
Potential Benefits of Flavonoids on the Progression of Atherosclerosis by Their Effect on Vascular Smooth Muscle Excitability


Abstract
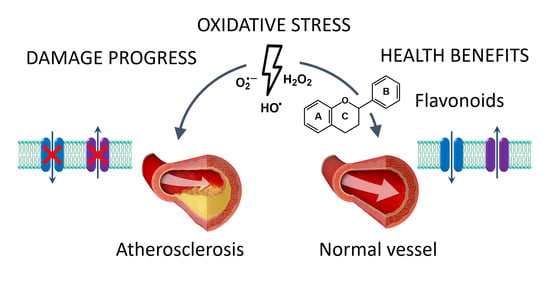
1. Introduction
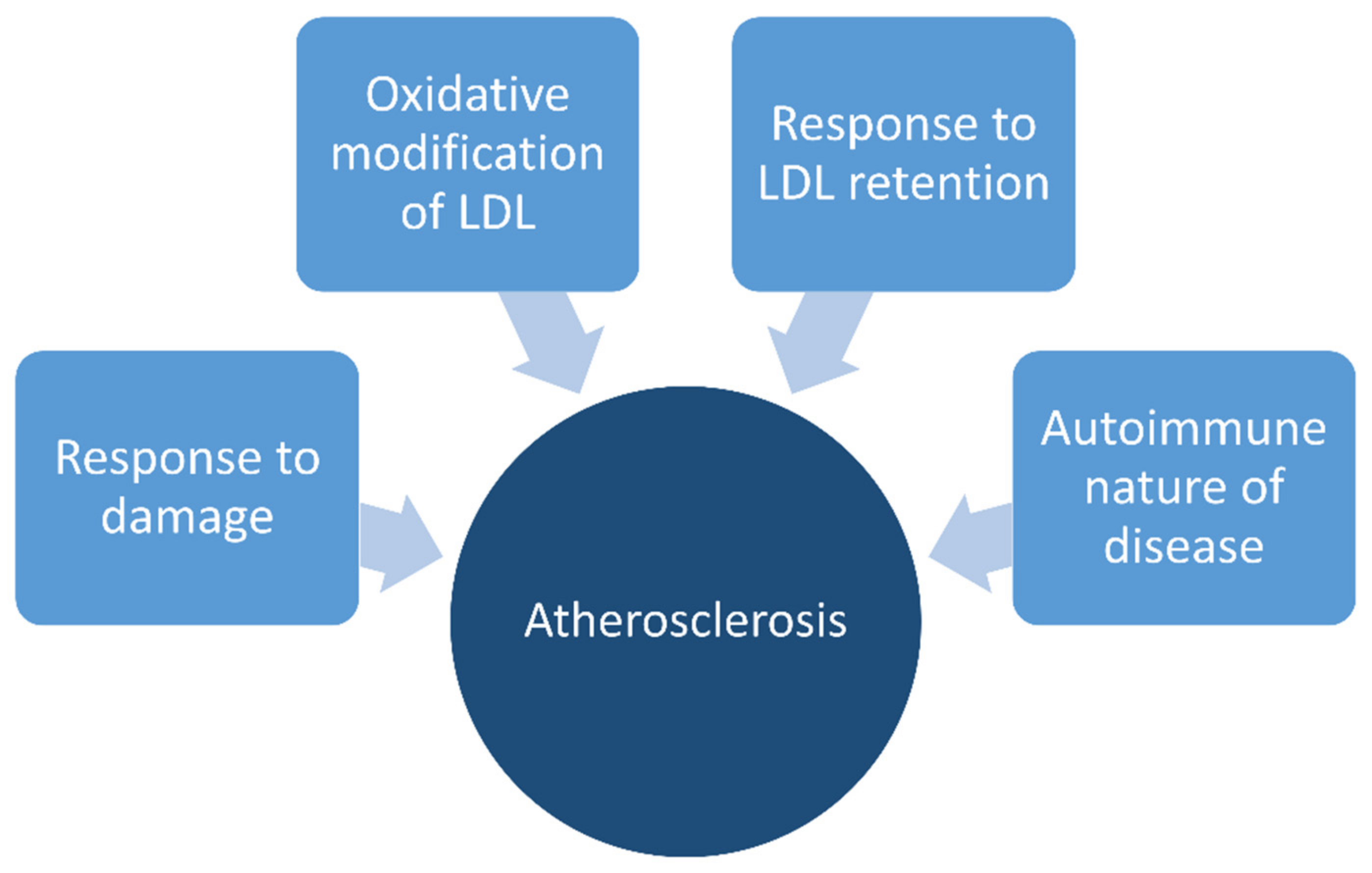
2. Atherosclerosis
2.1. General Concepts
2.2. Stages of Atherosclerosis
2.3. Role of Oxidation
2.4. Damage in Vascular Smooth Muscle Cells
2.5. Natural Compounds for the Treatment of Atherosclerosis
3. Flavonoids in Atherosclerosis
3.1. General Concepts
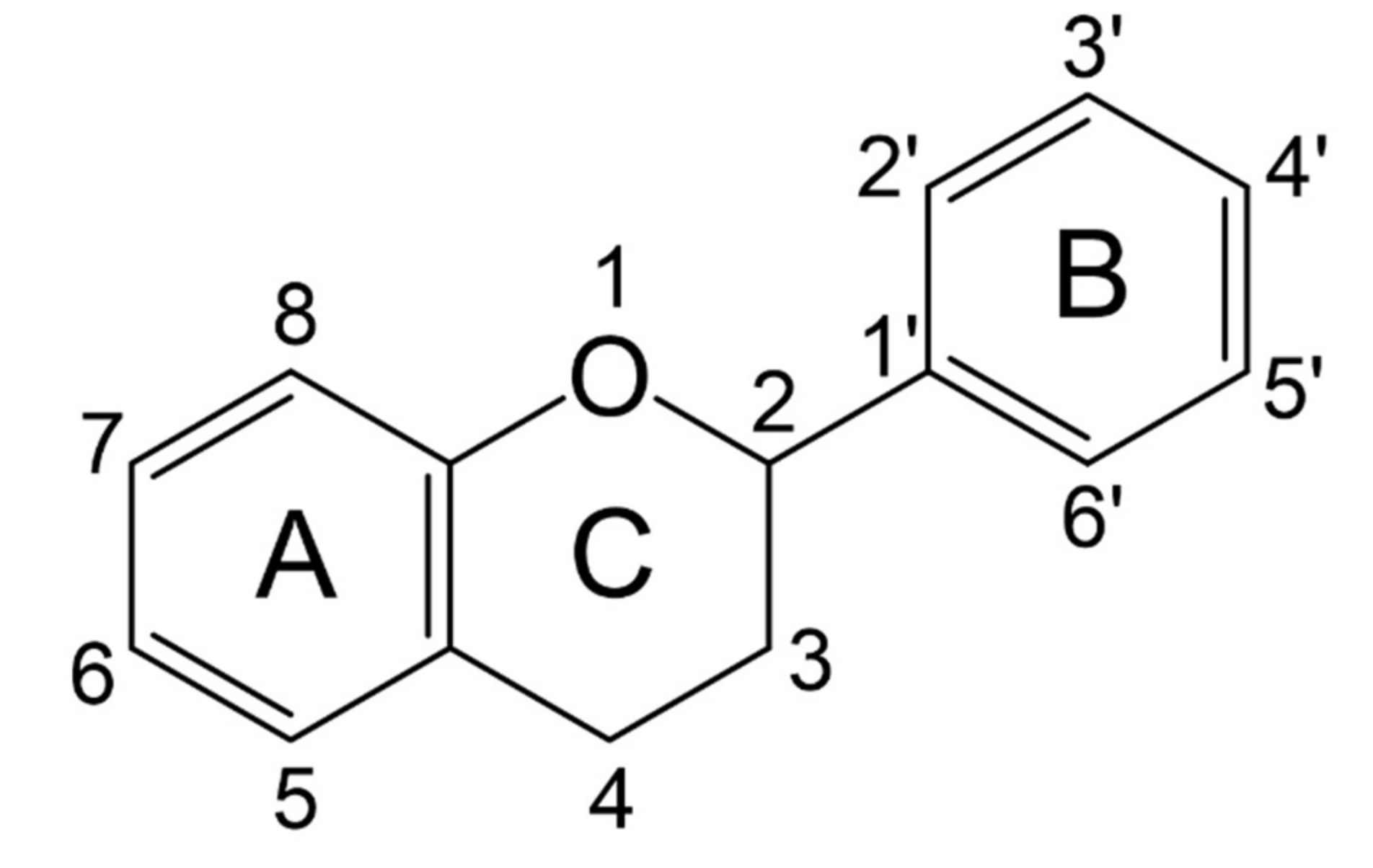
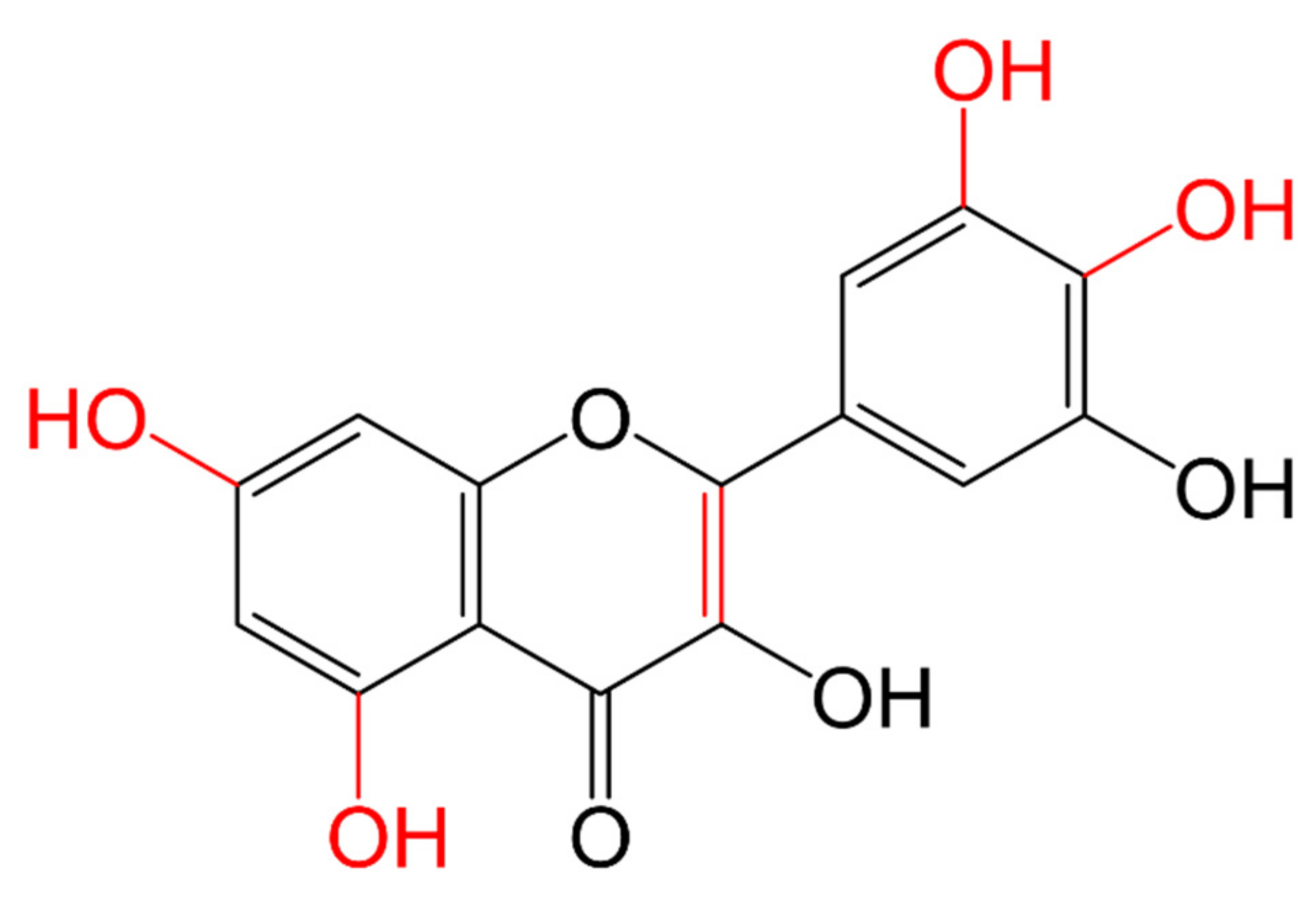
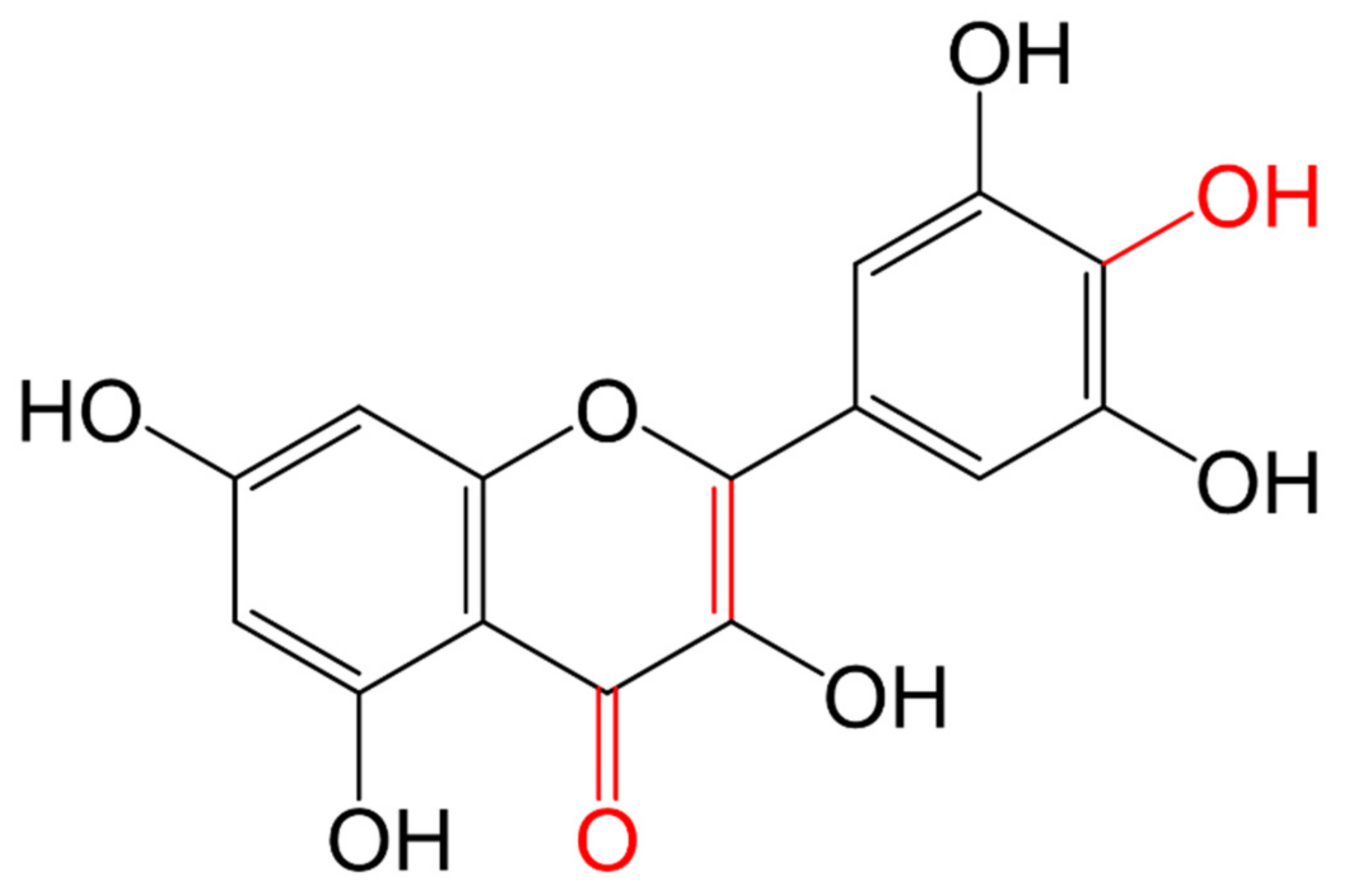
3.1.1. Classification and Structure
3.1.2. Flavonoids Diet Source and Absorption

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups of Flavonoids | Structure | Description | Diet Source and Site of Absorption | Examples | References |
|---|---|---|---|---|---|
| Chalcones |  | They have ring C open, without the oxygen bridge | Vegetables, fruits, and tea. Small intestine | Isoliquiritigenin, chalconaringenin, phloretin | [104,107,108] |
| Aurones |  | Rings A and C form a benzofuranone | Flowering plants. Intestine | Aureusidin, leptosidin | [106,107] |
| Anthocyanidins |  | They have at least one -OH group attached in position C3 and C4′, and full conjugated double bonds | Plant pigments of fruits, flowers, leaves, red wine. Small intestine | Cyanidin, malvidin, pelargonidin | [104,107,109] |
| Flavanols |  | They have a single bond between C2 and C3 and an -OH group at C3. They are also known as flavan-3-ols | Pears, apples, grapes, cocoa, and tea. Oral and small intestine | Catechin, epicatechin, epicatechin derivatives | [104,107] |
| Flavonols |  | They have the 3-hydroxyflavone backbone (-OH at C3) | Onions, broccoli, tea, spinach, kale, fruits. Proximal small intestine | Quercetin | [104,107,109] |
| Flavanones |  | They have a single bond between C2 and C3 and present a carbonyl group at C4 | Citrus fruits peels. Small intestine and colon | Eriodictyol, hesperetin, naringenin | [104,109] |
| Flavones |  | They present a carbonyl group at C4 and do not have an -OH group at C3 | Celery, garlic, and chamomile tea. Oral, small intestine, and colon | Apigenin, luteolin | [105,109] |
| Isoflavonoids or Isoflavones |  | Ring B (Phenyl group) is attached at C3 | Soy, and legumes. Colon | Genistein, daidzein | [104,109] |
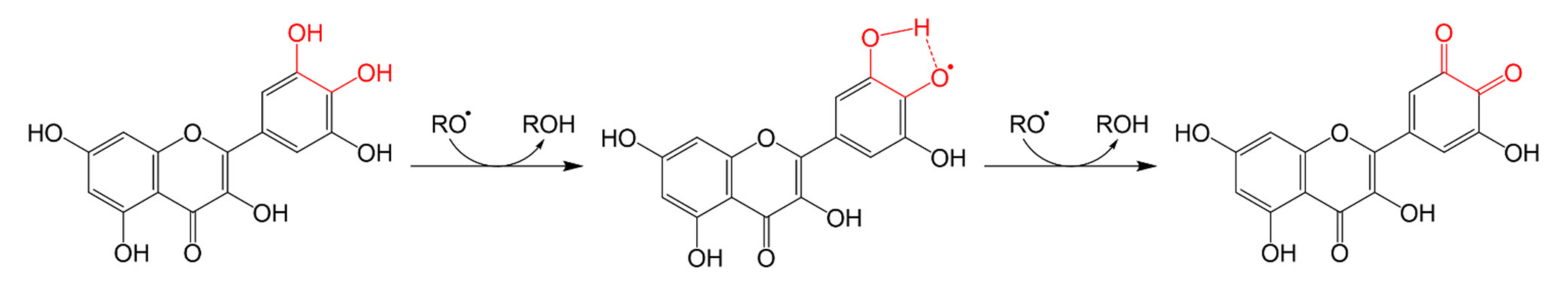
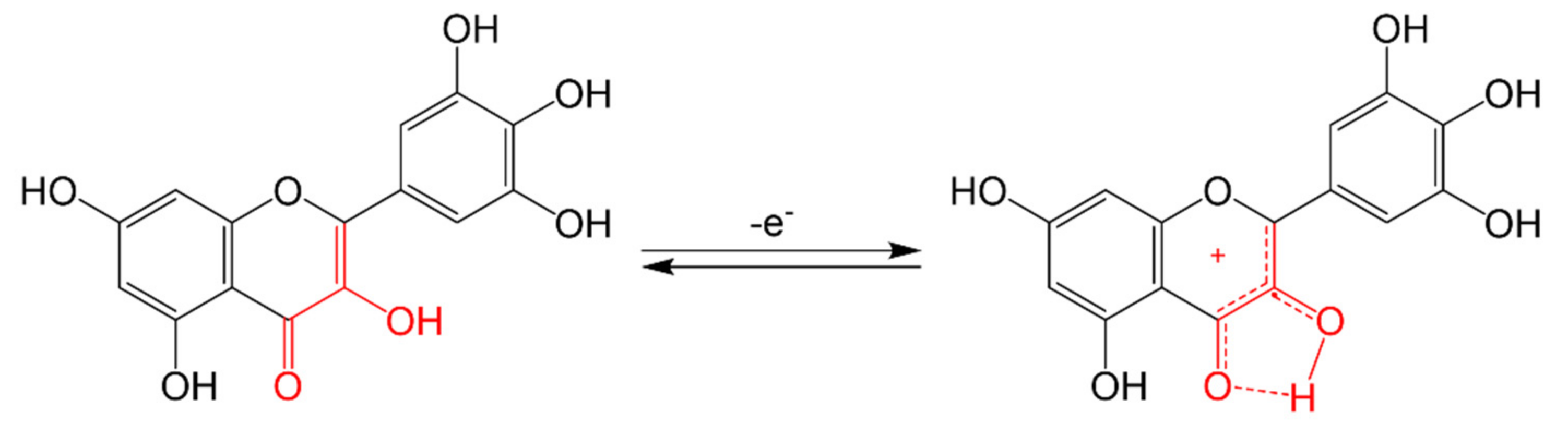
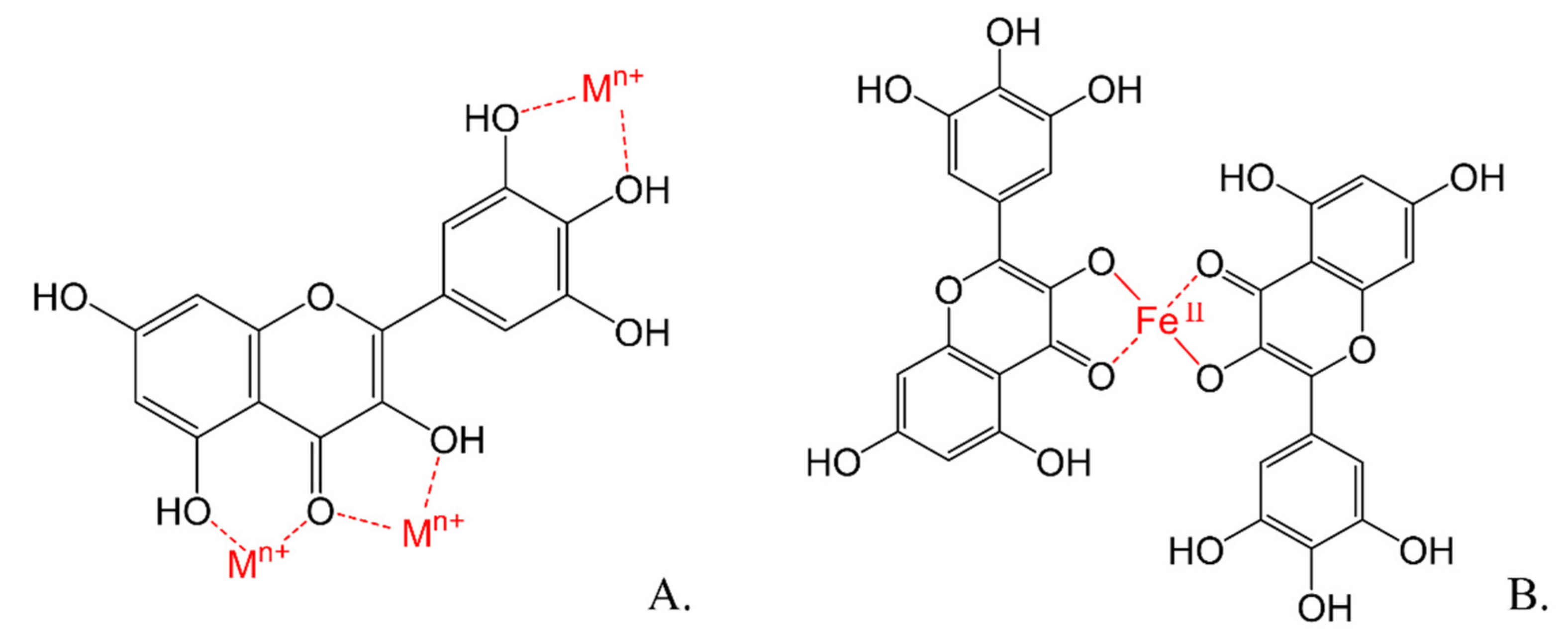
3.1.3. Antioxidant Mechanisms of Flavonoids
3.2. Effect of Flavonoids in Atherosclerosis
3.3. Effect of Flavonoids in VSMC’s Ion Channels
4. Effects of Flavonoids on Atherosclerosis through Modulation of Ion Channels in VSMC Activity
5. Future Perspectives in the Treatment
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Buckley, M.L.; Ramji, D.P. The influence of dysfunctional signaling and lipid homeostasis in mediating the inflammatory responses during atherosclerosis. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1498–1510. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S. Heart Disease and Stroke Statistics—2019 Update: A Report from the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- WHO—World Health Organization. World Heart Day 2017; WHO: Geneva, Switzerland, 2017; Available online: https://www.who.int/cardiovascular_diseases/world-heart-day-2017/en/ (accessed on 15 April 2021).
- Stocker, R.; Keaney, J.F. Role of Oxidative Modifications in Atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef]
- Galkina, E.; Ley, K. Immune and inflammatory mechanisms of atherosclerosis. Annu. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef]
- Wang, S.; Petzold, M.; Cao, J.; Zhang, Y.; Wang, W. Direct medical costs of hospitalizations for cardiovascular diseases in Shanghai, China: Trends and projections. Medicine 2015, 94, e837. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, B.N.; Wang, S.B.; Wang, S.H.; Du, G.H. Vasorelaxant effect of formononetin in the rat thoracic aorta and its mechanisms. J. Asian Nat. Prod. Res. 2012, 14, 46–54. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, H.; Wen, X.; Ho, C.-T.; Li, S. Citrus flavonoids and the intestinal barrier: Interactions and effects. Compr. Rev. Food Sci. Food Saf. 2021, 20, 225–251. [Google Scholar] [CrossRef]
- Rusznyák, S.; Szent-Györgyi, A. Vitamin P: Flavonols as Vitamins. Nature 1936, 138, 27. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Scarano, A.; Chieppa, M.; Santino, A. Looking at Flavonoid Biodiversity in Horticultural Crops: A Colored Mine with Nutritional Benefits. Plants 2018, 7, 98. [Google Scholar] [CrossRef]
- Bondonno, C.P.; Croft, K.D.; Ward, N.; Considine, M.J.; Hodgson, J.M. Dietary flavonoids and nitrate: Effects on nitric oxide and vascular function. Nutr. Rev. 2015, 73, 216–235. [Google Scholar] [CrossRef]
- Lakshmi, S.V.V.; Padmaja, G.; Kuppusamy, P.; Kutala, V.K. Oxidative Stress in Cardiovascular Disease. Indian J. Biochem. Biophys. 2009, 46, 421–440. [Google Scholar]
- Akbarali, H.I. Oxidative Stress and Ion Channels. Syst. Biol. Free Radic. Antioxid. 2014, 355–373. [Google Scholar] [CrossRef]
- Steinbach, S.K.; Husain, M. Vascular smooth muscle cell differentiation from human stem/progenitor cells. Methods 2016, 101, 85–92. [Google Scholar] [CrossRef]
- Oppi, S.; Lüscher, T.F.; Stein, S. Mouse Models for Atherosclerosis Research..Which Is My Line? Front. Cardiovasc. Med. 2019, 6, 46. [Google Scholar] [CrossRef]
- Farnier, M.; Davignon, J. Current and future treatment of hyperlipidemia: The role of statins. Am. J. Cardiol. 1998, 82, 3J–10J. [Google Scholar] [CrossRef]
- Insull, W., Jr. The pathology of Atherosclerosis: Plaque development and plaque responses to medical treatment. Am. J. Med. 2009, 122, S3–S14. [Google Scholar] [CrossRef]
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924. [Google Scholar] [CrossRef]
- Hajri, T. Effects of oxidized lipids and lipoproteins on cardiac function. Front. Biosci (Landmark Ed.) 2018, 23, 1822–1847. [Google Scholar] [CrossRef]
- Ross, R. Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Tabas, I.; Williams, K.J.; Borén, J. Subendothelial lipoprotein retention as the initiating process in Atherosclerosis: Update and therapeutic implications. Circulation 2014, 116, 1832–1844. [Google Scholar] [CrossRef]
- Wick, G.; Perschinka, H.; Milloning, G. Atherosclerosis as an autoimmune disease: An update. Trends Immunol. 2011, 22, 665–669. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Virella, G. Pathogenic role of modified LDL antibodies and immune complexes in atherosclerosis. J. Atheroscler. Thromb. 2013, 20, 743–754. [Google Scholar] [CrossRef]
- Morel, D.W.; Hessler, J.R.; Chisholm, G.M. Low density lipoprotein cytotoxicity induced by free radical peroxidation of lipid. J. Lipid Res. 1983, 24, 1070–1076. [Google Scholar] [CrossRef]
- Henriksen, T.; Mahoney, E.M.; Seinberg, D. Interactions of plasma lipoproteins with endothelial cells. Ann. New York Acad. Sci. 1982, 401, 102–116. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Bobryshev, Y.V.; Orekhov, A.N. Macrophage-mediated cholesterol handling in atherosclerosis. J. Cell. Mol. Med. 2016, 20, 17–28. [Google Scholar] [CrossRef]
- Gao, P.J.; Li, Y.; Sun, A.J.; Lin, J.J.; Ji, K.D.; Zhang, Y.Z.; Zhu, D.L. Differentiation of vascular myofibroblasts induced by transforming growth factor beta1 requires the involvement of protein kinase C alpha. J. Mol. Cell. Cardiol. 2003, 35, 1105–1112. [Google Scholar] [CrossRef]
- Camejo, G.; Fager, G.; Rosengren, B.; Hurt-Camejo, E.; Bondjers, G. Binding of low density lipoproteins by proteoglycans synthesized by proliferating and quiescent human arterial smooth muscle cells. J. Biol. Chem. 1993, 268, 14131–14137. [Google Scholar] [CrossRef]
- Skålén, K.; Gustafsson, M.; Rydberg, E.K.; Hultén, L.M.; Wiklund, O.; Innerarity, T.L.; Borén, J. Subendothelial retention of atherogenic lipoproteins in early atherosclerosis. Nature 2002, 417, 750–754. [Google Scholar] [CrossRef]
- Zhu, J.; Katz, R.J.; Quyummi, A.A. Association of serum antibodies to heat-shock protein 65 with coronary calcification levels. Suggestion of pathogen-triggered autoimmunity in early atherosclerosis. Circulation 2014, 109, 36–41. [Google Scholar] [CrossRef]
- Witztum, J.L.; Lichtman, A.H. The Influence of Innate and Adaptive Immune Responses on Atherosclerosis. Annu. Rev. Pathol. 2014, 9, 73–102. [Google Scholar] [CrossRef] [PubMed]
- Riganó, R.; Profumo, E.; Buttari, B. Heat shock proteins and autoimmunity in patients with carotid atherosclerosis. Ann. N. Y. Acad. Sci. 2016, 1107, 1–10. [Google Scholar] [CrossRef]
- Aboonabi, A.; Rose’Meyer, R.; Singh, I. The association between metabolic syndrome components and the development of atherosclerosis. J. Hum. Hypertens 2019, 1–12. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Adv. Cardiovasc. Dis 2017, 11, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Avan, A.; Tavakoly-Sany, S.B.; Ghayour-Mobarhan, M.; Rahimi, H.R.; Tajfard, M.; Ferns, G. Serum C-reactive protein in the prediction of cardiovascular diseases: Overview of the latest clinical studies and public health practice. J. Cell Physiol. 2018, 233, 8508–8525. [Google Scholar] [CrossRef]
- Speer, M.Y.; Yang, H.-Y.; Brabb, T.; Leaf, E.; Look, A.; Lin, W.-L.; Giachelli, C.M. Smooth Muscle Cells Give Rise to Osteochondrogenic Precursors and Chondrocytes in Calcifying Arteries. Circ. Res. 2009, 104, 733–741. [Google Scholar] [CrossRef]
- Taylor, A.J.; Burke, A.P.; O’Malley, P.G.; Farb, A.; Malcom, G.T.; Smialek, J.; Virmani, R. A comparison of the Framingham risk index, coronary artery calcification, and culprit plaque morphology in sudden cardiac death. Circulation 2000, 101, 1243–1248. [Google Scholar] [CrossRef]
- Hruska, K.A. Vascular Smooth Muscle Cells in the Pathogenesis of Vascular Calcification. Circ Res. 2009, 104, 710–711. [Google Scholar] [CrossRef] [PubMed]
- Rudijanto, A. The role of vascular smooth muscle cells on the pathogenesis of atherosclerosis. Acta Med. Indones. 2007, 39, 86–93. [Google Scholar]
- Sánchez-González, D.J.; Méndez-Bolaina, E.; Trejo-Bahena, N.I. Platelet-Rich Plasma Peptides: Key for Regeneration. Int. J. Pept. 2012, 532519. [Google Scholar] [CrossRef]
- Virmani, R.; Kolodgie, F.D.; Burke, A.P.; Farb, A.; Schwartz, S.M. Lessons from Sudden Coronary Death. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Yahagi, K.; Kolodgie, F.D.; Otsuka, F.; Finn, A.V.; Davis, H.R.; Joner, M.; Virmani, R. Pathophysiology of native coronary, vein graft, and in-stent Atherosclerosis. Nat. Rev. Cardiol. 2016, 13, 79–98. [Google Scholar] [CrossRef]
- Velican, C.; Velican, D. Intimal thickening in developing coronary arteries and its relevance to atherosclerotic involvement. Atherosclerosis 1976, 23, 345–355. [Google Scholar] [CrossRef]
- Ikari, Y.; McManus, B.M.; Kenyon, J.; Schwartz, S.M. Neonatal intima formation in the human coronary artery. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2036–2040. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Glagov, S.; Guyton, J.R.; Insull, W., Jr.; Rosenfeld, M.E.; Wissler, R.W. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1994, 89, 2462–2478. [Google Scholar] [CrossRef]
- Okura, Y.; Marijke, B.; Itabe, H.; Scheidegger, K.J.; Kalangos, A.; Delafontaine, P. Oxidized low-density lipoprotein is associated with apoptosis of vascular smooth muscle cells in human atherosclerotic plaques. Circulation 2000, 102, 2680–2686. [Google Scholar] [CrossRef]
- Tran-Lundmark, K.T.; Trana, P.K.; Paulsson-Berneb, G.; Fridénc, V.; Soininend, R.; Tryggvasone, K.; Wightf, T.N. Heparan Sulfate in Perlecan Promotes Mouse Atherosclerosis: Roles in Lipid Permeability, Lipid Retention, and Smooth Muscle Cell Proliferation. Circ. Res. 2008, 103, 43–52. [Google Scholar] [CrossRef]
- Clarke, M.C.H.; Littlewood, T.D.; Figg, N.; Maguire, J.J.; Davenport, A.P.; Goddard, M.; Bennett, M.R. Chronic Apoptosis of Vascular Smooth Muscle Cells Accelerates Atherosclerosis and Promotes Calcification and Medial Degeneration. Circ. Res. 2008, 102, 1529–1538. [Google Scholar] [CrossRef]
- Misra, A.; Feng, Z.; Chandran, R.R.; Kabir, I.; Rotllan, N.; Aryal, B.; Greif, D.M. Integrin beta3 regulates clonality and fate of smooth muscle-derived atherosclerotic plaque cells. Nat. Commun. 2018, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.S.; Hilgendorf, I.; Weber, G.F.; Theurl, I.; Iwamoto, Y.; Figueiredo, J.-L.; Swirski, F.K. Local proliferation dominates lesional macrophage accumulation in atherosclerosis. Nat. Med. 2013, 19, 1166–1172. [Google Scholar] [CrossRef]
- Nahrendorf, M. Myeloid cell contributions to cardiovascular health and disease. Nat. Med. 2018, 24, 711–720. [Google Scholar] [CrossRef]
- Qiao, J.-H.; Tripathi, J.; Mishra, N.K.; Tripathi, S. Role of Macrophage Colony-Stimulating Factor in Atherosclerosis, Studies of Osteopetrotic Mice. Am. J. Pathol. 1997, 150, 1687–1699. [Google Scholar]
- Swirski, F.K.; Pittet, M.J.; Kircher, M.F.; Aikawa, E.; Jaffer, F.A.; Libby, P.; Weissleder, R. Monocyte accumulation in mouse atherogenesis is progressive and proportional to extent of disease. Proc. Natl. Acad. Sci. USA 2006, 103, 10340–10345. [Google Scholar] [CrossRef]
- Campbell, J.; Rennick, R.E.; Kalevitch, S.G.; Campbell, G.R. Heparan Sulfate-Degrading Enzymes Induce Modulation of Smooth Muscle Phenotype. Exp. Cell Res. 1992, 200, 156–167. [Google Scholar] [CrossRef]
- Ait-Oufella, H.; Pouresmail, V.; Simon, T.; Blanc-Brude, O.; Kinugawa, K.; Merval, R.; Mallat, Z. Defective Mer Receptor Tyrosine Kinase Signaling in Bone Marrow Cells Promotes Apoptotic Cell Accumulation and Accelerates atherosclerosis. Arter. Thromb Vasc. Biol. 2008, 28, 1429–1431. [Google Scholar] [CrossRef]
- Clarke, M.C.H.; Talib, S.; Figg, N.L.; Bennett, M.R. Vascular Smooth Muscle Cell Apoptosis Induces Interleukin-1-Directed Inflammation: Effects of Hyperlipidemia-Mediated Inhibition of Phagocytosis. Circ. Res. 2010, 106, 363–372. [Google Scholar] [CrossRef]
- New, S.E.P.; Goettsch, C.; Aikawa, M.; Marchini, J.F.; Shibasaki, M.; Yabusaki, K.; Aikawa, E. Macrophage-Derived Matrix Vesicles: An Alternative Novel Mechanism for Microcalcification in Atherosclerotic Plaques. Circ. Res. 2013, 113, 72–77. [Google Scholar] [CrossRef]
- Kapustin, A.N.; Chatrou, M.L.L.; Drozdov, I.; Zheng, Y.; Davidson, S.M.; Soong, D.; Shanahan, C.M. Vascular Smooth Muscle Cell Calcification Is Mediated by Regulated Exosome Secretion. Circ. Res. 2015, 116, 1312–1323. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Goettsch, C.; Bertazzo, S.; Maldonado, N.; Ruiz, J.L.; Goh, W.; Aikawa, E. Genesis and growth of extracellular-vesicle-derived microcalcification in atherosclerotic plaques. Nat. Mater. 2016, 15, 335–343. [Google Scholar] [CrossRef]
- Davies, M.J.; Thomas, A. Thrombosis and acute coronary artery lesions in sudden cardiac ischemic death. N. Eng. J. Med. 1984, 310, 1137–1140. [Google Scholar] [CrossRef]
- Lassegue, B.; Clempus, R.E. Vascular NAD(P)H oxidases: Specific features, expression, and regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R277–R297. [Google Scholar] [CrossRef] [PubMed]
- Wolin, M.S. Interactions of Oxidants with Vascular Signaling Systems. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Sorescu, M.; Ushio-Fukai, M. NAD(P)H Oxidase, Role in Cardiovascular Biology and Disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef]
- Touyz, R.; Schiffrin, E. Ang II-stimulated superoxide production is mediated via phospholipase D in human vascular smooth muscle cells. Hypertension 1999, 34 Pt 2, 976–982. [Google Scholar] [CrossRef]
- Rao, G.N.; Berk, B.C. Active oxygen species stimulate vascular smooth muscle cell growth and proto-oncogene expression. Circ. Res. 1992, 70, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: From basic research to clinical application. Am. J. Med. 1991, 91 (Suppl. 3C), 31S–38S. [Google Scholar] [CrossRef]
- Tousoulis, D.; Kampoli, A.M.; Tentolouris, C.; Papageorgiou, N.; Stefanadis, C. The role of nitric oxide on endothelial function. Curr. Vasc. Pharmacol. 2012, 10, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Bogeski, I.; Niemeyer, B.A. Redox Regulation of Ion Channels. Antioxid. Redox. Signal. 2014, 21, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, N.; Hoshi, T.; Heinemann, S.H. Oxidative Modulation of Voltage-Gated Potassium Channels. Antioxid. Redox Signaling. 2014, 21, 933–952. [Google Scholar] [CrossRef]
- Trebak, M. STIM/Orai signalling complexes in vascular smooth muscle. J. Physiol. 2012, 590, 4201–4208. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell. Dev. Biol. 2000, 16, 521–555. [Google Scholar] [CrossRef]
- Roderick, H.L.; Berridge, M.J.; Bootman, M.D. Calcium-induced calcium release. Curr. Biol. 2003, 13, R425. [Google Scholar] [CrossRef]
- Brenner, R.; Peréz, G.J.; Bonev, A.D.; Eckman, D.M.; Kosek, J.C.; Wiler, S.W.; Aldrich, R.W. Vasoregulation by the beta1 subunit of the calcium-activated potassium channel. Nature 2000, 407, 870–876. [Google Scholar] [CrossRef]
- Ghosh, D.; Syed, A.U.; Prada, M.P.; Nystoriak, M.A.; Santana, L.F.; Nieves-Cintrón, M.; Navedo, M.F. Calcium Channels in Vascular Smooth Muscle. Vasc. Pharmacol. Smooth Muscle 2017, 49–87. [Google Scholar] [CrossRef]
- Jentsch, T.J.; Stein, V.; Weinreich, F.; Zdebik, A.A. Molecular structure and physiological function of chloride channels. Physiol. Rev. 2002, 82, 503–568. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Yamazaki, J. Chloride channels and their functional roles in smooth muscle tone in the vasculature. Jpn. J. Pharmacol. 2001, 85, 351–357. [Google Scholar] [CrossRef]
- Benham, C.D.; Hess, P.; Tsien, R.W. Two types of calcium channels in single smooth muscle cells from rabbit ear artery studied with whole-cell and single-channel recordings. Circ. Res. 1987, 61, I10–116. [Google Scholar]
- Keef, K.D.; Hume, J.R.; Zhong, J. Regulation of cardiac and smooth muscle Ca (2+) channels (Ca(V)1.2a, b) by protein kinases. Am. J. Physiol Cell Physiol. 2001, 281, C1743–C1756. [Google Scholar] [CrossRef]
- Kuo, I.Y.; Howitt, L.; Sandow, S.L.; McFarlane, A.; Hansen, P.B.; Hill, C.E. Role of T-type channels in vasomotor function: Team player or chameleon? Pflugers Arch 2014, 466, 767–779, PubMed: 24482062. [Google Scholar] [CrossRef]
- Gebremedhin, D.; Kaldunski, M.; Jacobs, E.R.; Harder, D.R.; Roman, R.J. Coexistence of two types of Ca (2+)-activated K+ channels in rat renal arterioles. Am. J. Physiol. Ren. Physiol. 1996, 270, F69–F81. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Toyama, K.; Lemaitre, V.; Takai, J.; Fan, F.; Jenkins, D.P.; Wulff, H.; Gutterman, D.D.; Park, F.; Miura, H. The intermediate conductance calcium-activated potassium channel KCa3.1 regulates vascular smooth muscle cell proliferation via controlling calcium-dependent signaling. J. Biol. Chem. 2013, 288, 15843–15853. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gutterman, D.D.; Rusch, N.J.; Bubolz, A.; Liu, Y. Nitration and functional loss of voltage-gated K+ channels in rat coronary microvessels exposed to high glucose. Diabetes 2004, 53, 2436–2442. [Google Scholar] [CrossRef]
- Owsianik, G.; Talavera, K.; Voets, T.; Nilius, B. Permeation and selectivity of TRP channels. Annu. Rev. Physiol. 2006, 68, 685–717. [Google Scholar] [CrossRef]
- Chalmers, S.; Olson, M.; Macmillan, D.; Rainbow, R.; Mccarron, J. Ion channels in smooth muscle: Regulation by the sarcoplasmic reticulum and mitochondria. Cell Calcium 2007, 42, 447–466. [Google Scholar] [CrossRef]
- Scolaro, B.; Kim, S.J.H.; de Castro, I.A. Bioactive compounds as an alternative for drug co-therapy: Overcoming challenges in cardiovascular disease prevention. Crit. Rev. Food Sci. Nutr. 2018, 58, 958–971. [Google Scholar] [CrossRef]
- Belinky, P.A.; Aviram, M.; Fuhrman, B.; Rosenblat, M.; Vaya, J. The antioxidative effects of the isoflavan glabridin on endogenous constituents of LDL during its oxidation. Atherosclerosis 1998, 137, 49–61. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Martínez-González, M.A. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef]
- Wang, B.; Liao, P.-P.; Liu, L.-H.; Fang, X.; Li, W.; Guan, S.-M. Baicalin and geniposide inhibit the development of atherosclerosis by increasing Wnt1 and inhibiting dickkopf-related protein-1 expression. J. Geriatr. Cardiol. 2016, 13, 846–854. [Google Scholar] [CrossRef]
- Sun, G.B.; Qin, M.; Ye, J.X.; Pan, R.L.; Meng, X.B.; Wang, M. Inhibitory effects of myricitrin on oxidative stress-induced endothelial damage and early Atherosclerosis in ApoE-/- mice. Toxicol. Appl. Pharmacol. 2013, 271, 114–126. [Google Scholar] [CrossRef]
- Xiao, H.B.; Lu, X.Y.; Sun, Z.L.; Zhang, H.B. Kaempferol regulates OPN-CD44 pathway to inhibit the atherogenesis of apolipoprotein E deficient mice. Toxicol. Appl. Pharmacol. 2011, 257, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Proudfoot, J.M.; Hodgson, J.M.; McKinley, A.J.; Hime, N.; Magat, M.; Croft, K.D. Specific dietary polyphenols attenuate atherosclerosis in apolipoprotein E-knockout mice by alleviating inflammation and endothelial dysfunction. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Belinky, P.; Vaya, J.; Levy, R.; Hayek, T.; Coleman, R. Macrophage enrichment with the isoflavan glabridin inhibits NADPH oxidase-induced cell-mediated oxidation of low density lipoprotein. A possible role for protein kinase C. J. Biol. Chem. 1999, 274, 13790–13799. [Google Scholar] [CrossRef]
- Ciumărnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, S.C.; Răchis, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.G.; Para, I.; et al. The Effects of Flavonoids in Cardiovascular Diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef]
- Hamid, A.A.; Aminuddin, A.; Yunus, M.H.M.; Murthy, J.K.; Hui, C.K.; Ugusman, A. Antioxidative and anti-inflammatory activities of Polygonum minus: A review of literature. Rev. Cardiovasc. Med. 2020, 21, 275–287. [Google Scholar]
- Ma, C.; Zhang, J.; Yang, S.; Hua, Y.; Su, J.; Shang, Y.; Fan, G. Astragalus Flavone Ameliorates Atherosclerosis and Hepatic Steatosis via Inhibiting Lipid-Disorder and Inflammation in apoE-/- Mice. Front. Pharmacol. 2020, 11, 610550. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, Y.; Li, D.; Xie, T.; Li, Y.; Li, J.; Wei, G. Integrating Network Pharmacology and Component Analysis Study on Anti-Atherosclerotic Mechanisms of Total Flavonoids of Engelhardia roxburghiana Leaves in Mice. Chem Biodivers. 2020, 17, e1900629. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Shi, X.; Lv, T.; Yuan, Q.; Zhu, J. Anti-proliferative and anti-migratory effects of Scutellaria strigillosa Hemsley extracts against vascular smooth muscle cells. J. Ethnopharmacol. 2019, 235, 155–163. [Google Scholar] [CrossRef]
- Tabares-Guevara, J.H.; Lara-Guzmán, O.J.; Londoño-Londoño, J.A. Natural Biflavonoids Modulate Macrophage-Oxidized LDL Interaction in Vitro and Promote Atheroprotection in Vivo. Front. Immunol. 2017, 8, 923. [Google Scholar] [CrossRef] [PubMed]
- Pangestika, I.; Oksal, E.; Tengku Muhammad, T.S.; Amir, H.; Syamsumir, D.F.; Wahid, M.E.A.; Andriani, Y. Inhibitory effects of tangeretin and trans-ethyl caffeate on the HMG-CoA reductase activity: Potential agents for reducing cholesterol levels. Saudi Journal of Biological Sciences. Saudi. J. Biol. Sci. 2020, 27, 1947–1960. [Google Scholar] [CrossRef]
- McNaught, A.D.; Wilkinson, A. IUPAC Compendium of Chemical Terminology, 2nd ed.; IUPAC-Blackwell Scientific: Oxford, UK, 1997. [Google Scholar] [CrossRef]
- Ekalu, A.; Habila, J.D. Flavonoids: Isolation, characterization, and health benefits. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 45. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The Bioavailability, Transport, and Bioactivity of Dietary Flavonoids: A Review from a Historical Perspective. Compr. Rev. Food Sci. Food Saf. 2018. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef]
- Hadni, H.; Elhallaoui, M. 3D-QSAR, docking and ADMET properties of aurone analogues as antimalarial agents. Heliyon 2020, 6, e03580. [Google Scholar] [CrossRef]
- Maaliki, D.; Shaito, A.A.; Pintus, G.; El-Yazbi, A.; Eid, A.H. Flavonoids in hypertension: A brief review of the underlying mechanisms. Curr. Opin. Pharmacol. 2019, 45, 57–65. [Google Scholar] [CrossRef]
- Qi, J.; Cui, J.; Mi, B.; Yan, X.; Xu, W.; Ma, H. Isoliquiritigenin Inhibits Atherosclerosis by Blocking TRPC5 Channel Expression. Cardiovasc Ther. 2020, 2020, 1926249. [Google Scholar] [CrossRef]
- Behl, T.; Bungau, S.; Kumar, K.; Zengin, G.; Khan, F.; Kumar, A.; Mosteanu, D.-E. Pleotropic Effects of Polyphenols in Cardiovascular System. Biomed. Pharmacother. 2020, 130, 110714. [Google Scholar] [CrossRef]
- Fuhrman, B.; Aviram, M. Flavonoids protect LDL from oxidation and attenuate atherosclerosis. Curr. Opin. Lipidol. 2001, 12, 41–48. [Google Scholar] [CrossRef]
- Aviram, M.; Fuhrman, B. Wine flavonoids protect against LDL oxidation and Atherosclerosis. Ann. N. Y. Acad. Sci. 2002, 957, 146–161. [Google Scholar] [CrossRef]
- Faggio, C.; Sureda, A.; Morabito, S. Flavonoids and platelet aggregation: A brief review. Eur. J. Pharmacol. 2017, 807, 91–101. [Google Scholar] [CrossRef]
- Enciso, E.; Arroyo, J. Efecto Antiinflamatorio y Antioxidante de los Flavonoides de las Hojas de Jungia rugosa Less (matico de puma) en un Modelo Experimental en Ratas. Rev. An. Fac. Med. 2011, 72, 231–237. [Google Scholar] [CrossRef][Green Version]
- Havsteen, B. The Biochemistry and Medical Significance of the Flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Dolatabadi, J. Molecular aspects on the interaction of quercetin and its metal complexes with DNA. Int. J. Biol. Macromol. 2011, 48, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Tarahovsky, Y.S.; Kim, Y.A.; Yagolnik, E.A.; Muzafarov, E.N. Flavonoid-membrane interactions: Involvement of flavonoid-metal complexes in raft signaling. Biochim. Biophys. Acta. 2014, 1838, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J. Antioxidant activities of flavonoids as bioactive components of food. Biochem. Soc. Trans. 1996, 24, 790–795. [Google Scholar] [CrossRef]
- Rice-Evans, C. Flavonoid Antioxidants. Curr. Med. Chem. 2001, 8, 797–807. [Google Scholar] [CrossRef]
- Lemańska, K.; Szymusiak, H.; Tyrakowska, B.; Zieliński, R.; Soffers, A.E.; Rietjens, I.M. The influence of pH on antioxidant properties and the mechanism of antioxidant action of hydroxyflavones. Free Radic. Biol. Med. 2001, 31, 869–881. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Costantino, L.; Rastelli, G.; Albasini, A. A Rational Approach to the Design of Flavones as Xanthine Oxidase Inhibitors. Eur. J. Med. Chem. 1996, 31, 693–699. [Google Scholar] [CrossRef]
- Cotelle, N.; Bernier, J.L.; Catteau, J.P.; Pommery, J.; Wallet, J.C.; Gaydou, E.M. Antioxidant Properties of Hydroxy-flavones. Free Radic. Biol. Med. 1996, 20, 35–43. [Google Scholar] [CrossRef]
- Hayashi, T.; Sawa, K.; Kawasaki, M.; Arisawa, M.; Shimizu, M.; Morita, N. Inhibition of Cow’s Milk Xanthine Oxidase by Flavonoids. J. Nat. Prod. 1988, 51, 345–348. [Google Scholar] [CrossRef]
- Sadik, C.D.; Sies, H.; Schewe, T. Inhibition of 15-lipoxygenases by flavonoids: Structure–activity relations and mode of action. Biochem. Pharmacol. 2003, 65, 773–781. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Biomembr. Part C Biol. 1978, 302–310. [Google Scholar] [CrossRef]
- Ratty, A.K.; Das, N.P. Effects of flavonoids on nonenzymatic lipid peroxidation: Structure-activity relationship. Biochem. Med. Metab. Biol. 1988, 39, 69–79. [Google Scholar] [CrossRef]
- Hertog, M.G.; Feskens, E.J.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Cirillo, P.; Conte, S.; Cimmino, G.; Pellegrino, G.; Ziviello, F.; Barra, G.; Trimarco, B. Nobiletin inhibits oxidized-LDL mediated expression of Tissue Factor in human endothelial cells through inhibition of NF-κB. Biochem. Pharmacol. 2017, 128, 26–33. [Google Scholar] [CrossRef]
- Stainer, A.R.; Sasikumar, P.; Bye, A.P.; Unsworth, A.J.; Holbrook, L.M.; Tindall, M.; Lovegrove, J.-A.; Gibbins, J.M. The Metabolites of the Dietary Flavonoid Quercetin Possess Potent Antithrombotic Activity, and Interact with Aspirin to Enhance Antiplatelet Effects. TH Open. 2019, 3, e244–e258. [Google Scholar] [CrossRef]
- Fuhrman, B.; Volkova, N.; Coleman, R.; Aviram, M. Grape Powder Polyphenols Attenuate Atherosclerosis Development in Apolipoprotein E Deficient (E0) Mice and Reduce Macrophage Atherogenicity. J. Nutr. 2005, 135, 722–728. [Google Scholar] [CrossRef]
- Marunaka, Y.; Marunaka, R.; Sun, H.; Yamamoto, T.; Kanamura, N.; Inui, T.; Taruno, A. Actions of Quercetin, a Polyphenol, on Blood Pressure. Molecules 2017, 22, 209. [Google Scholar] [CrossRef]
- Bondonno, N.; Bondonno, C.; Rich, L.; Ward, N.; Hodgson, J.; Croft, K. Flavonoid-rich apple improves endothelial function in individuals at risk for cardiovascular disease. J. Nutr. Intermed. Metab. 2017, 8, 79–80. [Google Scholar] [CrossRef]
- Nardi, A.; Calderone, V.; Chericoni, S.; Morelli, I. Natural modulators of large-conductance calcium-activated potassium channels. Planta Med. 2003, 69, 885–892. [Google Scholar] [PubMed]
- Calderone, V.; Chericoni, S.; Martinelli, C.; Testai, L.; Nardi, A.; Morelli, I.; Martinotti, E. Vasorelaxing effects of flavonoids: Investigation on the possible involvement of potassium channels. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 370, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Liu, L.; Guo, X.; Zhang, S.; Wang, J.; Zhou, F.; Yao, P. Quercetin attenuates high fat diet-induced atherosclerosis in apolipoprotein E knockout mice: A critical role of NADPH oxidase. Food Chem. Toxicol. 2017, 105, 22–33. [Google Scholar] [CrossRef]
- Okawa, M.; Kinjo, J.; Nohara, T.; Ono, M. DPPH (1,1-diphenyl-2- picrylhydrazyl) radical scavenging activity of flavonoids obtained from some medicinal plants. Biol. Pharm. Bull. 2001, 24, 1202–1205. [Google Scholar] [CrossRef]
- Dasilva, G.; Pazos, M.; García-Egido, E.; Gallardo, J.M.; Ramos-Romero, S.; Torres, J.L.; Medina, I. A lipidomic study on the regulation of inflammation and oxidative stress targeted by marine ω-3 PUFA and polyphenols in high-fat high-sucrose diets. J. Nutr. Biochem. 2017, 43, 53–67. [Google Scholar] [CrossRef]
- Sobottka, A.M.; Werner, W.; Blaschke, G.; Kiefer, W.; Nowe, U.; Dannhardt, G.; Schapoval, E.E.; Schenkel, E.P.; Scriba, G.K. Effect of flavonol derivatives on the carrageenin-induced paw edema in the rat and inhibition of cyclooxygenase-1 and 5- lipoxygenase in vitro. Arch. Pharm (Weinh.) 2000, 333, 205–210. [Google Scholar] [CrossRef]
- Welton, A.F.; Tobias, L.D.; Fiedler-Nagy, C.; Anderson, W.; Hope, W.; Meyers, K.; Coffey, J.W. Effect of flavonoids on arachidonic acid metabolism. Prog. Clin. Biol. Res. 1986, 213, 231–242. [Google Scholar]
- Elliott, A.J.; Scheiber, S.A.; Thomas, C.; Pardini, R.S. Inhibition of glutathione reductase by flavonoids. Biochem. Pharmacol. 1992, 44, 1603–1608. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 125124. [Google Scholar] [CrossRef]
- Zhang, X.; Han, X.; Zhang, P.; Zhou, T.; Chen, Y.; Jin, J.; Ma, X. Morin attenuates oxidized low-density lipoprotein-mediated injury by inducing autophagy via activating AMPK signalling in HUVECs. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1053–1060. [Google Scholar] [CrossRef]
- Baba, S.; Natsume, M.; Yasuda, A.; Nakamura, Y.; Tamura, T.; Osakabe, N.; Kanegae, M.; Kondo, K. Plasma LDL and HDL cholesterol and oxidized LDL concentrations are altered in normo and hypercholesterolemic humans after intake of different levels of cocoa powder. J. Nutr. 2007, 137, 1436–1441. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Allister, E.M.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Edwards, J.Y.; Huff, M.W. Naringenin Prevents Dyslipidemia, Apolipoprotein B Overproduction, and Hyperinsulinemia in LDL Receptor-Null Mice with Diet-Induced Insulin Resistance. Diabetes 2009, 58, 2198–2210. [Google Scholar] [CrossRef]
- Wittemer, S.M.; Ploch, M.; Windeck, T.; Müller, S.C.; Drewelow, B.; Derendorf, H.; Veit, M. Bioavailability and pharmacokinetics of caffeoylquinic acids and flavonoids after oral administration of Artichoke leaf extracts in humans. Phytomedicine 2005, 12, 28–38. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef]
- Duarte, J.; Francisco, V.; Perez-Vizcaino, F. Modulation of nitric oxide by flavonoids. Food Funct. 2014, 5, 1653–1668. [Google Scholar] [CrossRef]
- Fusi, F.; Spiga, O.; Trezza, A.; Sgaragli, G.; Saponara, S. The surge of flavonoids as novel, fine regulators of cardiovascular Cav channels. Eur. J. Pharm. 2017, 796, 158–174. [Google Scholar] [CrossRef]
- Marunaka, Y. Actions of quercetin, a flavonoid, on ion transporters: Its physiological roles. Ann. N. Y. Acad. Sci. 2017, 1398, 142–151. [Google Scholar] [CrossRef]
- Fusi, F.; Trezza, A.; Tramaglino, M.; Sgaragli, G.; Saponara, S.; Spiga, O. The beneficial health effects of flavonoids on the cardiovascular system: Focus on K+ channels. Pharmacol. Res. 2020, 152, 104625. [Google Scholar] [CrossRef] [PubMed]
- Figtree, G.A.; Griffiths, H.; Lu, Y.Q.; Webb, C.M.; MacLeod, K.; Collins, P. Plant-derived estrogens relax coronary arteries in vitro by a calcium antagonistic mechanism. J. Am. Coll. Cardiol. 2000, 35, 1977–1985. [Google Scholar] [CrossRef]
- Pan, Z.; Feng, T.; Shan, L.; Cai, B.; Chu, W.; Niu, H. Scutellarin-induced endothelium-independent relaxation in rat aorta. Phytother. Res. 2008, 22, 1428–1433. [Google Scholar] [CrossRef]
- Wani, S.A.; Iqbal, H.; Basir, S.F. Mechanism of Flavonoids action in smooth muscle relaxation. WJPPS 2017, 6, 514–550. [Google Scholar] [CrossRef]
- El Haouari, M.; Rosado, J.A. Modulation of Platelet Function and Signaling by Flavonoids. Mini Rev. Med. Chem. 2011, 11, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Campos-Toimil, M.; Orallo, F. Effects of (–)-epigallocatechin-3-gallate in Ca2+-permeable non-selective cation channels and voltage-operated Ca2+ channels in vascular smooth muscle cells. Life Sci. 2007, 80, 2147–2153. [Google Scholar] [CrossRef]
- Alvarez-Castro, E.; Campos-Toimil, M.; Orallo, F. (-)-Epigallocatechin-3-gallate induces contraction of the rat aorta by a calcium influx-dependent mechanism. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.Y.; Park, S.H.; Bae, J.H.; Cho, H.C.; Lim, J.G.; Park, W.S.; Song, D.K. Uncoupling by (–)-epigallocatechin-3-gallate of ATP-sensitive potassium channels from phosphatidylinositol polyphosphates and ATP. Pharmacol. Res. 2007, 56, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Fusi, F.; Saponara, S.; Pessina, F.; Gorelli, B.; Sgaragli, G. Effects of quercetin and rutin on vascular preparations: A comparison between mechanical and electrophysiological phenomena. Eur. J. Nutr. 2003, 42, 10–17. [Google Scholar] [CrossRef]
- Xu, Y.C.; Leung, G.P.H.; Wong, P.Y.D.; Vanhoutte, P.M.; Man, R.Y.K. Kaempferol stimulates large conductance Ca2+-activated K+ (BKCa) channels in human umbilical vein endothelial cells via a cAMP/PKA-dependent pathway. Br. J. Pharmacol. 2008, 154, 1247–1253. [Google Scholar] [CrossRef]
- Sun, X.; Ding, J.; Li, H.; Pan, N.; Gan, L.; Yang, X.L.; Xu, H.B. Activation of large-conductance calcium-activated potassium channels by puerarin: The underlying mechanism of puerarin-mediated vasodilation. J. Pharmacol. Exp. Ther. 2007, 323, 391–397. [Google Scholar] [CrossRef]
- Côrtes, S.F.; Rezende, B.A.; Corriu, C.; Medeiros, I.A.; Teixeira, M.M.; Lopes, M.J.; Lemos, V.S. Pharmacological evidence for the activation of potassium channels as the mechanism involved in the hypotensive and vasorelaxant effect of dioclein in rat small resistance arteries. Br. J. Pharmacol. 2001, 133, 849–858. [Google Scholar] [CrossRef]
- Saponara, S.; Testai, L.; Iozzi, D. (+/–)-Naringenin as large conductance Ca (2+)-activated K+ (BKCa) channel opener in vascular smooth muscle cells. Br. J. Pharm. 2006, 149, 1013–1021. [Google Scholar] [CrossRef]
- Gao, Q.; Yang, B.; Ye, Z.G.; Wang, J.; Bruce, I.C.; Xia, Q. Opening the calcium-activated potassium channel participates in the cardioprotective effect of puerarin. Eur. J. Pharmacol. 2007, 574, 179–184. [Google Scholar] [CrossRef]
- Cogolludo, A.; Frazziano, G.; Briones, A.M.; Cobeño, L.; Moreno, L.; Lodi, F.; Perez-Vizcaino, F. The dietary flavonoid quercetin activates BKCa currents in coronary arteries via production of H2O2. Role in vasodilatation. Cardiovasc. Res. 2007, 73, 424–431. [Google Scholar] [CrossRef]
- Ko, E.A.; Park, W.S.; Son, Y.K. The effect of tyrosine kinase inhibitor genistein on voltage-dependent K+ channels in rabbit coronary arterial smooth muscle cells. Vascul. Pharmacol. 2009, 50, 51–56. [Google Scholar] [CrossRef]
- Hernández-Abreu, O.; Castillo-España, P.; León-Rivera, I.; Ibarra-Barajas, M.; Villalobos-Molina, R.; González-Christen, J.; Estrada-Soto, S. Antihypertensive and vasorelaxant effects of tilianin isolated from Agastache mexicana are mediated by NO/cGMP pathway and potassium channel opening. Biochem. Pharmacol. 2009, 78, 54–61. [Google Scholar] [CrossRef]
- Zhu, X.; Fang, L.; Li, Y.; Du, G. Endothelium-dependent and –independent relaxation induced by pinocembrin in rat aortic rings. Vascul. Pharmacol. 2007, 46, 160–165. [Google Scholar] [CrossRef]
- Adaramoye, O.A.; Medeiros, I.A. Endothelium-independent vasodilation induced by kolaviron, a biflavonoid complex from Garcinia kola seeds, in rat superior mesenteric arteries. J. Smooth Muscle Res. 2009, 45, 39–53. [Google Scholar] [CrossRef]
- Kang, D.G.; Yin, M.H.; Oh, H.; Lee, D.H.; Lee, H.S. Vasorelaxation by amentoflavone isolated from Selaginella tamariscina. Planta Med. 2004, 70, 718–722. [Google Scholar] [CrossRef]
- Jiang, H.; Xia, Q.; Wang, X.; Song, J.; Bruce, I.C. Luteolin induces vasorelaxion in rat thoracic aorta via calcium and potassium channels. Pharmazie 2005, 60, 444–447. [Google Scholar]
- Fusi, F.; Cavalli, M.; Mulholland, D.; Crouch, N.; Coombes, P.; Dawson, G.; Saponara, S. Cardamonin is a bifunctional vasodilator that inhibits Ca(v)1.2 current and stimulates K(Ca)1.1 current in rat tail artery myocytes. J. Pharmacol. Exp. Ther. 2010, 332, 531–540. [Google Scholar] [CrossRef]
- Siasos, G.; Tousoulis, D.; Tsigkou, V.; Kokkou, E.; Oikonomou, E.; Vavuranakis, M.; Stefanadis, C. Flavonoids in Atherosclerosis: An Overview of Their Mechanisms of Action. Curr. Med. Chem. 2013, 20, 2641–2660. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Zhou, X.B.; Chen, C.; Mao, W. Systematic Investigation of Quercetin for Treating Cardiovascular Disease Based on Network Pharmacology. Comb. Chem. High. Throughput Screen. 2019, 22, 411–420. [Google Scholar] [CrossRef] [PubMed]









| Ion Channel | Molecular Target of ROS | Physiological Effect | Membrane Potential Effect | Effect on the Disease | Expression |
|---|---|---|---|---|---|
| L-type Cav 18–25 pS (Cav 1.2b) | Cysteine oxidation within alpha subunit, C1789, C1790, C1810 on alpha1C | ↓ Ca2+ currents | Lack of depolarizing stimuli. Reduced sensitivity toward physiological stimulation | Reduction in dihydropyridine binding sites. Less vasocontraction | Cav1.2b expression is not affected. Smooth muscle contractility alterations |
| T-type Cav 8 pS (Cav 3.1–3.3) | Extracellular cysteine oxidation | ↓ Ca2+ currents | Non-significant effects on membrane potential | Diminish Vasocontraction | ND |
| BKca 200–300 pS | Slo1 Cysteines oxidation residues: C14, C141, C430, C615, C911 and methionine residues: M536, M712, M739 | Decreased potassium outward currents ↓ [Ca2+]i | Abnormally depolarized resting membrane potential | Decreased Slo channel activity. Inactivation. Alters membrane function, losing homeostasis, and leading to disease. Less vasocontraction | Downregulated |
| IKCa 20–90 pS | ND | Decreased potassium outward currents ↓ [Ca2+]i | Abnormally depolarized resting membrane potential | SMCs proliferation and migration | Upregulated |
| SKCa 5–20 pS | ND | Decreased potassium outward currents ↓ [Ca2+]i | Abnormally depolarized resting membrane potential | SMCs proliferation and migration | Upregulated |
| Kv1.X | ND | Decreased currents ↓ [Ca2+]i | Abnormally depolarized resting membrane potential | Alters membrane function, losing homeostasis and leading to disease. Less vasocontraction | ND |
| TRPM4 | Oxidation of a Cysteine terminally, C1093 | ↑ Ca2+, Na+ influx desensitization | Membrane depolarization | ↑ Necrosis and cell death Opening of Cav1.2 channels resulting in SMC contraction | Cell-specific upregulated |
| TRPC3 | ND | ↑ Na+ influx | Membrane depolarization | ND | Cell-specific upregulated |
| TRPC6 | ND | ↑ Na+, Ca2+ influx | Membrane depolarization | Opening of Cav1.2 channels resulting in SMC contraction | Cell-specific upregulated |
| Plant | Flavonoids | Therapeutic Effect | References |
|---|---|---|---|
| Polygonum minus (Persicaria minor) (pygmy smartweed, small water pepper, or swamp willow weed) | Myricetin, quercetin, methyl-flavonol | Antioxidant, anti-inflammatory | [95,96] |
| Ajuga Iva(L.) (búgula almizclada) | Naringenin, apigenin-7-O-neohesperidoside | Antioxidant, anti-inflammatory, anti-hypercholesterolemia | [97] |
| Abelmoschus esculentus (ladies’ fingers or ochro) | Quercetin | Anti-inflammatory, antioxidant, hypolipidemic | [95] |
| Astragalus membranaceus (Mongolian milkvetch) | Total flavones | Inhibition of foam cell formation | [97] |
| Engelhardia roxburghiana (yellow basket-willow or roxburgh engelhartia) | Total flavonoids, naringenin, kaempferol, quercetin, isoengeletin, engeletin, astilbin, quercitrin | Decreased the serum lipids, downregulated NF-κB signaling | [98] |
| Scutellaria strigillosa Hemsley(namikiso (Japanese meaning: coming wave weed)) | Wogonin, wogonoside, baicalein, baicalin | Anti-proliferative and anti-migratory vascular smooth muscle cells | [99] |
| Garcinia madruno (madruno, charichuela, or madrono) | Pure biflavonoid aglycones morelloflavone, volkensiflavone | Protect low-density lipoprotein particle from both lipid and protein oxidation | [100] |
| Pandanus tectorius (pandan laut, tahitian screwpine, or pandanus) | Tangeretin | Anti-hypercholesterolemia | [101] |
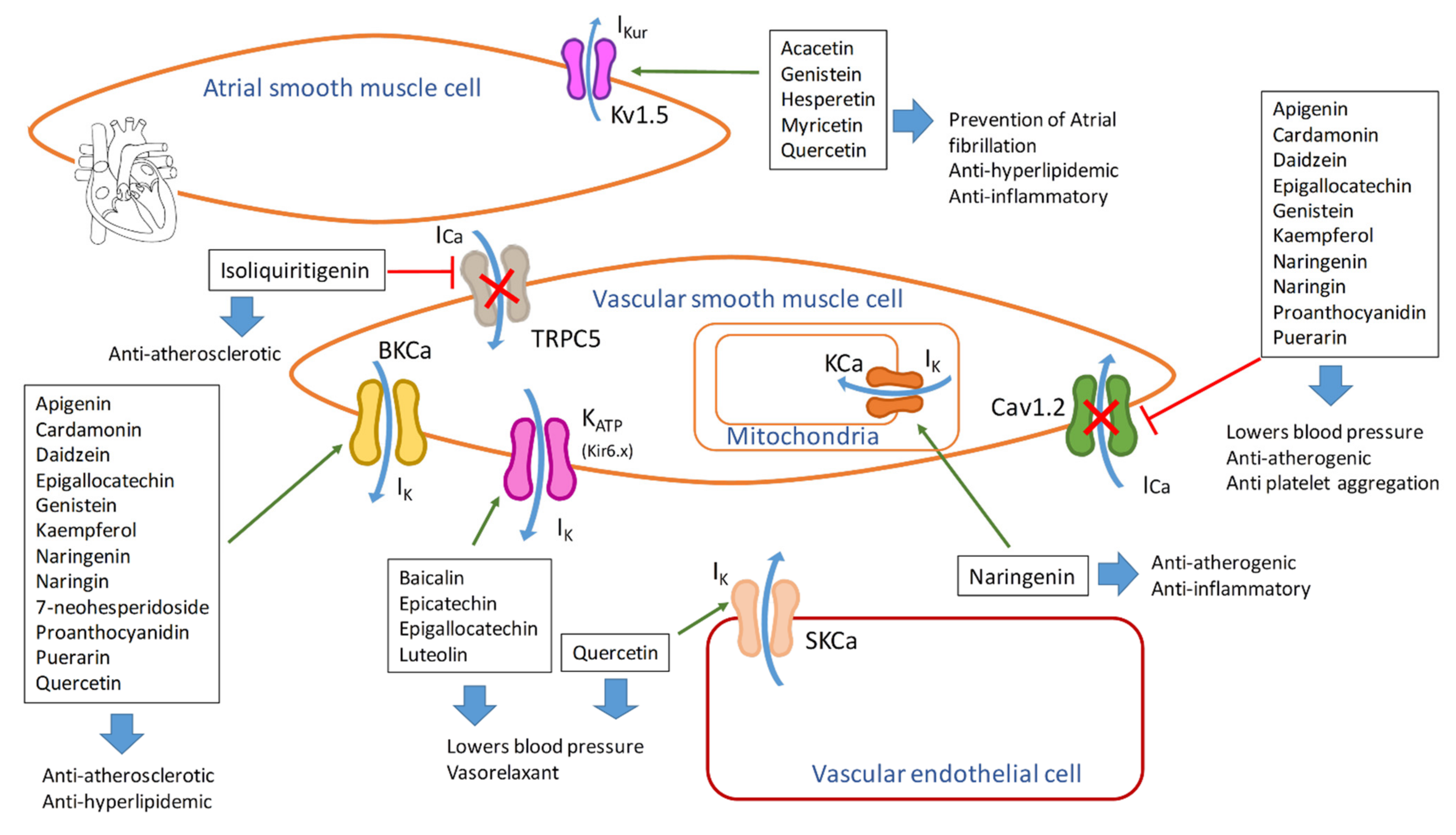
| Flavonoid | Ion Channel Target | Physiological Effect | Effect on Membrane Potential | Advantages in Atherosclerosis |
|---|---|---|---|---|
| Acacetin Apigenin trimethylether Genistein Hesperetin Myricetin Quercetin | Kv1.5 | Channel inhibitor | Decreased currents Inhibits ultrarapid delayed rectifier K+ currents (Ikur) | Prevention of atrial fibrillation Anti-hyperlipidemic, anti-inflammatory |
| Apigenin Cardamonin Daidzein Epigallocatechin Genistein Kaempferol Naringenin Naringin Proanthocyanidin Puerarin Quercetin | KCa1.1 (BKCa) | Channel activation Vasorelaxant | Increased currents, Hyperpolarization | Anti-atherosclerotic Anti-hyperlipidemic |
| Quercetin | SKCa | Channel activation Vasodilator effect | Increased currents, endothelial hyperpolarization, direct electrical coupling with VSMC hyperpolarization | Lowers blood pressure |
| Baicalin Epicatechin Epigallocatechin Luteolin | KATP | Channel activation Vasorelaxant | Increased currents Hyperpolarization | Lowers blood pressure |
| Epicatechin gallate Epigallocatechin gallate Genistein | Kir6.1 | Channel inhibitor | Decreased currents | Anti-inflammatory |
| Naringenin | Mitochondrial KCa | Vasodilator effect | Hyperpolarization | Anti-atherogenic, anti-inflammatory |
| Cardamonin Chrysin Daidzein Epigallocatechin gallate Galangin Genistein Hesperetin Kaempferol Morin Quercetin Scutellarein Tiliroside | Cav1.2 | Channel inhibitor Vasorelaxant | Decreased Ca currents | Lowers blood pressure Anti-atherogenic Anti-platelet aggregation |
| Isoliquiritigenin | TRPC5 | Channel inhibitor | Decreased Ca currents | Anti-atherosclerotic |
| Quercetin | NKCC1 (lung endothelial cells) | Channel activator | Increased Cl− currents | Lowers blood pressure by diminishing the expression of αENaC in renal cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grijalva-Guiza, R.E.; Jiménez-Garduño, A.M.; Hernández, L.R. Potential Benefits of Flavonoids on the Progression of Atherosclerosis by Their Effect on Vascular Smooth Muscle Excitability. Molecules 2021, 26, 3557. https://doi.org/10.3390/molecules26123557
Grijalva-Guiza RE, Jiménez-Garduño AM, Hernández LR. Potential Benefits of Flavonoids on the Progression of Atherosclerosis by Their Effect on Vascular Smooth Muscle Excitability. Molecules. 2021; 26(12):3557. https://doi.org/10.3390/molecules26123557
Chicago/Turabian StyleGrijalva-Guiza, Rosa Edith, Aura Matilde Jiménez-Garduño, and Luis Ricardo Hernández. 2021. "Potential Benefits of Flavonoids on the Progression of Atherosclerosis by Their Effect on Vascular Smooth Muscle Excitability" Molecules 26, no. 12: 3557. https://doi.org/10.3390/molecules26123557
APA StyleGrijalva-Guiza, R. E., Jiménez-Garduño, A. M., & Hernández, L. R. (2021). Potential Benefits of Flavonoids on the Progression of Atherosclerosis by Their Effect on Vascular Smooth Muscle Excitability. Molecules, 26(12), 3557. https://doi.org/10.3390/molecules26123557