
Stoichiometric Analysis of Shifting in Subcellular Compartmentalization of HSP70 within Ischemic Penumbra

 ,
,  , ,
, , 

Abstract
1. Introduction
2. Results
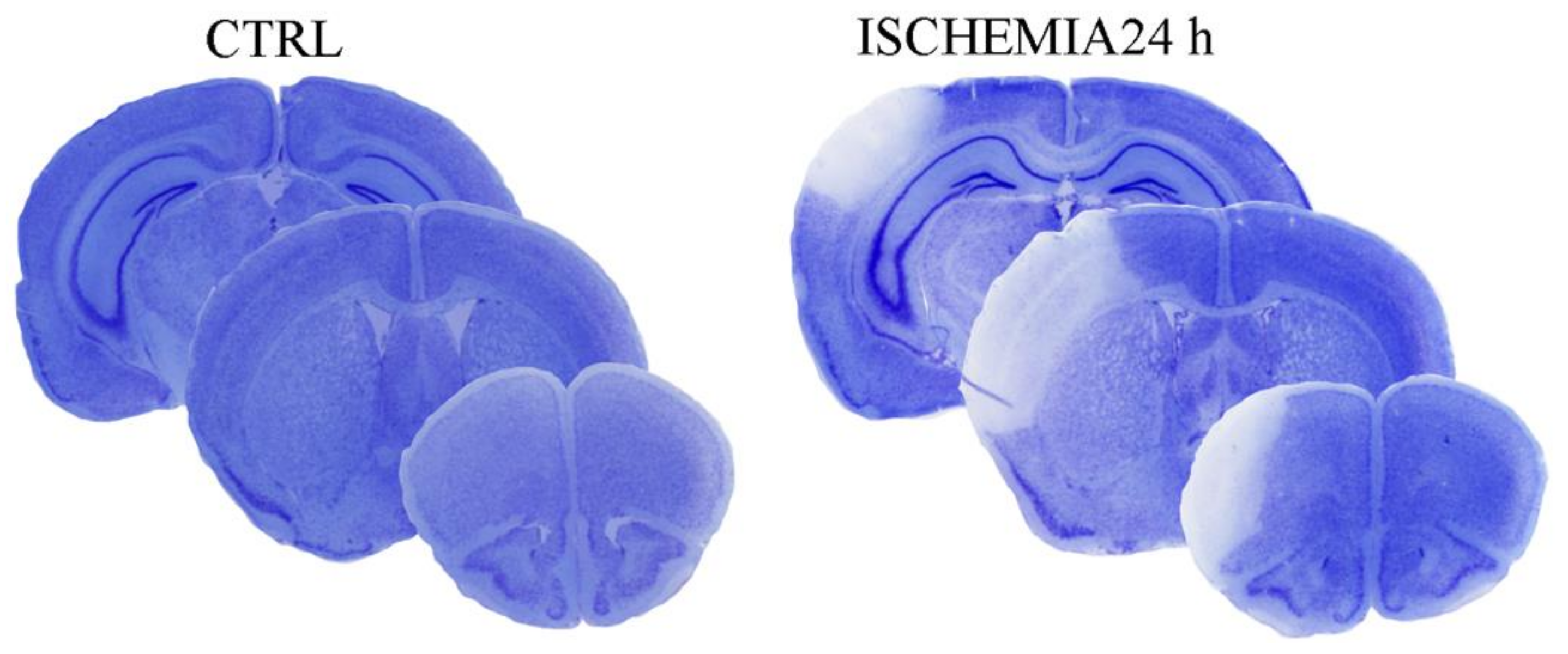
2.1. The Outcome of Permanent Ischemia
2.2. The Selection of the Peri-Infarct Regions
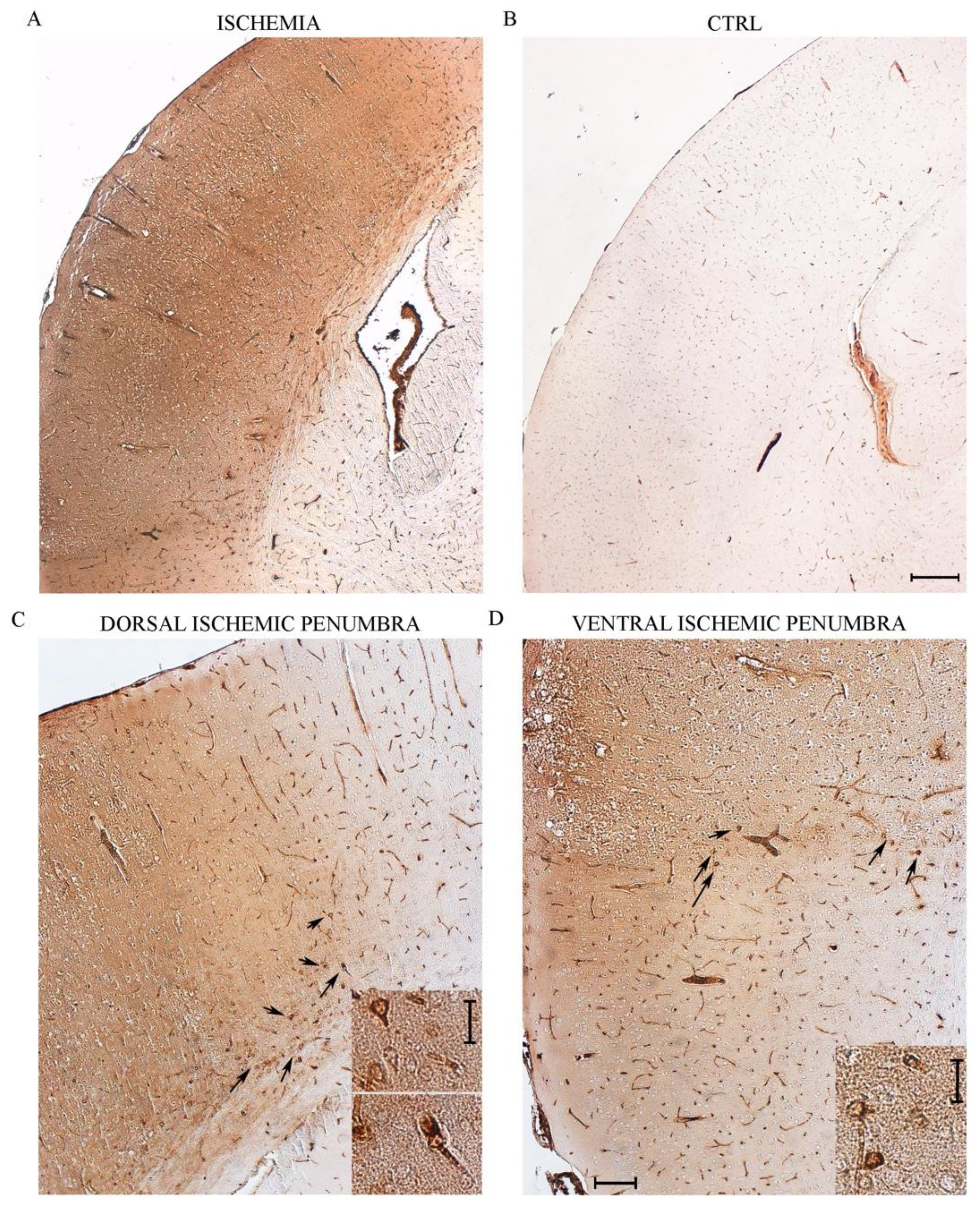
2.3. Immunohistochemistry at Light Microscopy for HSP70
2.4. From Light to Electron Microscopy through Identifying the Same Neurons Evident at Light Microscopy Semi-Thin Slices at TEM Electron Density
2.5. Ultrastructural Vacuolar Compartmentalization of HSP70
2.6. Ultrastructural of Mitochondrial Compartmentalization of HSP70
2.7. Number and Diameter of Mitochondria, Cytosolic Vacuoles and Cells within Area Penumbra
3. Discussion
4. Materials and Methods
4.1. Permanent Focal Ischemia in Mice
4.2. Histology
4.3. Immunohistochemistry
4.4. HSP70 Detection
4.5. Transmission Electron Microscopy (TEM)
4.6. Post-Embedding Immuno-Electron Microscopy
4.7. Ultrastructural Morphometry
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Weinstein, P.R.; Hong, S.; Sharp, F.R. Molecular identification of the ischemic penumbra. Stroke 2004, 35, 2666–26670. [Google Scholar] [CrossRef]
- Hata, R.; Maeda, K.; Hermann, D.; Mies, G.; Hossmann, K.A. Dynamics of regional brain metabolism and gene expression after middle cerebral artery occlusion in mice. J. Cereb. Blood Flow Metab. 2000, 20, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, H.; Sharp, F.R.; Hill, M.P.; Koistinaho, J.; Sagar, S.M.; Chan, P.H. Induction of 70-kDa heat shock protein and hsp70 mRNA following transient focal cerebral ischemia in the rat. J. Cereb. Blood Flow Metab. 1993, 13, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Han, Y.; Lee, J.E.; Yenari, M.A. The 70-kDa heat shock protein (Hsp70) as a therapeutic target for stroke. Expert Opin. Ther. Targets 2018, 22, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Rajdev, S.; Hara, K.; Kokubo, Y.; Mestril, R.; Dillmann, W.; Weinstein, P.R.; Sharp, F.R. Mice overexpressing rat heat shock protein 70 are protected against cerebral infarction. Ann. Neurol. 2000, 47, 782–791. [Google Scholar] [CrossRef]
- Sun, Y.; Ouyang, Y.B.; Xu, L.; Chow, A.M.; Anderson, R.; Hecker, J.G.; Giffard, R.G. The carboxyl-terminal domain of inducible Hsp70 protects from ischemic injury in vivo and in vitro. J. Cereb. Blood Flow Metab. 2006, 26, 937–950. [Google Scholar] [CrossRef]
- Kudo, T.; Kanemoto, S.; Hara, H.; Morimoto, N.; Morihara, T.; Kimura, R.; Tabira, T.; Imaizumi, K.; Takeda, M. A molecular chaperone inducer protects neurons from ER stress. Cell Death Differ. 2008, 15, 364–375. [Google Scholar] [CrossRef]
- Mohanan, A.; Deshpande, S.; Jamadarkhana, P.G.; Kumar, P.; Gupta, R.C.; Chauthaiwale, V.; Dutt, C. Delayed intervention in experimental stroke with TRC051384--a small molecule HSP70 inducer. Neuropharmacology 2011, 60, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Demyanenko, S.; Nikul, V.; Rodkin, S.; Davletshin, A.; Evgen’ev, M.B.; Garbuz, D.G. Exogenous recombinant Hsp70 mediates neuroprotection after photothrombotic stroke. Cell Stress Chaperones 2021, 26, 103–114. [Google Scholar] [CrossRef]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1997; pp. 1–190. [Google Scholar]
- Sanz, O.; Estrada, A.; Ferrer, I.; Planas, A. Differential cellular distribution and dynamics of HSP70, cyclooxygenase-2 and c-Fos in the rat brain after transient focal ischemia or kainic acid. Neuroscience 1997, 80, 221–232. [Google Scholar] [CrossRef]
- Kokubo, Y.; Matson, G.B.; Liu, J.; Mancuso, A.; Kayama, T.; Sharp, F.R.; Weinstein, P.R. Correlation between changes in apparent diffusion coefficient and induction of heat shock protein, cell-specific injury marker expression and protein synthesis reduction on diffusion-weighted magnetic resonance images after temporary focal cerebral ischemia in rats. J. Neurosurg. 2002, 96, 1084–1093. [Google Scholar]
- Brea, D.; Agulla, J.; Staes, A.; Gevaert, K.; Campos, F.; Sobrino, T.; Blanco, M.; Dávalos, A.; Castillo, J.; Ramos-Cabrer, P. Study of Protein Expression in Peri-Infarct Tissue after Cerebral Ischemia. Sci. Rep. 2015, 5, 12030. [Google Scholar] [CrossRef]
- Scott, M.A.; Locke, M.; Buck, L.T. Tissue-specific expression of inducible and constitutive Hsp70 isoforms in the western painted turtle. J. Exp. Biol. 2003, 206, 303–311. [Google Scholar] [CrossRef]
- Kinouchi, H.; Sharp, F.R.; Koistinaho, J.; Hicks, K.; Kamii, H.; Chan, P.H. Induction of heat shock hsp70 mRNA and HSP70 kDa protein in neurons in the ‘penumbra’ following focal cerebral ischemia in the rat. Brain Res. 1993, 619, 334–338. [Google Scholar] [CrossRef]
- Sharp, F.R.; Lu, A.; Tang, Y.; Millhorn, D.E. Multiple molecular penumbras after focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2000, 20, 1011–1032. [Google Scholar] [CrossRef]
- Dietrich, H.H.; Dacey, R.G., Jr. Molecular keys to the problems of cerebral vasospasm. Neurosurgery 2000, 46, 517–530. [Google Scholar] [CrossRef]
- Ouyang, Y.B.; Lu, Y.; Yue, S.; Xu, L.J.; Xiong, X.X.; White, R.E.; Sun, X.; Giffard, R.G. miR-181 regulates GRP78 and influences outcome from cerebral ischemia in vitro and in vivo. Neurobiol. Dis. 2012, 45, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Sharp, F.R.; Zhan, X.; Liu, D.Z. Heat shock proteins in the brain: Role of Hsp70, Hsp27, and HO-1 (Hsp32) and their therapeutic potential. Transl. Stroke Res. 2013, 4, 685–692. [Google Scholar] [CrossRef]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Vishwanathan, V.; Birje, A.; Sinha, D.; D’Silva, P. Evolving paradigms on the interplay of mitochondrial Hsp70 chaperone system in cell survival and senescence. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Mivechi, N.F.; Moskophidis, D. Insights into regulation and function of the major stress-induced hsp70 molecular chaperone in vivo: Analysis of mice with targeted gene disruption of the hsp70.1 or hsp70.3 gene. Mol. Cell Biol. 2001, 21, 8575–8591. [Google Scholar] [CrossRef]
- Lee, J.S.; Seo, J.S. Differential expression of two stress-inducible hsp70 genes by various stressors. Exp. Mol. Med. 2002, 34, 131–136. [Google Scholar] [CrossRef][Green Version]
- Jakobsson, M.E.; Moen, A.; Bousset, L.; Egge-Jacobsen, W.; Kernstock, S.; Melki, R.; Falnes, P.Ø. Identification and characterization of a novel human methyltransferase modulating Hsp70 protein function through lysine methylation. J. Biol. Chem. 2013, 288, 27752–27763. [Google Scholar] [CrossRef]
- Mambula, S.S.; Calderwood, S.K. Heat shock protein 70 is secreted from tumor cells by a nonclassical pathway involving lysosomal endosomes. J. Immunol. 2006, 177, 7849–7857. [Google Scholar] [CrossRef]
- Quintana-Gallardo, L.; Martín-Benito, J.; Marcilla, M.; Espadas, G.; Sabidó, E.; Valpuesta, J.M. The cochaperone CHIP marks Hsp70- and Hsp90-bound substrates for degradation through a very flexible mechanism. Sci. Rep. 2019, 9, 5102. [Google Scholar] [CrossRef]
- Backhauss, C.; Karkoutly, C.; Welsch, M.; Krieglstein, J. A mouse model of focal cerebral ischemia for screening neuroprotective drug effects. J. Pharmacol. Toxicol. Methods 1992, 27, 27–32. [Google Scholar] [CrossRef]
- Mastroiacovo, F.; Busceti, C.L.; Biagioni, F.; Moyanova, S.G.; Meisler, M.H.; Battaglia, G.; Caricasole, A.; Bruno, V.; Nicoletti, F. Induction of the Wnt antagonist, Dickkopf-1, contributes to the development of neuronal death in models of brain focal ischemia. J. Cereb. Blood Flow Metab. 2009, 29, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Deguchi, K.; Yamashita, T.; Ohta, Y.; Shang, J.; Tian, F.; Liu, N.; Panin, V.L.; Ikeda, Y.; Matsuura, T.; et al. Temporal and spatial differences of multiple protein expression in the ischemic penumbra after transient MCAO in rats. Brain Res. 2010, 1343, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, G.; Lenzi, P.; Busceti, C.L.; Ferrucci, M.; Falleni, A.; Bruno, V.; Paparelli, A.; Fornai, F. Mechanisms involved in the formation of dopamine-induced intracellular bodies within striatal neurons. J. Neurochem. 2007, 101, 1414–1427. [Google Scholar] [CrossRef] [PubMed]
- Bendayan, M.; Zollinger, M. Ultrastructural localization of antigenic sites on osmium-fixed tissues applying the protein A-gold technique. J. Histochem Cytochem. 1983, 31, 101–109. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, D.; Mattii, L.; Moscato, S.; Bernardini, N.; Segnani, C.; Dolfi, A.; Bianchi, F. Immunohistochemical demonstration of the small GTPase RhoA on epoxy- resin embedded sections. Micron 2004, 35, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, P.; Lazzeri, G.; Biagioni, F.; Busceti, C.L.; Gambardella, S.; Salvetti, A.; Fornai, F. The Autophagoproteasome a Novel Cell Clearing Organelle in Baseline and Stimulated Conditions. Front. Neuroanat. 2016, 10, 78. [Google Scholar] [CrossRef]
- Stirling, J.W. Use of tannic acid and silver enhancer to improve staining for electron microscopy and immunogold labeling. J. Histochem. Cytochem. 1993, 41, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Natale, G.; Lenzi, P.; Lazzeri, G.; Falleni, A.; Biagioni, F.; Ryskalin, L.; Fornai, F. Compartment-dependent mitochondrial alterations in experimental ALS, the effects of mitophagy and mitochondriogenesis. Front. Cell. Neurosci. 2015, 9, 434. [Google Scholar] [CrossRef]
- Lucocq, J.M.; Habermann, A.; Watt, S.; Backer, J.M.; Mayhew, T.M.; Griffiths, G. A rapid method for assessing the distribution of gold labeling on thin sections. J. Histochem. Cytochem. 2004, 52, 991–1000. [Google Scholar] [CrossRef]
- Fornai, F.; Lenzi, P.; Lazzeri, G.; Ferrucci, M.; Fulceri, F.; Giorgi, F.S.; Falleni, A.; Ruggieri, S.; Paparelli, A. Fine ultrastructure and biochemistry of PC12 cells: A comparative approach to understand neurotoxicity. Brain Res. 2007, 1129, 174–190. [Google Scholar] [CrossRef]
- Lenzi, P.; Marongiu, R.; Falleni, A.; Gelmetti, V.; Busceti, C.L.; Michiorri, S.; Valente, E.M.; Fornai, F. A subcellular analysis of genetic modulation of PINK1 on mitochondrial alterations, autophagy and cell death. Arch. Ital. Biol. 2012, 150, 194–217. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Dors/Ventr 1 | Ischemia Dors/Ventr 1 | |
|---|---|---|
| Number of mitochondria/cell | 20.77 ± 1.49/22.52 ± 1.37 | 19.82 ± 1.82/19.7 ± 0.62 |
| Number of vacuoles/cell | 6.97 ± 0.19/6.9 ± 0.39 | 6.58 ± 0.8/6.05 ± 0.87 |
| Mean of mitochondria area | 0.25 ± 0.01/0.24 ± 0.002 | 0.35 ± 0.003/0.36 ± 0.008 |
| Mean of vacuoles area | 0.70 ± 0.02/0.67 ± 0.02 | 2.06 ± 0.10/2.021 ± 0.06 |
| Total mitochondrial area/cell | 5.19 ± 0.3/5.46 ± 0.36 | 6.71 ± 0.10/7.57 ± 0.06 |
| Total vacuolar area/cell | 4.79 ± 0.36/4.79 ± 0.38 | 12.93 ± 2.08/14.59 ± 1.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastroiacovo, F.; Biagioni, F.; Lenzi, P.; Ryskalin, L.; Puglisi-Allegra, S.; Nicoletti, F.; Frati, A.; Fornai, F. Stoichiometric Analysis of Shifting in Subcellular Compartmentalization of HSP70 within Ischemic Penumbra. Molecules 2021, 26, 3578. https://doi.org/10.3390/molecules26123578
Mastroiacovo F, Biagioni F, Lenzi P, Ryskalin L, Puglisi-Allegra S, Nicoletti F, Frati A, Fornai F. Stoichiometric Analysis of Shifting in Subcellular Compartmentalization of HSP70 within Ischemic Penumbra. Molecules. 2021; 26(12):3578. https://doi.org/10.3390/molecules26123578
Chicago/Turabian StyleMastroiacovo, Federica, Francesca Biagioni, Paola Lenzi, Larisa Ryskalin, Stefano Puglisi-Allegra, Ferdinando Nicoletti, Alessandro Frati, and Francesco Fornai. 2021. "Stoichiometric Analysis of Shifting in Subcellular Compartmentalization of HSP70 within Ischemic Penumbra" Molecules 26, no. 12: 3578. https://doi.org/10.3390/molecules26123578
APA StyleMastroiacovo, F., Biagioni, F., Lenzi, P., Ryskalin, L., Puglisi-Allegra, S., Nicoletti, F., Frati, A., & Fornai, F. (2021). Stoichiometric Analysis of Shifting in Subcellular Compartmentalization of HSP70 within Ischemic Penumbra. Molecules, 26(12), 3578. https://doi.org/10.3390/molecules26123578