Separation and Characterization of Phenolamines and Flavonoids from Rape Bee Pollen, and Comparison of Their Antioxidant Activities and Protective Effects Against Oxidative Stress

Abstract
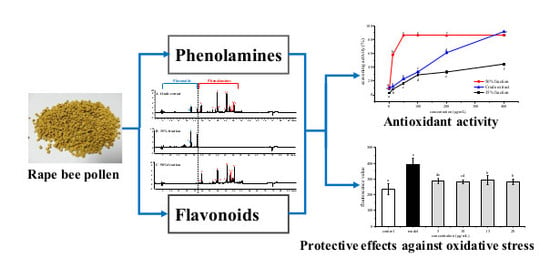
1. Introduction
2. Results
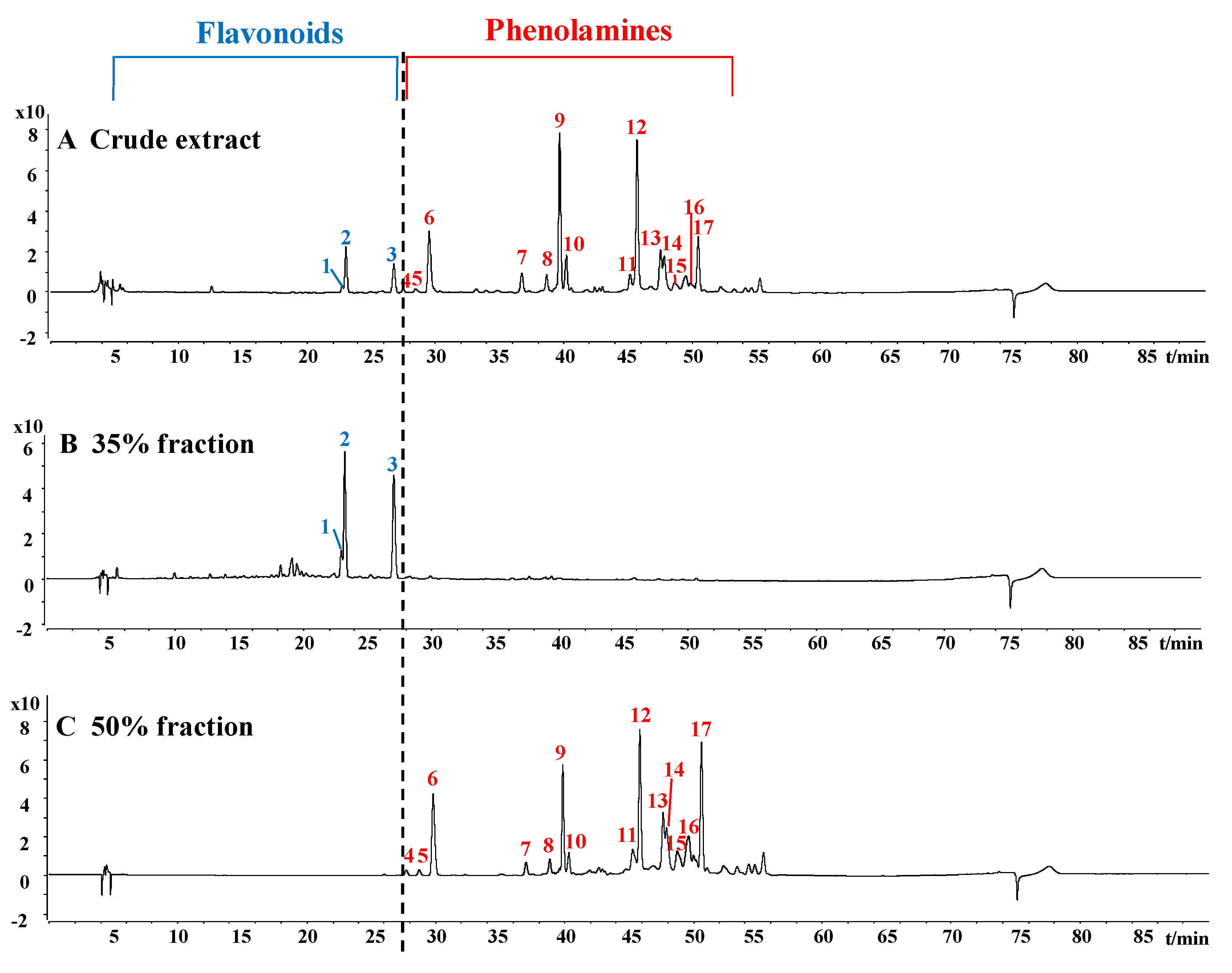
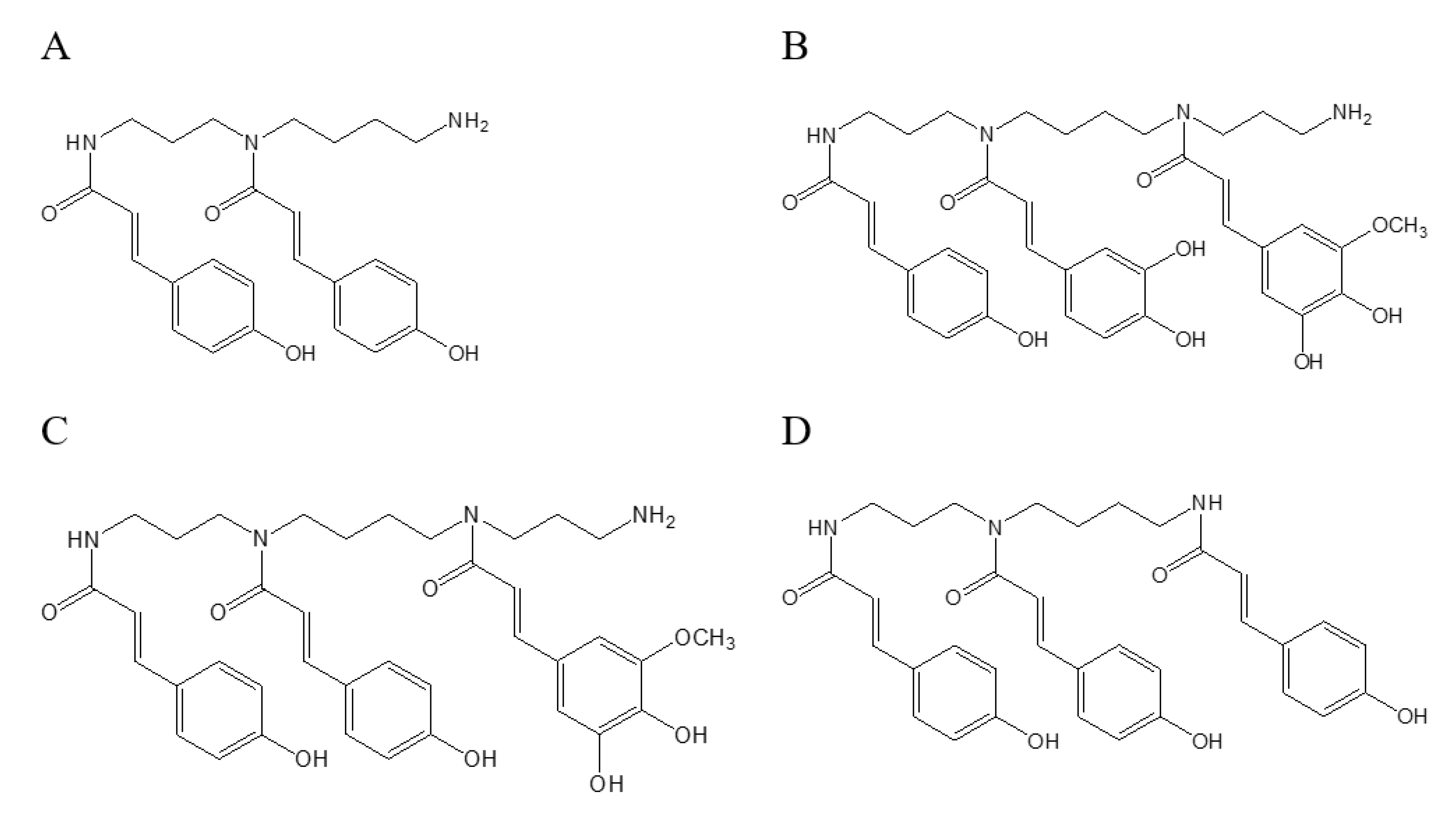
2.1. Characterization of Phenolamines and Flavonoids from Rape Bee Pollen
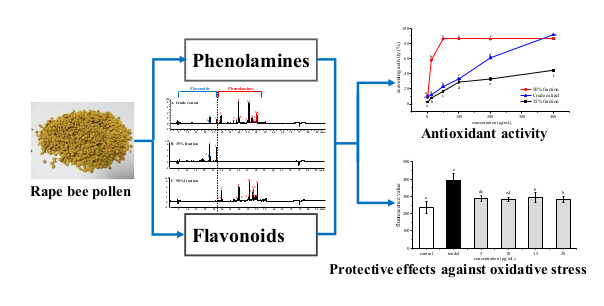
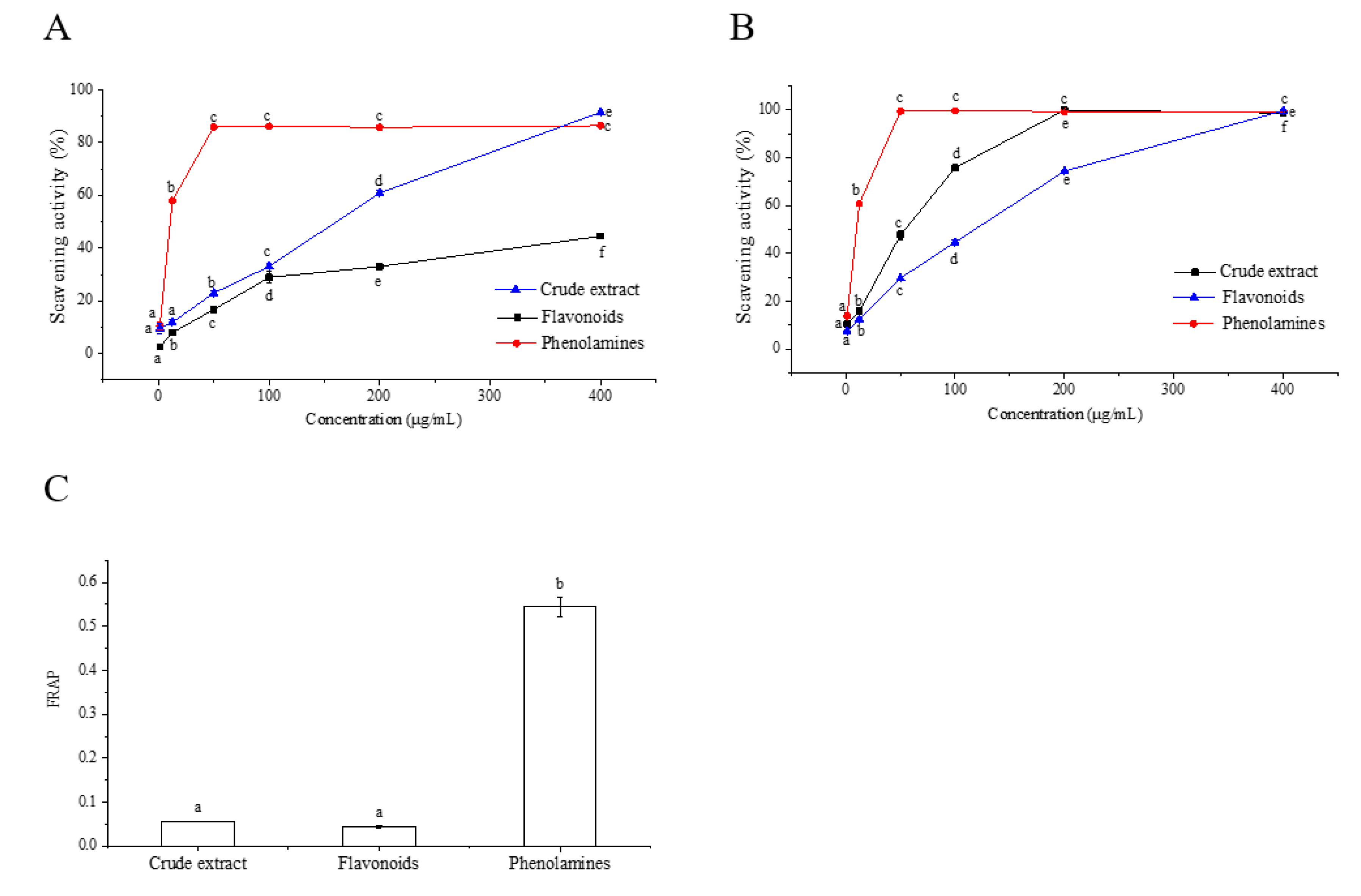
2.2. Antioxidant Activities of Phenolamines and Flavonoids
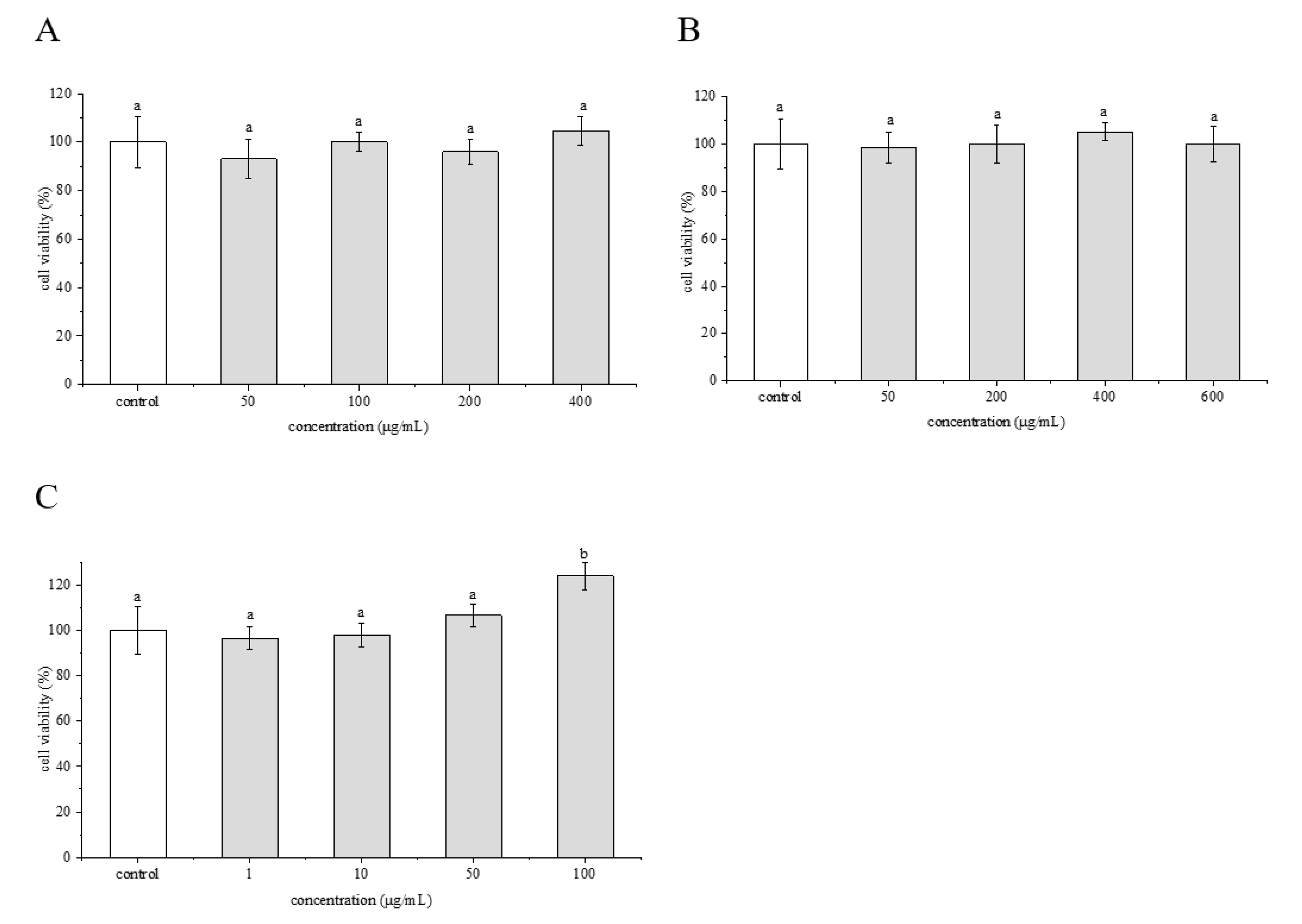
2.3. Cytotoxic Effects of Phenolamines and Flavonoids on HepG2 Cells
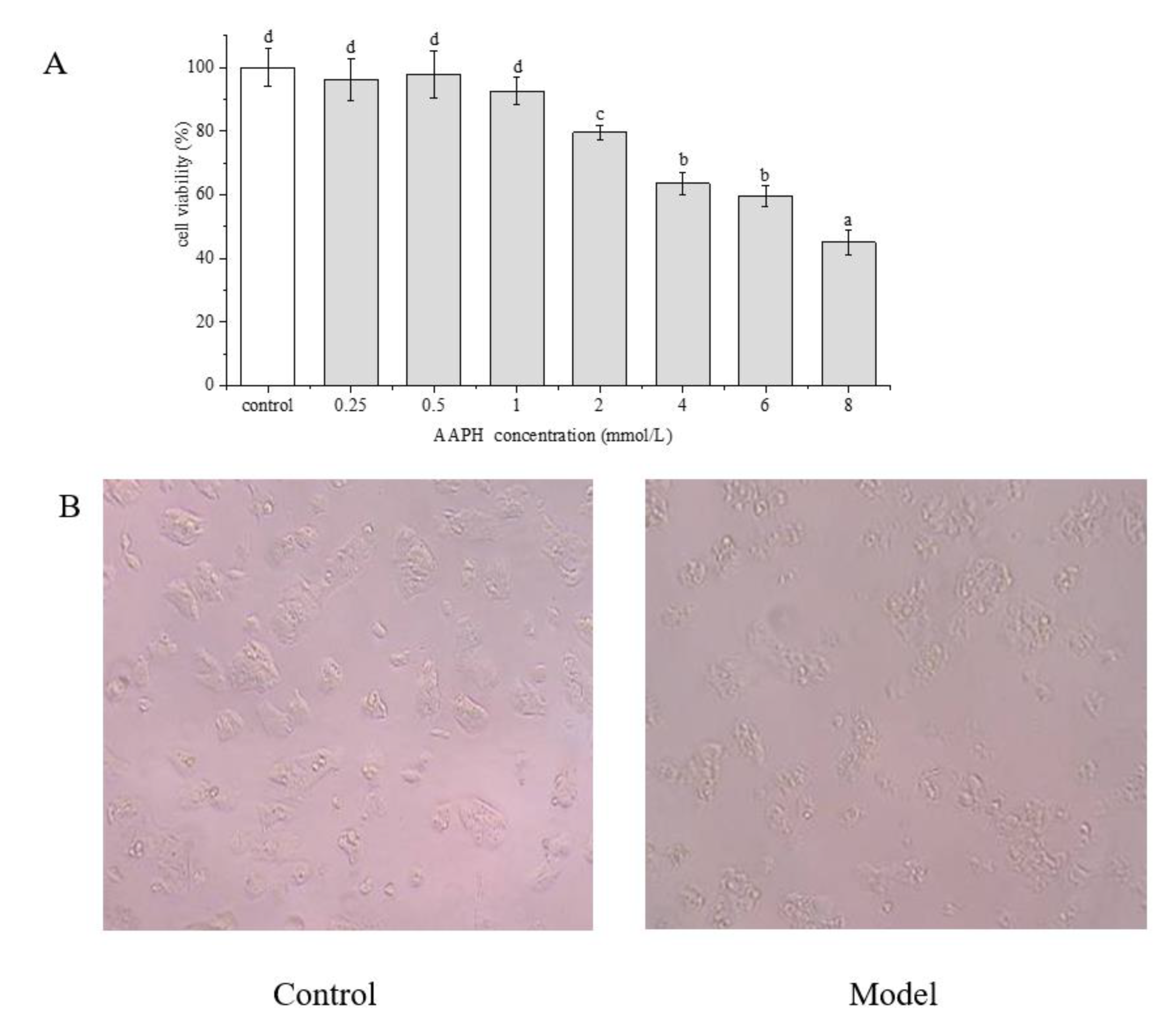
2.4. Effect of AAPH on HepG2 Cells
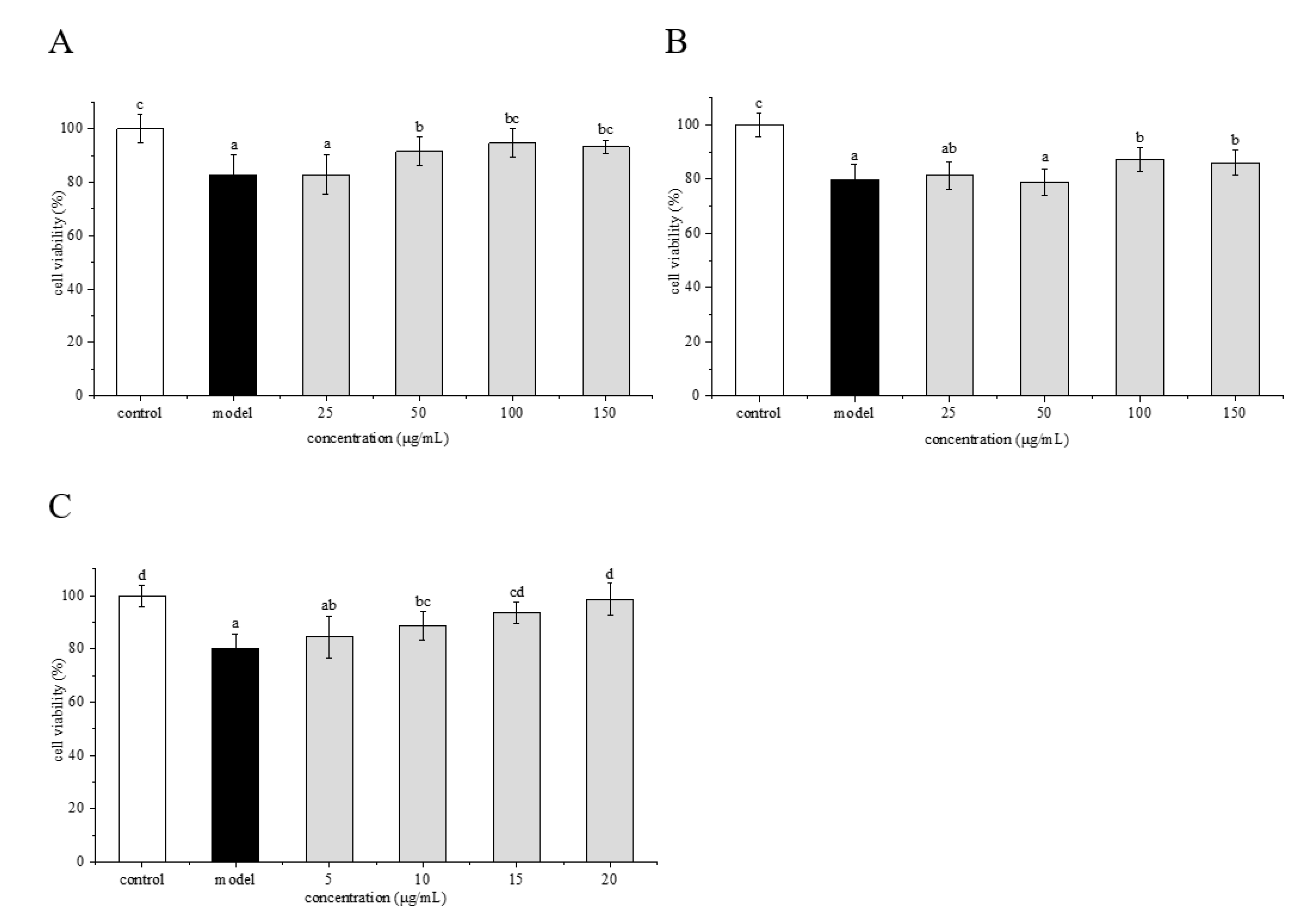
2.5. Protective Effects of Phenolamines and Flavonoids on HepG2 Cells Injured by AAPH
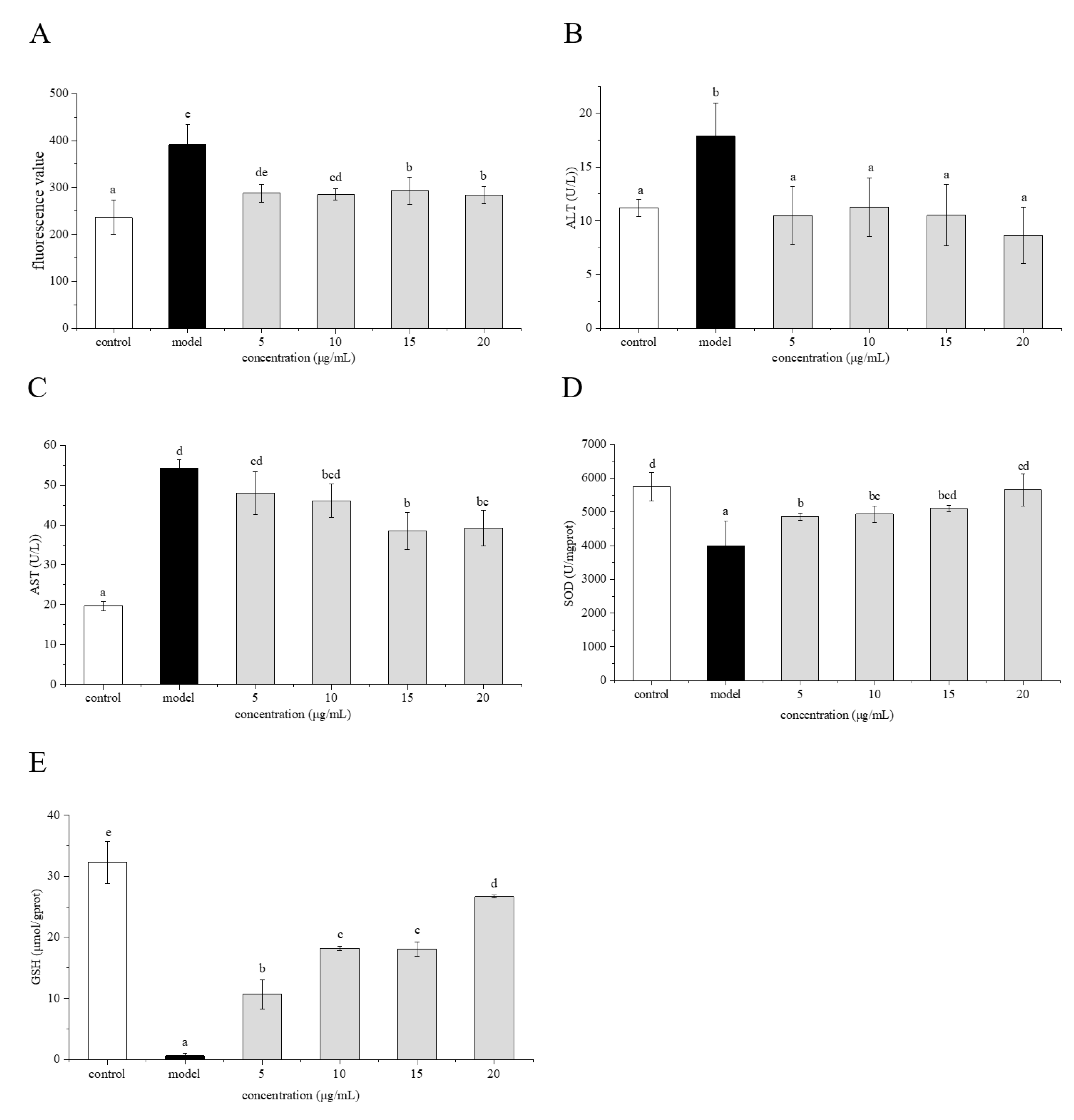
2.6. Preventive Effects of Phenolamines on ROS Generation
2.7. Effects of Phenolamines on AST and ALT Levels
2.8. Effects of Phenolamines on SOD Activity and GSH Content
3. Discussion
4. Materials and Methods
4.1. Materials and Chemical Reagents
4.2. Preparation of Rape Bee Pollen Crude Extract
4.3. Separation of Phenolamines and Flavonoids From Crude Extract
4.4. HPLC-ESI-QTOF-MS/MS Analysis
4.5. Antioxidant Activity
4.5.1. DPPH Radical Scavenging Activity Assay
4.5.2. ABTS Radical Scavenging Activity Assay
4.5.3. Ferric Reducing Antioxidant Power (FRAP) Assay
4.6. Cell Culture
4.7. Cytotoxic Effects of Crude Extract and Different Fractions on HepG2 Cells
4.8. Evaluation of Viability of HepG2 Cells Treated by AAPH
4.9. Assay of the Protective Effects of Crude Extract and Different Fractions on HepG2 Cells Injured by AAPH
4.10. Measurement of ROS
4.11. Determination of ALT and AST
4.12. Measurement of SOD and GSH
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Cao, H.; Chen, C.; Chen, X.; Wei, Q.; Zhao, F. Effects of fermentation by Ganoderma lucidum and Saccharomyces cerevisiae on rape pollen morphology and its wall. J. Food Sci. Technol. 2017, 54, 4026–4034. [Google Scholar] [CrossRef]
- Bassard, J.-E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef]
- Leja, M.; Mareczek, A.; Wyżgolik, G.; Klepacz-Baniak, J.; Czekońska, K. Antioxidative properties of bee pollen in selected plant species. Food Chem. 2007, 100, 237–240. [Google Scholar] [CrossRef]
- Morais, M.; Moreira, L.F.; Feás, X.; Estevinho, L.M.; Estevinho, L.M. Honeybee-collected pollen from five Portuguese Natural Parks: Palynological origin, phenolic content, antioxidant properties and antimicrobial activity. Food Chem. Toxicol. 2011, 49, 1096–1101. [Google Scholar] [CrossRef]
- Chen, X.; Wu, R.-Z.; Zhu, Y.; Ren, Z.-M.; Tong, Y.-L.; Yang, F.; Dai, G. Study on the inhibition of Mfn1 by plant-derived miR5338 mediating the treatment of BPH with rape bee pollen. BMC Complement. Altern. Med. 2018, 18, 38. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Śmigiel, A.C.; Wolska, I. Pollen and bee bread as new health-oriented products: A review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Pascoal, A.; Rodrigues, S.; Teixeira, A.; Feás, X.; Estevinho, L.M.; Estevinho, L.M. Biological activities of commercial bee pollens: Antimicrobial, antimutagenic, antioxidant and anti-inflammatory. Food Chem. Toxicol. 2014, 63, 233–239. [Google Scholar] [CrossRef]
- Sun, L.; Guo, Y.; Zhang, Y.; Zhuang, Y. Antioxidant and Anti-tyrosinase Activities of Phenolic Extracts from Rape Bee Pollen and Inhibitory Melanogenesis by cAMP/MITF/TYR Pathway in B16 Mouse Melanoma Cells. Front. Pharmacol. 2017. [Google Scholar] [CrossRef]
- Ariza, M.T.; Forbes-Hernández, T.; Rodríguez, P.R.; Afrin, S.; Gasparrini, M.; Cervantes, L.; Soria, C.; Martínez-Ferri, E.; Battino, M.; Giampieri, F. Strawberry and Achenes Hydroalcoholic Extracts and Their Digested Fractions Efficiently Counteract the AAPH-Induced Oxidative Damage in HepG2 Cells. Int. J. Mol. Sci. 2018, 19, 2180. [Google Scholar] [CrossRef]
- Joko, S.; Watanabe, M.; Fuda, H.; Takeda, S.; Furukawa, T.; Hui, S.-P.; Shrestha, R.; Chiba, H. Comparison of chemical structures and cytoprotection abilities between direct and indirect antioxidants. J. Funct. Foods 2017, 35, 245–255. [Google Scholar] [CrossRef]
- Wu, J.H.; Huo, J.Y.; Huang, M.Q.; Zhao, M.M.; Luo, X.L.; Sun, B.G. Structural characterization of a tetrapeptide from sesame flavor-type Baijiu and its preventive effects against AAPH-iInduced oxidative stress in HepG2 cells. J. Agr. Food Chem. 2017, 65, 10495–10504. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Chuang, C.-C.; Wu, S.; Zuo, L. Reactive oxygen species in redox cancer therapy. Cancer Lett. 2015, 367, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.; Liu, Y.; Sun, B. Wheat bran feruloyl oligosaccharides modulate the phase II detoxifying/antioxidant enzymes via Nrf2 signaling. Int. J. Boil. Macromol. 2015, 74, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Cha, S.-H.; Wijesinghe, W.A.J.P.; Kang, S.-M.; Lee, S.-H.; Kim, E.-A.; Song, C.B.; Jeon, Y.-J. Protective effect of marine algae phlorotannins against AAPH-induced oxidative stress in zebrafish embryo. Food Chem. 2013, 138, 950–955. [Google Scholar] [CrossRef]
- Kim, E.-A.; Lee, S.-H.; Ko, C.-I.; Cha, S.-H.; Kang, M.-C.; Kang, S.-M.; Ko, S.-C.; Lee, W.-W.; Ko, J.-Y.; Lee, J.-H.; et al. Protective effect of fucoidan against AAPH-induced oxidative stress in zebrafish model. Carbohydr. Polym. 2014, 102, 185–191. [Google Scholar] [CrossRef]
- Mărghitaş, L.A.; Stanciu, O.G.; Dezmirean, D.S.; Bobis, O.; Popescu, O.; Bogdanov, S.; Campos, M.D.G.R. In vitro antioxidant capacity of honeybee-collected pollen of selected floral origin harvested from Romania. Food Chem. 2009, 115, 878–883. [Google Scholar] [CrossRef]
- Gacche, R.; Shegokar, H.D.; Gond, D.S.; Yang, Z.; Jadhav, A.D. Evaluation of Selected Flavonoids as Antiangiogenic, Anticancer, and Radical Scavenging Agents: An Experimental and In Silico Analysis. Cell Biophys. 2011, 61, 651–663. [Google Scholar] [CrossRef]
- Sousa, C.; Andrade, P.B.; Valentão, P. Relationships of Echium plantagineum L. bee pollen, dietary flavonoids and their colonic metabolites with cytochrome P450 enzymes and oxidative stress. RSC Adv. 2016, 6, 6084–6092. [Google Scholar] [CrossRef]
- Negri, G.; Teixeira Érica, W.; Alves, M.L.T.M.F.; Moreti, A.C.D.C.C.; Otsuk, I.P.; Borguini, R.G.; Salatino, A. Hydroxycinnamic Acid Amide Derivatives, Phenolic Compounds and Antioxidant Activities of Extracts of Pollen Samples from Southeast Brazil. J. Agric. Food Chem. 2011, 59, 5516–5522. [Google Scholar] [CrossRef]
- Pihlava, J.-M. Identification of hordatines and other phenolamides in barley (Hordeum vulgare) and beer by UPLC-QTOF-MS. J. Cereal Sci. 2014, 60, 645–652. [Google Scholar] [CrossRef]
- Lv, H.; Wang, X.; He, Y.; Wang, H.; Suo, Y. Identification and quantification of flavonoid aglycones in rape bee pollen from Qinghai-Tibetan Plateau by HPLC-DAD-APCI/MS. J. Food Compos. Anal. 2015, 38, 49–54. [Google Scholar] [CrossRef]
- Wang, R.-D.; Su, G.-H.; Wang, L.; Xia, Q.; Liu, R.; Lu, Q.; Zhang, J.-L. Identification and mechanism of effective components from rape ( Brassica napus L.) bee pollen on serum uric acid level and xanthine oxidase activity. J. Funct. Foods 2018, 47, 241–251. [Google Scholar] [CrossRef]
- Mihajlovic, L.; Radosavljević, J.; Burazer, L.; Smiljanic, K.; Velickovic, T.C. Composition of polyphenol and polyamide compounds in common ragweed (Ambrosia artemisiifolia L.) pollen and sub-pollen particles. Phytochem. 2015, 109, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Chen, X.; Netrusov, A.I.; Zhou, Q.; Guo, D.; Liu, X.; He, H.; Xin, X.; Wang, Y.; Chen, L. Screening and Identifying Antioxidative Components in Ginkgo biloba Pollen by DPPH-HPLC-PAD Coupled with HPLC-ESI-MS2. PloS ONE 2017. [Google Scholar] [CrossRef] [PubMed]
- Handrick, V.; Vogt, T.; Frolov, A. Profiling of hydroxycinnamic acid amides in Arabidopsis thaliana pollen by tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2789–2801. [Google Scholar] [CrossRef]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef]
- Yang, K.; Wu, D.; Ye, X.; Liu, D.; Chen, J.; Sun, P. Characterization of Chemical Composition of Bee Pollen in China. J. Agric. Food Chem. 2013, 61, 708–718. [Google Scholar] [CrossRef]
- Kite, G.C.; Larsson, S.; Veitch, N.C.; Porter, E.A.; Liang, X.; Simmonds, M.S.J. Acyl Spermidines in Inflorescence Extracts of Elder (Sambucus nigra L., Adoxaceae) and Elderflower Drinks. J. Agric. Food Chem. 2013, 61, 3501–3508. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Sun, J.; Chen, P.; Harnly, J. UHPLC-PDA-ESI/HRMS/MSnAnalysis of Anthocyanins, Flavonol Glycosides, and Hydroxycinnamic Acid Derivatives in Red Mustard Greens (Brassica juncea Coss Variety). J. Agric. Food Chem. 2011, 59, 12059–12072. [Google Scholar] [CrossRef]
- Sobolev, V.S.; Sy, A.A.; Gloer, J.B. Spermidine and Flavonoid Conjugates from Peanut (Arachis hypogaea) Flowers. J. Agric. Food Chem. 2008, 56, 2960–2969. [Google Scholar] [CrossRef]
- Sun, J.; Song, Y.-L.; Zhang, J.; Huang, Z.; Huo, H.-X.; Zheng, J.; Zhang, Q.; Zhao, Y.-F.; Li, J.; Tu, P. Characterization and Quantitative Analysis of Phenylpropanoid Amides in Eggplant (Solanum melongenaL.) by High Performance Liquid Chromatography Coupled with Diode Array Detection and Hybrid Ion Trap Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2015, 63, 3426–3436. [Google Scholar] [CrossRef] [PubMed]
- Wiese, S.; Wubshet, S.G.; Nielsen, J.; Staerk, D. Coupling HPLC-SPE-NMR with a microplate-based high-resolution antioxidant assay for efficient analysis of antioxidants in food – Validation and proof-of-concept study with caper buds. Food Chem. 2013, 141, 4010–4018. [Google Scholar] [CrossRef] [PubMed]
- Dudonneé, S.; Vitrac, X.; Coutieère, P.; Woillez, M.; Meérillon, J.-M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Benhar, M. Roles of mammalian glutathione peroxidase and thioredoxin reductase enzymes in the cellular response to nitrosative stress. Free. Radic. Boil. Med. 2018, 127, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Kalaycıoğlu, Z.; Kaygusuz, H.; Döker, S.; Kolaylı, S.; Erim, F.B. Characterization of Turkish honeybee pollens by principal component analysis based on their individual organic acids, sugars, minerals, and antioxidant activities. LWT 2017, 84, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | [M–H]–(m/z) | MS2 ion Fragments (m/z) | Proposed Formula | Error (ppm) | Tentative Identification | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 23.169 | 625.1402 | 301.0356, 463.0888 | C27H30O17 | 0.86 | Quercetin di-glucoside | 22,23 |
| 2 | 23.422 | 609.1450 | 283.0253, 285.0409, 446.0856 | C27H30O16 | 1.12 | kaempferol di-glucoside | 24 |
| 3 | 27.328 | 609.1460 | 255.0308, 284.0329, 285.0395, 429.0833 | C27H30O16 | 0.14 | kaempferol di-glucoside | 24 |
| Peak | RT (min) | [M–H]–(m/z) | MS2 ion Fragments (m/z) | Proposed Formula | Error (ppm) | Tentative Identification | Reference |
|---|---|---|---|---|---|---|---|
| 4 | 28.055 | 436.2247 | 119.0500, 316.1677 | C25H31N3O4 | 1.24 | di-p-coumaroyl spermidine | 22 |
| 5 | 29.079 | 436.2241 | 119.0505, 316.1673 | C25H31N3O4 | 0.08 | di-p-coumaroyl spermidine | 22 |
| 6 | 29.882 | 436.2245 | 119.0498, 316.1669 | C25H31N3O4 | 0.42 | di-p-coumaroyl spermidine | 22 |
| 7 | 37.234 | 701.3173 | 135.0452, 165.0560, 399.2043, 535.2560, 555.2834, 565.2671 | C38H46N4O9 | 2.53 | p-coumaroyl caffeoyl hydroxyferuloyl spermine | * |
| 8 | 39.080 | 685.3245 | 119.0503, 145.0284, 165.0557, 399.2039, 519.2608 | C38H46N4O8 | 0.08 | di-p-coumaroyl hydroxyferuloyl spermine | * |
| 9 | 40.087 | 685.3247 | 119.0499, 145.0287, 165.0557, 399.2045, 519.2619 | C38H46N4O8 | 0.43 | di-p-coumaroyl hydroxyferuloyl spermine | * |
| 10 | 40.576 | 685.3190 | 119.0505, 145.0279, 165.0534, 399.2058, 519.2603 | C38H46N4O8 | 1.68 | di-p-coumaroyl hydroxyferuloyl spermine | * |
| 11 | 45.453 | 439.1684 | 135.0447, 161.0243, 165.0556, 415.2000 | C16H24N8O7 | 1.39 | unknown | |
| 12 | 46.060 | 439.1667 | 135.0448, 161.0241, 165.0553, 415.1984 | C16H24N8O7 | 1.60 | unknown | |
| 13 | 47.839 | 582.2604 | 119.0491, 145.0291, 342.1464, 462.2025 | C34H37N3O6 | 0.90 | tri-p-coumaroyl spermidine | 23 |
| 14 | 48.948 | 582.2599 | 119.0498, 145.0302, 342.1457, 462.2035 | C34H37N3O6 | 1.62 | tri-p-coumaroyl spermidine | 23 |
| 15 | 49.033 | 582.2607 | 119.0501, 145.0296, 342.1463, 462.2027 | C34H37N3O6 | 0.54 | tri-p-coumaroyl spermidine | 23 |
| 16 | 49.790 | 582.2602 | 119.0501, 145.0294, 342.1464, 462.2036 | C34H37N3O6 | 1.28 | tri-p-coumaroyl spermidine | 23 |
| 17 | 50.835 | 582.2602 | 119.0494, 145.0286, 342.1459, 462.2032 | C34H37N3O6 | 1.21 | tri-p-coumaroyl spermidine | 23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, R.; Lu, Q. Separation and Characterization of Phenolamines and Flavonoids from Rape Bee Pollen, and Comparison of Their Antioxidant Activities and Protective Effects Against Oxidative Stress. Molecules 2020, 25, 1264. https://doi.org/10.3390/molecules25061264
Zhang H, Liu R, Lu Q. Separation and Characterization of Phenolamines and Flavonoids from Rape Bee Pollen, and Comparison of Their Antioxidant Activities and Protective Effects Against Oxidative Stress. Molecules. 2020; 25(6):1264. https://doi.org/10.3390/molecules25061264
Chicago/Turabian StyleZhang, Huifang, Rui Liu, and Qun Lu. 2020. "Separation and Characterization of Phenolamines and Flavonoids from Rape Bee Pollen, and Comparison of Their Antioxidant Activities and Protective Effects Against Oxidative Stress" Molecules 25, no. 6: 1264. https://doi.org/10.3390/molecules25061264
APA StyleZhang, H., Liu, R., & Lu, Q. (2020). Separation and Characterization of Phenolamines and Flavonoids from Rape Bee Pollen, and Comparison of Their Antioxidant Activities and Protective Effects Against Oxidative Stress. Molecules, 25(6), 1264. https://doi.org/10.3390/molecules25061264