
Chemical Characteristics of Croatian Traditional Istarski pršut (PDO) Produced from Two Different Pig Genotypes

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. Chemical Properties
2.2. Amino Acid Composition
2.3. Fatty Acid Composition
2.4. Lipolysis and Lipid Oxidation Indicators
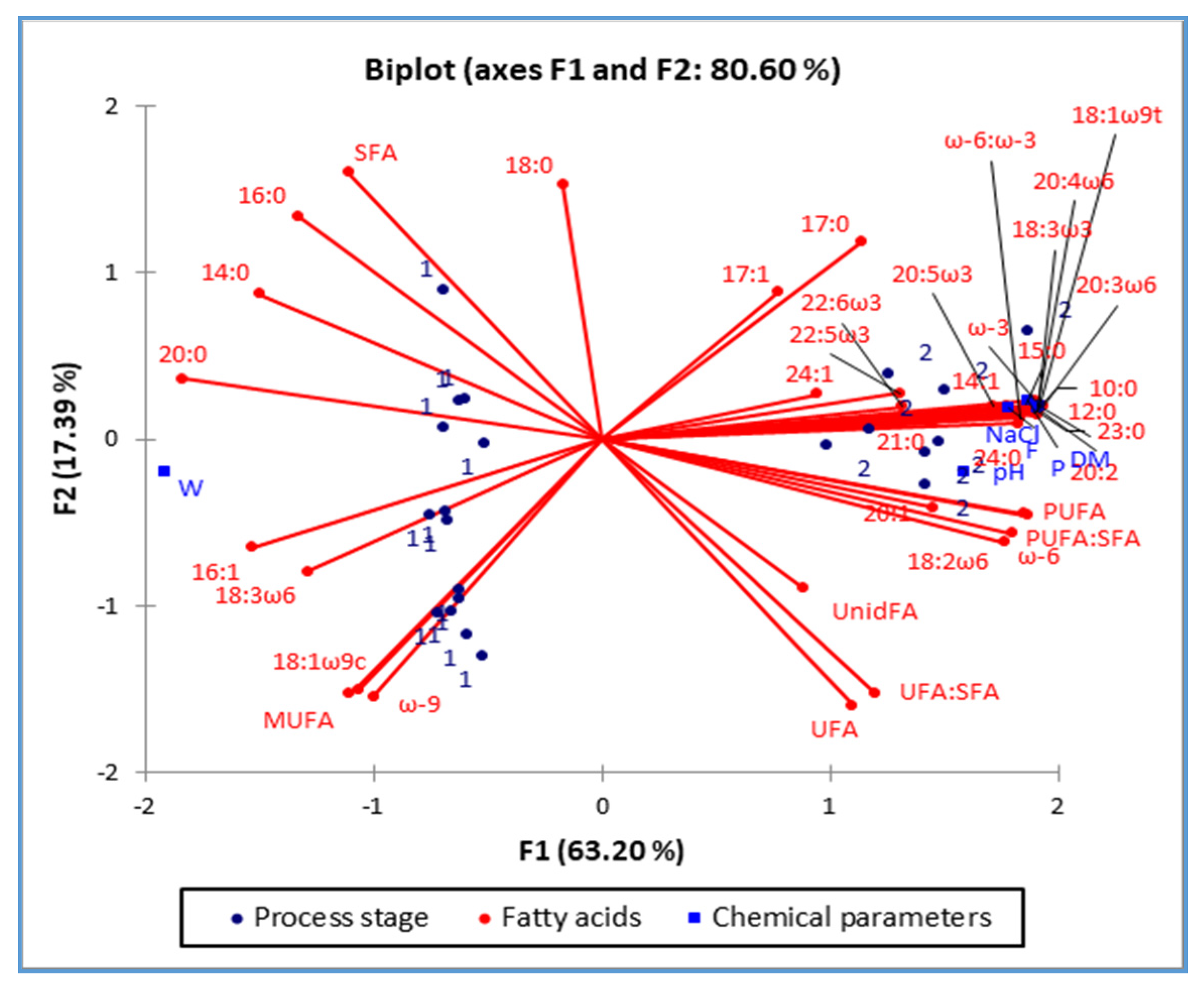
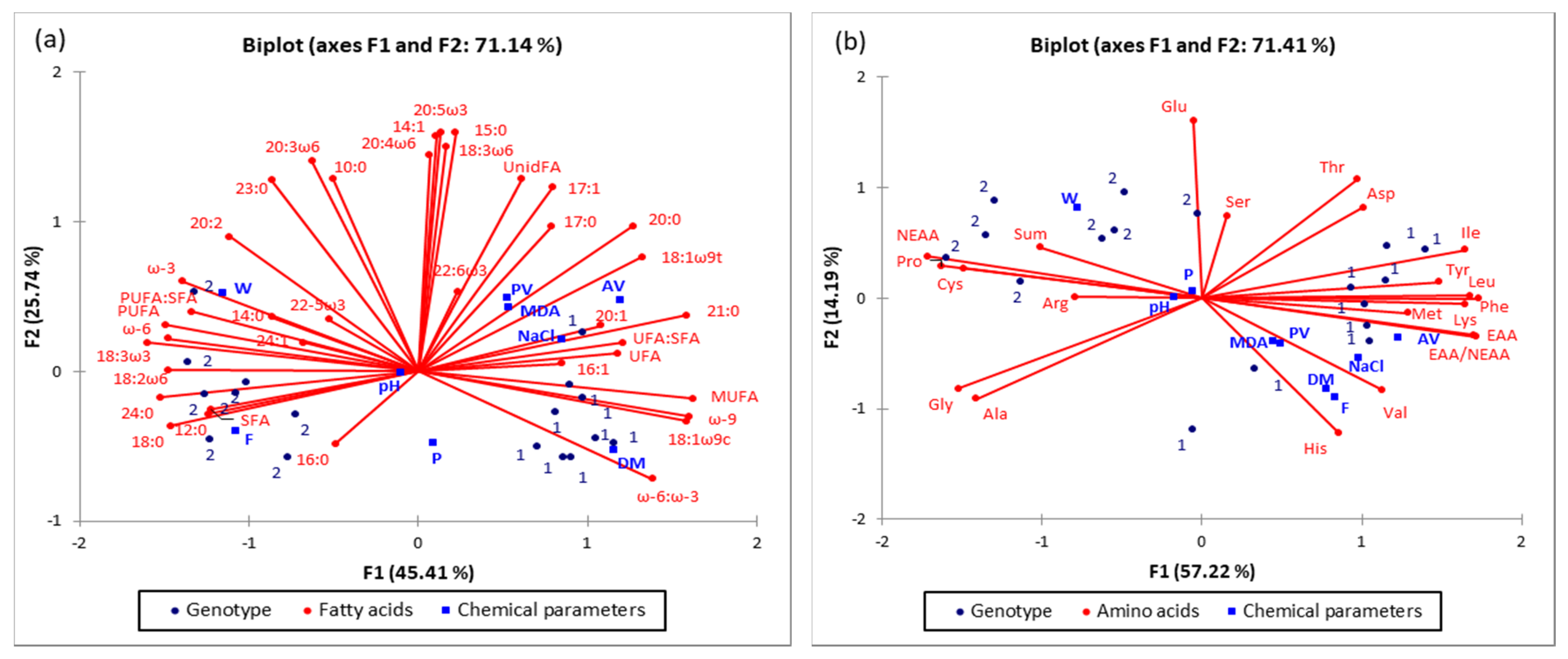
2.5. PCA Analysis
3. Materials and Methods
3.1. Raw Ham Selection and Processing
- Genotype 1 (LWxL): ♀ Large White (LW) × ♂ Swedish Landrace (L) and
- Genotype 2 (LWxL)xD: ♀ (LWxL) × ♂ Duroc (D).
3.2. Sampling
3.3. Chemical Analyses
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Petričević, S.; Marušić Radovčić, N.; Lukić, K.; Listeš, E.; Medić, H. Differentiation of dry-cured hams from different processing methods by means of volatile compounds, physico-chemical and sensory analysis. Meat Sci. 2018, 137, 217–227. [Google Scholar] [CrossRef]
- Čandek-Potokar, M.; Škrlep, M. Factor in pig production that impact the quality of dry-cured ham: A review. Animal 2012, 6, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.; Quiles, R.; Mariscal, C.; García Ruiz, A. Pig sire type and sex effects on carcass traits, meat quality and physicochemical and sensory characteristics of Serrano dry-cured ham. J. Sci. Food Agric. 2005, 85, 1914–1924. [Google Scholar] [CrossRef]
- Čandek-Potokar, M.; Zlender, B.; Lefaucheur, L.; Bonneau, M. Effects of age and/or weight at slaughter on longissimus dorsi muscle: Biochemical traits and sensory quality in pigs. Meat Sci. 1998, 48, 287–300. [Google Scholar] [CrossRef]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat–1: Composition of the lipid fraction and sensory characteristics of M. longissimus lumborum. Meat Sci. 1999, 53, 59–65. [Google Scholar] [CrossRef]
- Ruiz, J.; Ventanas, J.; Cava, R.; Andrés, A.I.; García, C. Texture and appearance of dry cured ham as affected by fat content and fatty acid composition. Food Res. Int. 2000, 33, 91–95. [Google Scholar] [CrossRef]
- Brewer, M.S.; Zhu, L.G.; McKeith, F.K. Marbling effects on quality characteristics of pork loin chops: Consumer purchase intent, visual and sensory characteristics. Meat Sci. 2001, 59, 153–163. [Google Scholar] [CrossRef]
- Fuentes, V.; Ventanas, S.; Ventanas, J.; Estevez, M. The genetic background affects composition, oxidative stability and quality traits of Iberian dry-cured hams: Purebred Iberian versus reciprocal Iberian×Duroc crossbred pigs. Meat Sci. 2014, 96, 737–743. [Google Scholar] [CrossRef]
- Bermúdez, R.; Franco, D.; Carballo, J.; Lorenzo, J.M. Sensory properties and physico-chemical changes in the biceps femoris muscle during processing of dry-cured ham from celta pigs. Effects of cross-breeding with duroc and landrace pigs. J. Food Sci. 2017, 29, 123–137. [Google Scholar] [CrossRef]
- Armero, E.; Flores, M.; Toldrá, F.; Barbosa, J.A.; Olivet, J.; Pla, M.; Baselga, M. Effect of pig sire type and sex on carcass traits, meat quality and sensory quality of dry-cured ham. J. Sci. Food Agric. 1999, 79, 1147–1154. [Google Scholar] [CrossRef]
- Fontanesi, L.; Schiavo, G.; Gallo, M.; Baiocco, C.; Galimberti, G.; Bovo, S.; Russo, V.; Buttazzoni, L. Genome-wide association study for ham weight loss at first salting in Italian Large White pigs: Towards the genetic dissection of a key trait for dry-cured ham production. Anim. Genet. 2016, 48, 103–107. [Google Scholar] [CrossRef]
- Cilla, I.; Altarriba, J.; Guerrero, L.; Gispert, M.; Martínez, L.; Moreno, C.; Beltrán, J.A.; Guàrdia, M.D.; Diestre, A.; Arnau, J.; et al. Effect of different Duroc line sires on carcass composition, meat quality and dry-cured ham acceptability. Meat Sci. 2006, 72, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Peloso, J.V.; Lopes, P.S.; Gomide, L.A.M.; Guimarães, S.E.F.; Carneiro, P.L.S. Carcass and ham quality characteristics of heavy pigs from different genetic groups intended for the production of dry-cured hams. Meat Sci. 2010, 86, 371–376. [Google Scholar] [CrossRef]
- Seong, P.N.; Park, K.M.; Kang, S.M.; Kang, G.H.; Cho, S.H.; Park, B.Y.; Ba, H.V. Effect of Particular Breed on the Chemical Composition, Texture, Color, and Sensorial Characteristics of Dry-cured Ham. Asian-Australas. J. Anim. Sci. 2014, 27, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Viterbo, V.S.; Lopez, B.I.M.; Kang, H.; Kim, H.; Song, C.W.; Seo, K.S. Genome wide association study of fatty acid composition in Duroc swine. Asian-Australas. J. Anim. Sci. 2018, 31, 1127–1133. [Google Scholar] [CrossRef]
- Bertolini, F.; Schiavo, G.; Galimberti, G.; Bovo, S.; D’Andrea, M.; Gallo, M.; Buttazzoni, L.; Rothschild, M.F.; Fontanesi, L. Genome-wide association studies for seven production traits highlight genomic regions useful to dissect dry-cured ham quality and production traits in Duroc heavy pigs. Animal 2018, 12, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Božac, R.; Kos, I.; Janječić, Z.; Kuzmanović, Ž.; Konjačić, M.; Nežak, J. Effect of different crossbreeds on chemical and sensory profiling of Istrian dry-cured ham. Acta Aliment. 2011, 40, 315–326. [Google Scholar] [CrossRef]
- Marušić Radovčić, N.; Poljanec, I.; Vidinski, P.; Novina, K.; Medic, H. Influence of different pig genotype on aroma, colour and fatty acid composition of smoked dry-cured ham. MESO Prvi Hrvat. Čas. o Mesu 2019, 6, 548–561. [Google Scholar] [CrossRef]
- Karolyi, D. Chemical properties and quality of istrian dry-cured ham. MESO Prvi Hrvat. Čas. o Mesu 2006, 4, 224–228. [Google Scholar]
- Krvavica, M.; Đugum, J. Effect of desalting on some physical characteristics of Istrian dry-cured ham. MESO Prvi Hrvat. Čas. o Mesu 2007, 1, 32–37. Available online: https://hrcak.srce.hr/index.php?show=clanak&id_clanak_jezik=34681 (accessed on 11 June 2020).
- Purriños, L.; Bermúdez, R.; Franco, D.; Carballo, J.; Lorenzo, J.M. Development of volatile compounds during the manufacture of dry-cured “Lacón” a Spanish traditional meat product. J. Food Sci. 2011, 76, 89–97. [Google Scholar] [CrossRef]
- Antequera, T.; Lopez-Bote, C.J.; Cordoba, J.J.; Garcia, C.; Ascensio, M.A.; Ventanas, J.; Garcia-Regueiro, J.A.; Diaz, I. Lipid oxidative changes in the processing of Iberian pig hams. Food Chem. 1992, 45, 105–110. [Google Scholar] [CrossRef]
- Buscailhon, S.; Gandemer, G.; Monin, G. Time-related changes in volatile compounds of lean tissue during processing of French dry-cured ham. J. Sci. Food Agric. 1994, 63, 69–75. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Technol. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Gandemer, G. Dry cured ham quality as related to lipid quality of raw material and lipid changes during processing: A review. Grasas y Aceites 1999, 60, 297–307. [Google Scholar] [CrossRef]
- Min, B.; Ahn, D.U. Mechanism of Lipid Peroxidation in Meat and Meat Products—A Review. Food Sci. Biotechnol. 2005, 14, 152–163. [Google Scholar]
- Pereira, A.L.F.; Abreu, V.K.G. Lipid peroxidation in meat and meat products. In Lipid Peroxidation; Mansour, M.A., Ed.; IntechOpen: London, UK, 2018; pp. 531–633. [Google Scholar] [CrossRef]
- Gómez, M.; Lorenzo, J.M. Effect of fat level on physicochemical, volatile compounds and sensory characteristics of dry-ripened “chorizo” from Celta pig breed. Meat Sci. 2013, 95, 658–666. [Google Scholar] [CrossRef]
- Amaral, A.B.; da Silva, M.V.; da Silva Lannes, S.C. Lipid oxidation in meat: Mechanisms and protective factors–A review. Food Sci. Technol. 2018, 38, 1–15. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A Comprehensive Review on Lipid Oxidation in Meat and Meat Products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Purrinos, L. Changes on Physico-chemical, Textural, Proteolysis, Lipolysis and Volatile Compounds During the Manufacture of Dry-cured “Lacón” from Celta Pig Breed. J. Biol. Sci. 2013, 13, 168–182. [Google Scholar] [CrossRef][Green Version]
- Salazar, E.; Abellán, A.; Cayuela, J.M.; Poto, Á.; Girón, F.; Zafrilla, P.; Tejada, L. Effect of processing time on the quality of dry-cured ham obtained from a native pig breed (Chato Murciano). Anim. Prod. Sci. 2015, 55, 113–121. [Google Scholar] [CrossRef]
- Marušić, N.; Vidaček, S.; Janči, T.; Petrak, T.; Medić, H. Determination of volatile compounds and quality parameters of traditional Istrian dry-cured ham. Meat Sci. 2014, 96, 1409–1416. [Google Scholar] [CrossRef]
- Lertpatarakomol, R.; Chaosap, C.; Chaweewan, K.; Sitthigripong, R.; Limsupavanich, R. Carcass characteristics and meat quality of purebred Pakchong 5 and crossbred pigs sired by Pakchong 5 or Duroc boar. Asian-Australas. J. Anim. Sci. 2019, 32, 585–591. [Google Scholar] [CrossRef]
- Ramírez, R.; Cava, R. Carcass composition and meat quality of three different Iberian×Duroc genotype pigs. Meat Sci. 2007, 75, 388–396. [Google Scholar] [CrossRef]
- Touma, S.; Oyadomari, M. Comparison of growth performances, carcass characteristics, and meat qualities of Okinawan indigenous Agu pigs and crossbred pigs sired by Agu or Duroc boar. Anim. Sci. J. 2020, 91, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kos, I. Physico-Chemical and Sensory Properties of Dalmatian Prosciutto of Different Pig Genotypes. Ph.D. Thesis, Faculty of Agriculture, University of Zagreb, Zagreb, Croatia, 2011. [Google Scholar]
- Sanchez, M.P.; Iannuccelli, N.; Basso, B.; Bidanel, J.; Billon, Y.; Gandemer, G.; Gilbert, H.; Larzul, C.; Legault, C.; Riquet, J.; et al. Identification of QTL with effects on intramuscular fat content fatty acid composition in a Duroc × Large White cross. BMC Genet. 2007, 8, 55. [Google Scholar] [CrossRef]
- Čandek-Potokar, M.; Monin, G.; Žlender, B. Pork quality, processing and sensory characteristics of dry-cured hams as influenced by Duroc crossing and sex. J. Anim. Sci. 2002, 80, 988–996. [Google Scholar] [CrossRef]
- Kim, J.A.; Cho, E.S.; Jeong, Y.D.; Choi, Y.H.; Kim, Y.S.; Choi, J.W.; Kim, J.S.; Jang, A.; Hong, J.K.; Sa, S.J. The effects of breed and gender on meat quality of Duroc, Pietrain, and their crossbred. J. Anim. Sci. Technol. 2020, 62, 409–419. [Google Scholar] [CrossRef]
- Ayuso, D.; González, A.; Peña, F.; Hernández-García, F.; Izquierdo, M. Effect of Fattening Period Length on Intramuscular and Subcutaneous Fatty Acid Profiles in Iberian Pigs Finished in the Montanera Sustainable System. Sustainability 2020, 12, 7937. [Google Scholar] [CrossRef]
- Eggert, J.; Grant, A.; Schinckel, A. Factors Affecting Fat Distribution in Pork Carcasses1. Prof. Anim. Sci. 2007, 23, 42–53. [Google Scholar] [CrossRef]
- Toldrá, F. Dry-Cured Meat Products; WileyBlackwell: Ames, IW, USA, 2002; pp. 27–62. [Google Scholar]
- Jiménez-Colmenero, F.; Ventanas, J.; Toldrá, F. Nutritional composition of dry-cured ham and its role in a healthy diet. Meat Sci. 2010, 84, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.-X.; Zhang, W.-G.; Xing, L.-J.; Zheng, J.-X.; Zhou, G.-H. Stability of Angiotensin I-converting Enzyme Inhibitory Activity of Peptides Extracted from Dry-cured Jinhua Ham. J. Food Nutr. Res. 2017, 5, 301–308. [Google Scholar] [CrossRef]
- Marušić, N.; Aristoy, M.-C.; Toldrá, F. Nutritional pork meat compounds as affected by ham dry-curing. Meat Sci. 2013, 93, 53–60. [Google Scholar] [CrossRef]
- Salazar, E.; Cayuela, J.M.; Abellán, A.; Bueno-Gavilá, E.; Tejada, L. Fatty Acids and Free Amino Acids Changes during Processing of a Mediterranean Native Pig Breed Dry-Cured Ham. Foods 2020, 9, 1170. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, C.S.; Schivazappa, C.; Virgili, R. Lipolysis in dry-cured ham maturation. Meat Sci. 2000, 55, 1–5. [Google Scholar] [CrossRef]
- Pateiro, M.; Franco, D.; Carril, J.A.; Lorenzo, J.M. Changes on physico-chemical properties, lipid oxidation and volatile compounds during the manufacture of celta dry-cured loin. J. Food Sci. Technol. 2014, 52, 4808–4818. [Google Scholar] [CrossRef] [PubMed]
- Larrea, V.; Perezmunuera, I.; Hernando, I.; Quiles, A.; Lluch, M. Chemical and structural changes in lipids during the ripening of Teruel dry-cured ham. Food Chem. 2007, 102, 494–503. [Google Scholar] [CrossRef]
- Cava, R.; Estévez, M.; Morcuende, D.; Antequera, T. Evolution of fatty acids from intramuscular lipid fractions during ripening of Iberian hams as affected by α-tocopheryl acetate supplementation in diet. Food Chem. 2003, 81, 199–207. [Google Scholar] [CrossRef]
- Sirtori, F.; Dimauro, C.; Bozzi, R.; Aquilani, C.; Franci, O.; Calamai, L.; Pezzati, A.; Pugliese, C. Evolution of volatile compounds and physical, chemical and sensory characteristics of Toscano PDO ham from fresh to dry-cured product. Eur. Food Res. Technol. 2019, 246, 409–424. [Google Scholar] [CrossRef]
- Andres, A.; Cava, R.; Martin, D.; Ventanas, J.; Ruiz-Carrascal, J. Lipolysis in dry-cured ham: Influence of salt content and processing conditions. Food Chem. 2005, 90, 523–533. [Google Scholar] [CrossRef]
- Storrustløkken, L.; Devle, H.M.; Håseth, T.T.; Egelandsdal, B.; Naess-Andresen, C.F.; Hollung, K.; Berg, P.; Ekeberg, D.; Alvseike, O. Lipid degradation and sensory characteristics ofM. biceps femorisin dry-cured hams from Duroc using three different processing methods. Int. J. Food Sci. Technol. 2015, 50, 522–531. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Sayago, A.; Fernández-Recamales, Á. Fatty Acid Profiling for the Authentication of Iberian Hams According to the Feeding Regime. Foods 2020, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.; Cava, R.; Ramirez, R. Effect of Physico-chemical Characteristics of Raw Muscles from Three Iberian × Duroc Genotypes on Dry-cured Meat Products Quality. Food Sci. Technol. Int. 2007, 13, 485–495. [Google Scholar] [CrossRef]
- Yim, D.-G.; Jung, J.-H.; Ali, M.; Nam, K.-C. Comparison of physicochemical traits of dry-cured ham from purebred Berkshire and crossbred Landrace × Yorkshire × Duroc (LYD) pigs. J. Anim. Sci. Technol. 2019, 61, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, S. Reducing lipid peroxidation for improving colour stability of beef and lamb: On-farm considerations. J. Sci. Food Agric. 2011, 92, 719–729. [Google Scholar] [CrossRef]
- Gandemer, G. Lipids in muscles and adipose tissues, changes during processing and sensory properties of meat products. Meat Sci. 2002, 62, 309–321. [Google Scholar] [CrossRef]
- Mariutti, L.R.; Bragagnolo, N. Influence of salt on lipid oxidation in meat and seafood products: A review. Food Res. Int. 2017, 94, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Zhou, H.-M.; Zhang, S.-L.; Pan, X.-Q.; Li, S.; Zhu, N.; Wu, Q.-R.; Wang, S.-W.; Qiao, X.-L.; Chen, W.-H. Changes of protein oxidation, lipid oxidation and lipolysis in Chinese dry sausage with different sodium chloride curing salt content. Food Sci. Hum. Wellness 2020, 9, 328–337. [Google Scholar] [CrossRef]
- Overholt, M.; Mancini, S.; Galloway, H.; Preziuso, G.; Dilger, A.; Boler, D. Effects of salt purity on lipid oxidation, sensory characteristics, and textural properties of fresh, ground pork patties. LWT 2016, 65, 890–896. [Google Scholar] [CrossRef]
- Kong, F.; Oliveira, A.; Tang, J.; Rasco, B.; Crapo, C. Salt effect on heat-induced physical and chemical changes of salmon fillet (O. gorbuscha). Food Chem. 2008, 106, 957–966. [Google Scholar] [CrossRef]
- Sakai, T.; Shimizu, Y.; Kawahara, S. Effect of NaCl on the Lipid Peroxidation-Derived Aldehyde, 4-Hydroxy-2-nonenal, Formation in Boiled Pork. Biosci. Biotechnol. Biochem. 2006, 70, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Song, O.Y.; Islam, A.; Son, J.H.; Jeong, J.Y.; Kim, H.E.; Yeon, L.S.; Khan, N.; Jamila, N.; Kim, K.S. Elemental composition of pork meat from conventional and animal welfare farms by inductively coupled plasma-optical emission spectrometry (ICP-OES) and ICP-mass spectrometry (ICP-MS) and their authentication via multivariate chemometric analysis. Meat Sci. 2021, 172, 108344. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tu, T.; Tang, X.; Zhao, S.; Qie, M.; Chen, A.; Yang, S. Authentication of organic pork and identification of geographical origins of pork in four regions of China by combined analysis of stable isotopes and multi-elements. Meat Sci. 2020, 165, 108129. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; Horwitz, W., Ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Holló, G.; Nuernberg, K.; Holló, I.; Csapó, J.; Seregi, J.; Repa, I.; Ender, K. Effect of feeding on the composition of longissimus muscle of Hungarian Grey and Holstein Friesian bulls. III. Amino acid composition and mineral content. Arch. Tierz. 2007, 50, 575–586. [Google Scholar] [CrossRef][Green Version]
- Csapó, J.; Csapó-Kiss, Z.S.; Tóth-Pósfai, I. Optimization of hydrolysis at determination of amino acid content in food and feed products. Acta Aliment. 1986, 1, 3–21. [Google Scholar]
- ISO. Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 2: Preparation of Methyl Esters of Fatty Acids; ISO 12966-2:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- ISO. Meat and Meat Products—Determination of Total Fat Content; ISO 1443:1973; International Organization for Standardization: Geneva, Switzerland, 1973. [Google Scholar]
- ISO. Animal and Vegetable Fats and Oils—Determination of Acid Value and Acidity; ISO 660:2009; International Organization for Standardization: Geneva, Switzerland, 2009. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Method 965.33, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; AOAC International: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Lemon, D.W. An Improved TBA Test for Rancidity; New Series Circular no. 51. 1975; Halifax Laboratory: Halifaks, NS, Canada, 1975. [Google Scholar]



{kind=link}
{kind=link}
| Indicators | Raw Ham | Matured Ham | p |
|---|---|---|---|
| Dry matter (DM), % | 27.62 ± 1.42 | 57.86 ± 2.26 | 0.000 |
| Water (W), % | 72.38 ± 1.42 | 42.14 ± 2.26 | 0.000 |
| Protein (P), % | 23.02 ± 0.55 | 37.70 ± 2.48 | 0.000 |
| Fat (IMF), % | 4.18 ± 1.42 | 16.12 ± 2.60 | 0.000 |
| NaCl, % | - | 8.29 ± 0.78 | - |
| pH | 5.77 ± 0.18 | 6.18 ± 0.28 | 0.000 |
| Indicators | LWxL | (LWxL)xD | p |
|---|---|---|---|
| Dry matter (DM), % | 63.00 ± 1.57 | 57.86 ± 2.26 | 0.000 |
| Water (W), % | 37.01 ± 1.57 | 42.14 ± 2.26 | 0.000 |
| Protein (P), % | 38.13 ± 3.51 | 37.70 ± 2.48 | 0.311 |
| Fat (IMF), % | 11.76 ± 2.17 | 16.12 ± 2.60 | 0.000 |
| NaCl, % | 9.04 ± 0.38 | 8.29 ± 0.78 | 0.002 |
| pH | 6.13 ± 0.18 | 6.18 ± 0.28 | 0.324 |
| Processing weight loss, % | 41.36 ± 3.48 | 40.48 ± 2.26 | 0.254 |
| Amino Acid (AA) | gAA/100 g Sample | gAA/100 g Protein | ||||
|---|---|---|---|---|---|---|
| LWxL | (LWxL)xD | p | LWxL | (LWxL)xD | p | |
| Threonine (Thr) | 1.72 ± 0.11 | 1.74 ± 0.18 | 0.392 | 4.62 ± 0.11 | 4.59 ± 0.04 | 0.226 |
| Valine (Val) | 2.02 ± 0.12 | 1.95 ± 0.21 | 0.168 | 5.40 ± 0.04 | 5.13 ± 0.19 | 0.000 |
| Methionine (Met) | 1.03 ± 0.07 | 1.02 ± 0.12 | 0.385 | 2.75 ± 0.05 | 2.68 ± 0.07 | 0.006 |
| Isoleucine (Ile) | 1.80 ± 0.13 | 1.76 ± 0.21 | 0.308 | 4.81 ± 0.12 | 4.64 ± 0.07 | 0.001 |
| Leucine (Leu) | 3.01 ± 0.20 | 2.95 ± 0.35 | 0.327 | 8.05 ± 0.11 | 7.78 ± 0.15 | 0.000 |
| Phenylalanine (Phe) | 1.55 ± 0.11 | 1.51 ± 0.18 | 0.297 | 4.15 ± 0.08 | 3.98 ± 0.07 | 0.000 |
| Histidine (His) | 1.68 ± 0.11 | 1.59 ± 0.18 | 0.083 | 4.51 ± 0.13 | 4.19 ± 0.16 | 0.000 |
| Lysine (Lys) | 3.31 ± 0.23 | 3.24 ± 0.42 | 0.323 | 8.86 ± 0.17 | 8.53 ± 0.23 | 0.001 |
| Arginine (Arg) | 2.26 ± 0.15 | 2.41 ± 0.30 | 0.089 | 6.07 ± 0.23 | 6.36 ± 0.35 | 0.021 |
| Total EAA | 18.38 ± 1.19 | 18.16 ± 2.08 | 0.386 | 49.24 ± 0.45 | 47.85 ± 0.62 | 0.000 |
| Aspartic acid (Asp) | 3.53 ± 0.24 | 3.57 ± 0.38 | 0.398 | 9.46 ± 0.09 | 9.41 ± 0.08 | 0.116 |
| Serine (Ser) | 1.49 ± 0.11 | 1.52 ± 0.19 | 0.314 | 3.97 ± 0.06 | 4.00 ± 0.13 | 0.278 |
| Glutamic acid (Glu) | 5.97 ± 0.46 | 6.25 ± 0.71 | 0.159 | 15.99 ± 0.47 | 16.45 ± 0.23 | 0.011 |
| Proline (Pro) | 1.55 ± 0.10 | 1.75 ± 0.16 | 0.003 | 4.15 ± 0.19 | 4.63 ± 0.24 | 0.000 |
| Glycine (Gly) | 1.77 ± 0.20 | 1.96 ± 0.15 | 0.021 | 4.75 ± 0.58 | 5.19 ± 0.43 | 0.047 |
| Alanine (Ala) | 2.22 ± 0.33 | 2.22 ± 0.21 | 0.494 | 5.70 ± 0.25 | 5.85 ± 0.14 | 0.073 |
| Cysteine (Cys) | 0.33 ± 0.03 | 0.41 ± 0.03 | 0.000 | 0.86 ± 0.10 | 1.09 ± 0.10 | 0.000 |
| Tyrosine (Tyr) | 1.57 ± 0.12 | 1.54 ± 0.16 | 0.293 | 4.19 ± 0.09 | 4.05 ± 0.08 | 0.001 |
| Total NEAA | 18.43 ± 1.15 | 19.20 ± 1.90 | 0.144 | 49.09 ± 0.45 | 50.66 ± 0.63 | 0.000 |
| EAA/NEAA | 1.00 ± 0.03 | 0.94 ± 0.02 | 0.000 | 1.00 ± 0.02 | 0.94 ± 0.02 | 0.000 |
| Total AA | 36.81 ± 2.29 | 37.36 ± 3.97 | 0.354 | 98.33 ± 0.14 | 98.51 ± 0.11 | 0.003 |
| NH3 | 0.61 ± 0.04 | 0.60 ± 0.06 | 0.254 | 1.65 ± 0.08 | 1.58 ± 0.05 | 0.022 |
| N% × 6.25 | 38.33 ± 3.87 | 37.60 ± 2.38 | 0.622 | - | - | - |
| Fatty Acid | Raw Ham | Matured Ham | p |
|---|---|---|---|
| C10:0 | 0.00 | 0.08 ± 0.01 | 0.000 |
| C12:0 | 0.00 | 0.08 ± 0.01 | 0.000 |
| C14:0 | 1.94 ± 0.33 | 1.40 ± 0.10 | 0.000 |
| C14:1 | 0.00 | 0.02 ± 0.01 | 0.000 |
| C15:0 | 0.00 | 0.05 ± 0.02 | 0.000 |
| C16:0 | 28.14 ± 3.19 | 24.22 ± 0.77 | 0.001 |
| C16:1 | 3.81 ± 0.54 | 2.53 ± 0.30 | 0.000 |
| C17:0 | 0.20 ± 0.06 | 0.28 ± 0.05 | 0.002 |
| C17:1 | 0.18 ± 0.05 | 0.22 ± 0.04 | 0.027 |
| C18:0 | 13.79 ± 1.72 | 13.72 ± 0.76 | 0.457 |
| C18:1ω-9t | 0.00 | 0.29 ± 0.03 | 0.000 |
| C18:1ω-9c | 44.64 ± 2.77 | 41.11 ± 0.79 | 0.001 |
| C18:2ω-6 | 5.90 ± 2.08 | 11.84 ± 1.37 | 0.000 |
| C18:3ω-3 | 0.24 ± 0.11 | 0.60 ± 0.08 | 0.000 |
| C18:3ω-6 | 0.00 | 0.05 ± 0.02 | 0.000 |
| C20:0 | 0.20 ± 0.05 | 0.00 | 0.000 |
| C20:1 | 0.54 ± 0.16 | 0.88 ± 0.14 | 0.000 |
| C20:2 | 0.00 | 0.62 ± 0.08 | 0.000 |
| C20:3ω-6 | 0.00 | 0.18 ± 0.02 | 0.000 |
| C20:4ω-6 | 0.00 | 0.72 ± 0.18 | 0.000 |
| C20:5ω-3 | 0.00 | 0.02 ± 0.01 | 0.000 |
| C21:0 | 0.00 | 0.02 ± 0.01 | 0.000 |
| C22:5ω-3 | 0.00 | 0.20 ± 0.24 | 0.001 |
| C22:6ω-3 | 0.00 | 0.02 ± 0.02 | 0.000 |
| C23:0 | 0.00 | 0.11 ± 0.01 | 0.000 |
| C24:0 | 0.00 | 0.03 ± 0.01 | 0.000 |
| C24:1 | 0.00 | 0.09 ± 0.18 | 0.029 |
| UnidFA | 0.39 ± 0.34 | 0.64 ± 0.12 | 0.029 |
| SFA | 44.27 ± 4.73 | 39.98 ± 0.98 | 0.010 |
| MUFA | 49.17 ± 2.97 | 45.13 ± 0.99 | 0.000 |
| PUFA | 6.14 ± 2.17 | 14.25 ± 1.57 | 0.000 |
| UFA | 55.30 ± 4.56 | 59.39 ± 1.01 | 0.011 |
| PUFA/SFA | 0.14 ± 0.05 | 0.36 ± 0.05 | 0.000 |
| UFA/SFA | 1.27 ± 0.21 | 1.49 ± 0.06 | 0.004 |
| ω-9 | 44.64 ± 2.77 | 41.40 ± 0.80 | 0.002 |
| ω-6 | 5.90 ± 0.52 | 12.79 ± 1.39 | 0.000 |
| ω-3 | 0.24 ± 0.11 | 0.84 ± 0.26 | 0.000 |
| ω-6/ω-3 | 23.54 ± 2.39 | 15.98 ± 3.05 | 0.001 |
| Fatty Acid | LWxL | (LWxL)xD | p |
|---|---|---|---|
| C10:0 | 0.07 ± 0.02 | 0.08 ± 0.01 | 0.124 |
| C12:0 | 0.07 ± 0.01 | 0.08 ± 0.01 | 0.000 |
| C14:0 | 1.29 ± 0.13 | 1.40 ± 0.10 | 0.032 |
| C14:1 | 0.03 ± 0.06 | 0.02 ± 0.01 | 0.249 |
| C15:0 | 0.09 ± 0.11 | 0.05 ± 0.02 | 0.173 |
| C16:0 | 23.55 ± 1.16 | 24.22 ± 0.77 | 0.086 |
| C16:1 | 2.84 ± 0.27 | 2.53 ± 0.30 | 0.014 |
| C17:0 | 0.40 ± 0.13 | 0.28 ± 0.05 | 0.009 |
| C17:1 | 0.37 ± 0.15 | 0.22 ± 0.04 | 0.008 |
| C18:0 | 11.23 ± 0.47 | 13.72 ± 0.76 | 0.000 |
| C18:1ω-9t | 0.46 ± 0.07 | 0.29 ± 0.03 | 0.000 |
| C18:1ω-9c | 46.42 ± 1.55 | 41.11 ± 0.79 | 0.000 |
| C18:2ω-6 | 8.48 ± 1.24 | 11.84 ± 1.37 | 0.000 |
| C18:3ω-3 | 0.17 ± 0.08 | 0.60 ± 0.08 | 0.000 |
| C18:3ω-6 | 0.06 ± 0.06 | 0.05 ± 0.02 | 0.263 |
| C20:0 | 0.23 ± 0.10 | 0.00 | 0.000 |
| C20:1 | 1.04 ± 0.09 | 0.88 ± 0.14 | 0.003 |
| C20:2 | 0.48 ± 0.12 | 0.62 ± 0.08 | 0.006 |
| C20:3ω-6 | 0.16 ± 0.06 | 0.18 ± 0.02 | 0.134 |
| C20:4ω-6 | 0.82 ± 0.25 | 0.72 ± 0.18 | 0.176 |
| C20:5ω-3 | 0.04 ± 0.06 | 0.02 ± 0.01 | 0.231 |
| C21:0 | 0.39 ± 0.07 | 0.02 ± 0.01 | 0.000 |
| C22:5ω-3 | 0.12 ± 0.04 | 0.20 ± 0.24 | 0.145 |
| C22:6ω-3 | 0.03 ± 0.01 | 0.02 ± 0.02 | 0.169 |
| C23:0 | 0.08 ± 0.05 | 0.11 ± 0.01 | 0.033 |
| C24:0 | 0.00 | 0.03 ± 0.01 | 0.000 |
| C24:1 | 0.00 | 0.09 ± 0.18 | 0.000 |
| UnidFA | 0.89 ± 0.31 | 0.64 ± 0.12 | 0.022 |
| SFA | 37.39 ± 1.27 | 39.98 ± 0.98 | 0.000 |
| MUFA | 51.16 ± 1.29 | 45.13 ± 0.99 | 0.000 |
| PUFA | 10.35 ± 1.53 | 14.25 ± 1.57 | 0.000 |
| UFA | 61.51 ± 1.00 | 59.39 ± 1.01 | 0.000 |
| PUFA/SFA | 0.28 ± 0.05 | 0.36 ± 0.05 | 0.001 |
| UFA/SFA | 1.65 ± 0.08 | 1.49 ± 0.06 | 0.000 |
| ω-9 | 46.87 ± 1.51 | 41.40 ± 0.80 | 0.000 |
| ω-6 | 9.52 ± 1.38 | 12.79 ± 1.39 | 0.000 |
| ω-3 | 0.36 ± 0.16 | 0.84 ± 0.26 | 0.000 |
| ω-6/ω-3 | 29.24 ± 8.73 | 15.98 ± 3.05 | 0.000 |
| Indicators | LWxL | (LWxL)xD | p |
|---|---|---|---|
| Acid value (AV), KOH/g fat | 38.50 ± 8.46 | 22.20 ± 5.16 | 0.000 |
| Peroxide value (PV), meq/kg fat | 17.80 ± 8.09 | 11.10 ± 7.45 | 0.035 |
| TBARS, mg MDA/kg sample | 0.42 ± 0.19 | 0.28 ± 0.12 | 0.037 |
| Variables | IMF | AV | PV | TBARS |
|---|---|---|---|---|
| C10:0 | 0.03 | −0.11 | 0.06 | 0.10 |
| C12:0 | 0.38 | −0.67 ** | −0.34 | −0.34 |
| C14:0 | 0.20 | −0.39 | −0.23 | −0.06 |
| C14:1 | 0.24 | 0.26 | 0.36 | 0.36 |
| C15:0 | 0.30 | 0.34 | 0.40 | 0.40 |
| C16:0 | 0.08 | −0.29 | −0.24 | −0.12 |
| C16:1 | −0.40 | 0.25 | 0.36 | 0.46 * |
| C17:0 | −0.57 | 0.46 * | 0.32 | 0.32 |
| C17:1 | −0.43 | 0.54 * | 0.44 * | 0.43 |
| C18:0 | 0.61 ** | −0.77 *** | −0.44 * | −0.40 |
| C18:1ω-9t | −0.70 *** | 0.77 *** | 0.38 | 0.41 |
| C18:1ω-9c | −0.42 | 0.70 *** | 0.37 | 0.32 |
| C18:2ω-6 | 0.51 * | −0.65 ** | −0.39 | −0.42 |
| C18:3ω-3 | 0.60 ** | −0.71 *** | −0.37 | −0.34 |
| C18:3ω-6 | −0.21 | 0.24 | 0.27 | 0.27 |
| C20:0 | −0.52 * | 0.78 *** | 0.50 * | 0.43 |
| C20:1 | −0.28 | 0.70 *** | 0.18 | 0.11 |
| C20:2 | 0.33 | −0.28 | −0.17 | −0.23 |
| C20:3ω-6 | −0.24 | −0.08 | 0.07 | 0.08 |
| C20:4ω-6 | −0.60 ** | 0.31 | 0.27 | 0.30 |
| C20:5ω-3 | −0.28 | 0.28 | 0.35 | 0.32 |
| C21:0 | −0.61 ** | 0.81 *** | 0.46 * | 0.40 |
| C22:5ω-3 | −0.28 | −0.16 | −0.05 | 0.01 |
| C22:6ω-3 | −0.34 | 0.23 | 0.29 | 0.28 |
| C23:0 | −0.20 | −0.17 | 0.02 | −0.01 |
| C24:0 | 0.66 ** | −0.69 ** | −0.40 | −0.49 * |
| C24:1 | −0.15 | −0.30 | −0.12 | −0.05 |
| UnidFA | −0.62 ** | 0.50 * | 0.23 | 0.21 |
| SFA | 0.41 | −0.63 ** | −0.39 | −0.28 |
| MUFA | −0.48 * | 0.73 *** | 0.41 | 0.37 |
| PUFA | 0.42 | −0.60 ** | −0.34 | −0.35 |
| UFA | −0.36 | 0.65 ** | 0.36 | 0.24 |
| PUFA/SFA | 0.35 | −0.51 * | −0.28 | −0.32 |
| UFA/SFA | −0.39 | 0.62 ** | 0.37 | 0.25 |
| ω-6 | 0.43 | −0.61 ** | −0.36 | −0.38 |
| ω-3 | 0.23 | −0.54 * | −0.22 | −0.17 |
| ω-6/ω-3 | −0.29 | 0.53 * | 0.15 | 0.09 |
| ω-9 | −0.44 * | 0.71 *** | 0.37 | 0.32 |
| IMF | 1.00 | −0.48* | −0.18 | −0.29 |
| AV | −0.48 * | 1.00 | 0.34 | 0.27 |
| PV | −0.18 | 0.34 | 1.00 | 0.95 *** |
| TBARS | −0.29 | 0.27 | 0.95 *** | 1.00 |
| NaCl | −0.56 * | 0.16 | 0.14 | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krvavica, M.; Lasić, D.; Kljusurić, J.G.; Đugum, J.; Janović, Š.; Milovac, S.; Bošnir, J. Chemical Characteristics of Croatian Traditional Istarski pršut (PDO) Produced from Two Different Pig Genotypes. Molecules 2021, 26, 4140. https://doi.org/10.3390/molecules26144140
Krvavica M, Lasić D, Kljusurić JG, Đugum J, Janović Š, Milovac S, Bošnir J. Chemical Characteristics of Croatian Traditional Istarski pršut (PDO) Produced from Two Different Pig Genotypes. Molecules. 2021; 26(14):4140. https://doi.org/10.3390/molecules26144140
Chicago/Turabian StyleKrvavica, Marina, Dario Lasić, Jasenka Gajdoš Kljusurić, Jelena Đugum, Špiro Janović, Srđan Milovac, and Jasna Bošnir. 2021. "Chemical Characteristics of Croatian Traditional Istarski pršut (PDO) Produced from Two Different Pig Genotypes" Molecules 26, no. 14: 4140. https://doi.org/10.3390/molecules26144140
APA StyleKrvavica, M., Lasić, D., Kljusurić, J. G., Đugum, J., Janović, Š., Milovac, S., & Bošnir, J. (2021). Chemical Characteristics of Croatian Traditional Istarski pršut (PDO) Produced from Two Different Pig Genotypes. Molecules, 26(14), 4140. https://doi.org/10.3390/molecules26144140