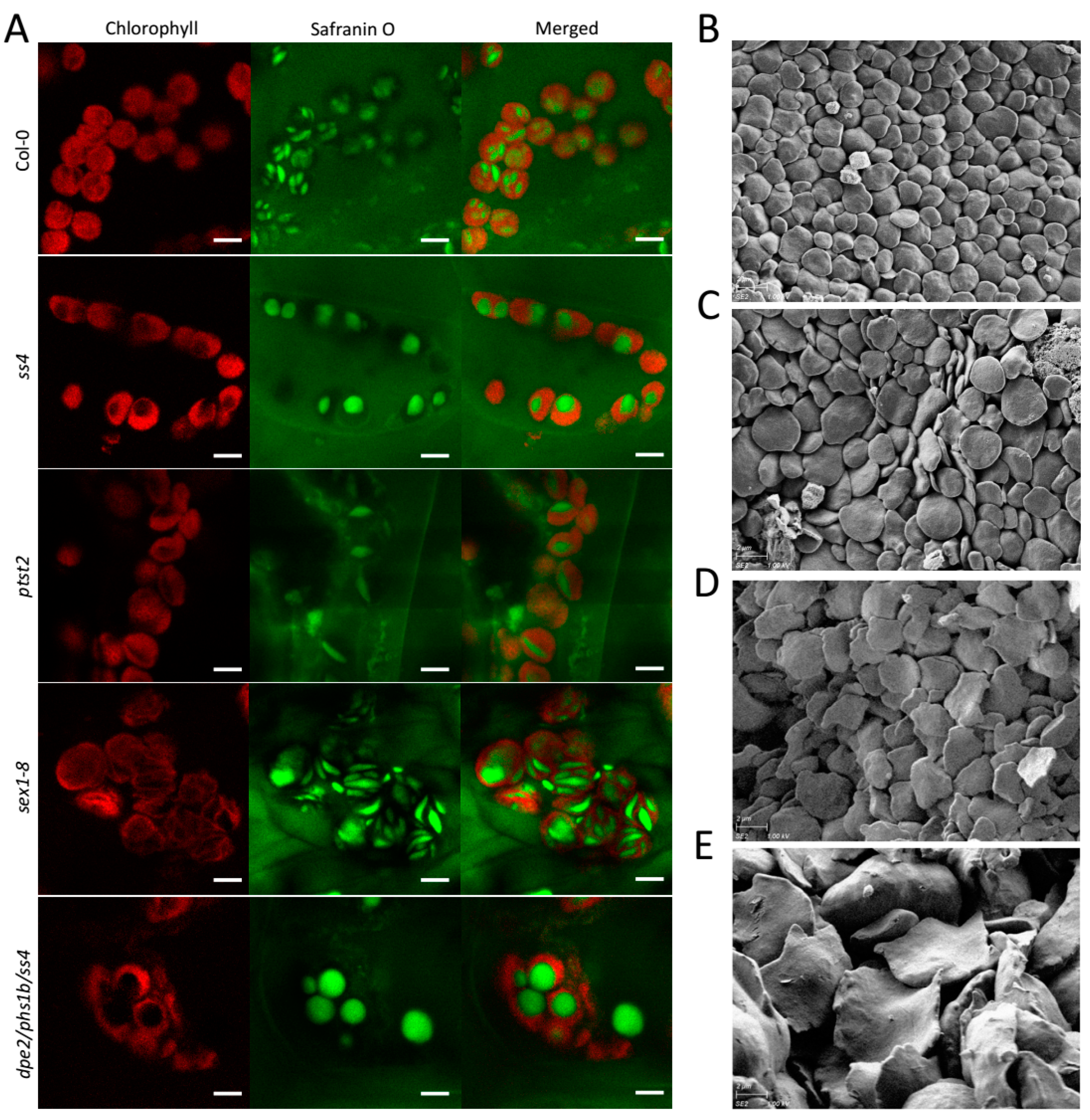
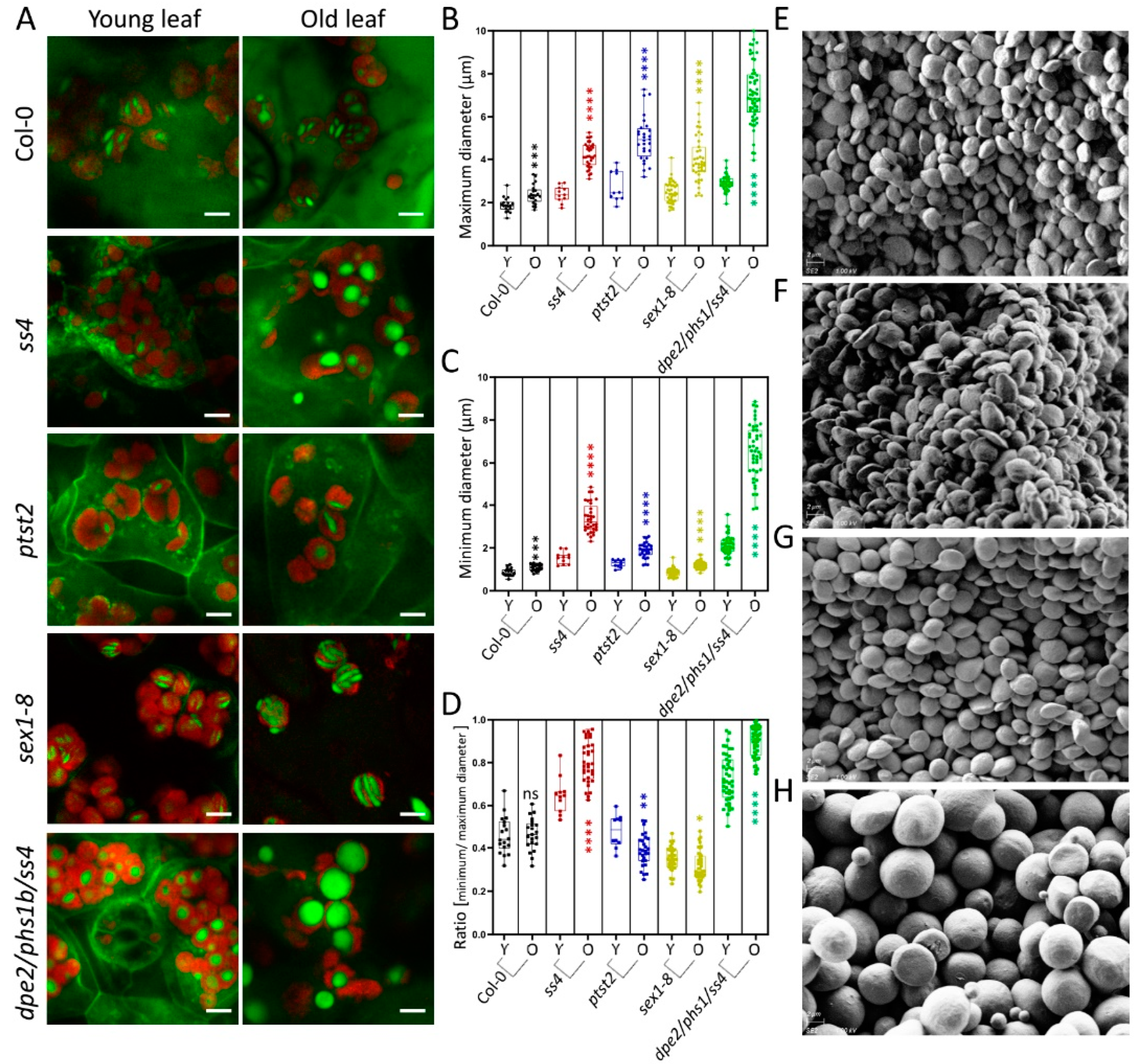
2.2. In Vivo Status of the Starch Granules in sex1-8 Indicates Alteration of Morphological Features during Leaf Aging
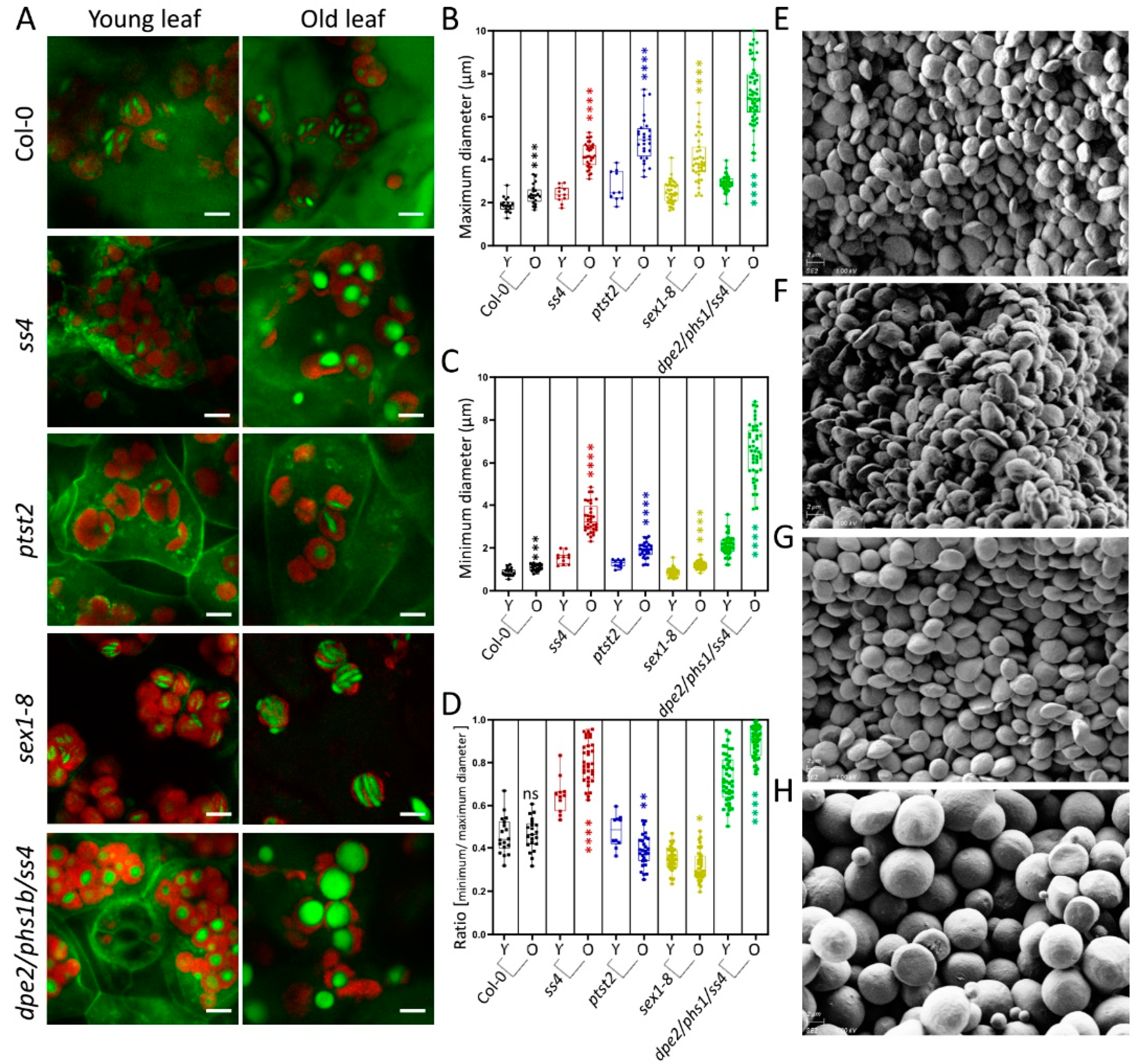
2.3. Starch Granule Size and Morphological Alterations during Leaf Aging in Various Starch-Related Mutants
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Safranin-O Staining and Laser Confocal Scanning Microscopy
4.3. Starch Granule Extraction
4.4. Scanning Electronic Microscopy
4.5. Native PAGE and Zymograms
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Tetlow, I.J.; Bertoft, E. A Review of Starch Biosynthesis in Relation to the Building Block-Backbone Model. Int. J. Mol. Sci. 2020, 21, 7011. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.M.; Zeeman, S.C. Starch: A Flexible, Adaptable Carbon Store Coupled to Plant Growth. Annu. Rev. Plant Biol. 2020, 71, 217–245. [Google Scholar] [CrossRef] [PubMed]
- Merida, A.; Fettke, J. Starch granule initiation in Arabidopsis thaliana chloroplasts. Plant J. 2021, 107, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Abt, M.R.; Zeeman, S.C. Evolutionary innovations in starch metabolism. Curr. Opin. Plant Biol. 2020, 55, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Malinova, I.; Qasim, H.M.; Brust, H.; Fettke, J. Parameters of Starch Granule Genesis in Chloroplasts of Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 761. [Google Scholar] [CrossRef] [PubMed]
- Roldán, I.; Wattebled, F.; Lucas, M.M.; Delvallé, D.; Planchot, V.; Jiménez, S.; Pérez, R.; Ball, S.; D’Hulst, C.; Mérida, Á. The phenotype of soluble starch synthase IV defective mutants of Arabidopsis thaliana suggests a novel function of elongation enzymes in the control of starch granule formation. Plant J. 2007, 49, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Seung, D.; Boudet, J.; Monroe, J.; Schreier, T.B.; David, L.C.; Abt, M.; Lu, K.-J.; Zanella, M.; Zeeman, S.C. Homologs of PROTEIN TARGETING TO STARCH Control Starch Granule Initiation in Arabidopsis Leaves. Plant Cell 2017, 29, 1657–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinova, I.; Fettke, J. Reduced starch granule number per chloroplast in the dpe2/phs1 mutant is dependent on initiation of starch degradation. PLoS ONE 2017, 12, e0187985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinova, I.; Mahlow, S.; Alseekh, S.; Orawetz, T.; Fernie, A.R.; Baumann, O.; Steup, M.; Fettke, J. Double Knockout Mutants of Arabidopsis Grown under Normal Conditions Reveal that the Plastidial Phosphorylase Isozyme Participates in Transitory Starch Metabolism. Plant Physiol. 2014, 164, 907–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Sharkey, T.D. The role of amylomaltase in maltose metabolism in the cytosol of photosynthetic cells. Planta 2004, 218, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Chia, T.; Thorneycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.C.; Smith, S.M.; Smith, A.M. A cytosolic glucosyltransferase is required for conversion of starch to sucrose in Arabidopsis leaves at night. Plant J. 2004, 37, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Fettke, J.; Chia, T.; Eckermann, N.; Smith, A.; Steup, M. A transglucosidase necessary for starch degradation and maltose metabolism in leaves at night acts on cytosolic heteroglycans (SHG). Plant J. 2006, 46, 668–684. [Google Scholar] [CrossRef] [PubMed]
- Malinova, I.; Alseekh, S.; Feil, R.; Fernie, A.R.; Baumann, O.; Schöttler, M.A.; Lunn, J.E.; Fettke, J. Starch Synthase 4 and Plastidal Phosphorylase Differentially Affect Starch Granule Number and Morphology. Plant Physiol. 2017, 174, 73–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlow, S.; Hejazi, M.; Kuhnert, F.; Garz, A.; Brust, H.; Baumann, O.; Fettke, J. Phosphorylation of transitory starch by α-glucan, water dikinase during starch turnover affects the surface properties and morphology of starch granules. New Phytol. 2014, 203, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Ritte, G.; Lloyd, J.; Eckermann, N.; Rottmann, A.; Kossmann, J.; Steup, M. The starch-related R1 protein is an -glucan, water dikinase. Proc. Natl. Acad. Sci. USA 2002, 99, 7166–7171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edner, C.; Li, J.; Albrecht, T.; Mahlow, S.; Hejazi, M.; Hussain, H.; Kaplan, F.; Guy, C.; Smith, S.M.; Steup, M.; et al. Glucan, water dikinase activity stimulates breakdown of starch granules by plastidial beta-amylases. Plant Physiol. 2007, 145, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Li, X.; Fettke, J. Starch Granules in Arabidopsis thaliana Mesophyll and Guard Cells Show Similar Morphology but Differences in Size and Number. Int. J. Mol. Sci. 2021, 22, 5666. [Google Scholar] [CrossRef]
- Hejazi, M.; Mahlow, S.; Fettke, J. The glucan phosphorylation mediated by α-glucan, water dikinase (GWD) is also essential in the light phase for a functional transitory starch turn-over. Plant Signal. Behav. 2014, 9, e28892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fettke, J.; Eckermann, N.; Tiessen, A.; Geigenberger, P.; Steup, M. Identification, subcellular localization and biochemical characterization of water-soluble heteroglycans (SHG) in leaves of Arabidopsis thaliana L.: Distinct SHG reside in the cytosol and in the apoplast. Plant J. 2005, 43, 568–585. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Zhou, Y.; Fettke, J. Starch Granule Size and Morphology of Arabidopsis thaliana Starch-Related Mutants Analyzed during Diurnal Rhythm and Development. Molecules 2021, 26, 5859. https://doi.org/10.3390/molecules26195859
Liu Q, Zhou Y, Fettke J. Starch Granule Size and Morphology of Arabidopsis thaliana Starch-Related Mutants Analyzed during Diurnal Rhythm and Development. Molecules. 2021; 26(19):5859. https://doi.org/10.3390/molecules26195859
Chicago/Turabian StyleLiu, Qingting, Yuan Zhou, and Joerg Fettke. 2021. "Starch Granule Size and Morphology of Arabidopsis thaliana Starch-Related Mutants Analyzed during Diurnal Rhythm and Development" Molecules 26, no. 19: 5859. https://doi.org/10.3390/molecules26195859
APA StyleLiu, Q., Zhou, Y., & Fettke, J. (2021). Starch Granule Size and Morphology of Arabidopsis thaliana Starch-Related Mutants Analyzed during Diurnal Rhythm and Development. Molecules, 26(19), 5859. https://doi.org/10.3390/molecules26195859