Antioxidant, α-Glucosidase, and Nitric Oxide Inhibitory Activities of Six Algerian Traditional Medicinal Plant Extracts and 1H-NMR-Based Metabolomics Study of the Active Extract

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
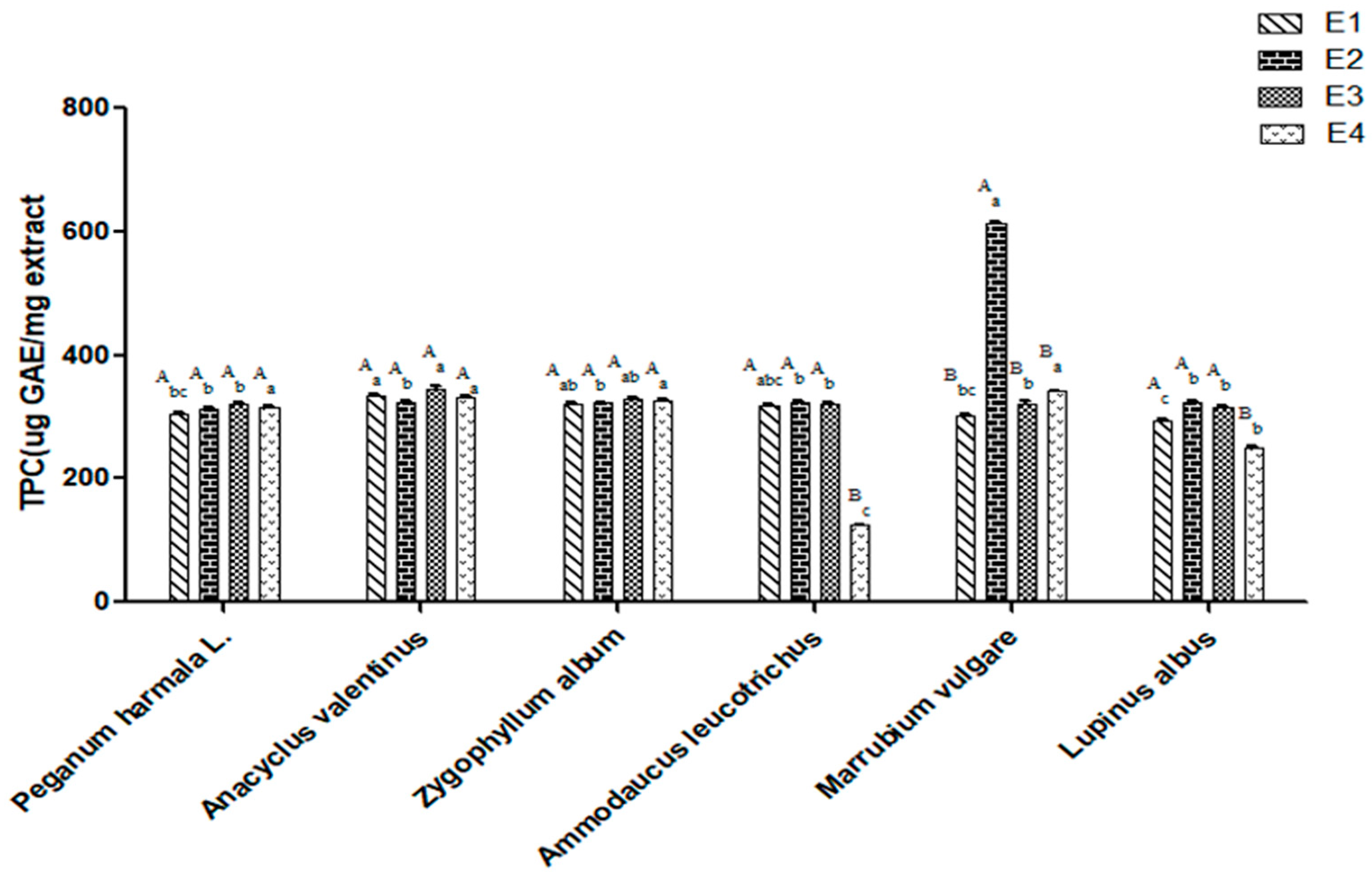
2.1. Effect of Ethanol Ratios on the Total Phenolic Content of Selected Plant Extracts
2.2. Effect of Ethanol Ratios on the Free Radical Scavenging Activity
2.3. Effect of Ethanol Ratios on the α-Glucosidase Inhibitory Activity
2.4. Effect of Ethanol Ratios on the Nitric Oxide Inhibitory Activity
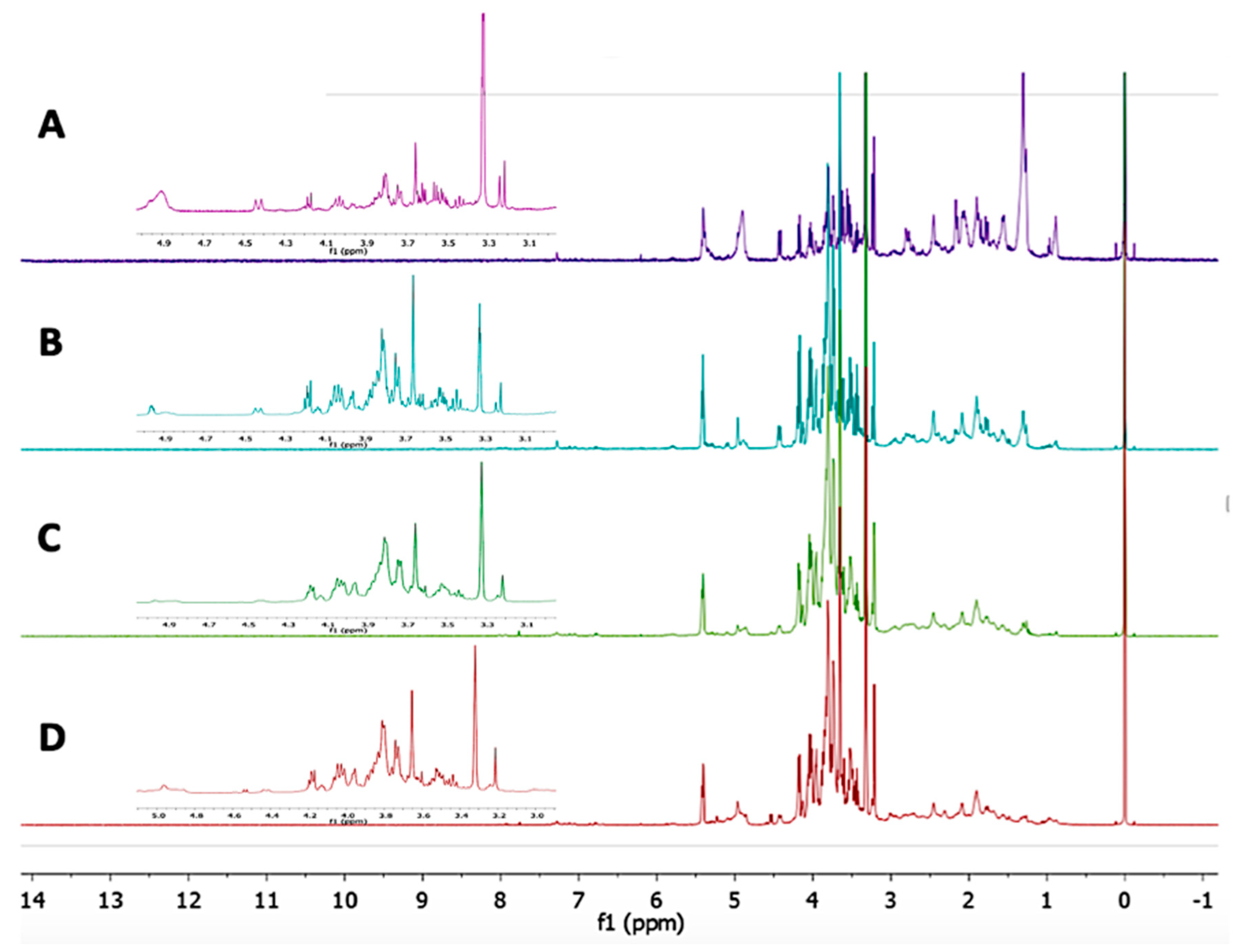
2.5. Identification of the Metabolites from L. Albus Extract
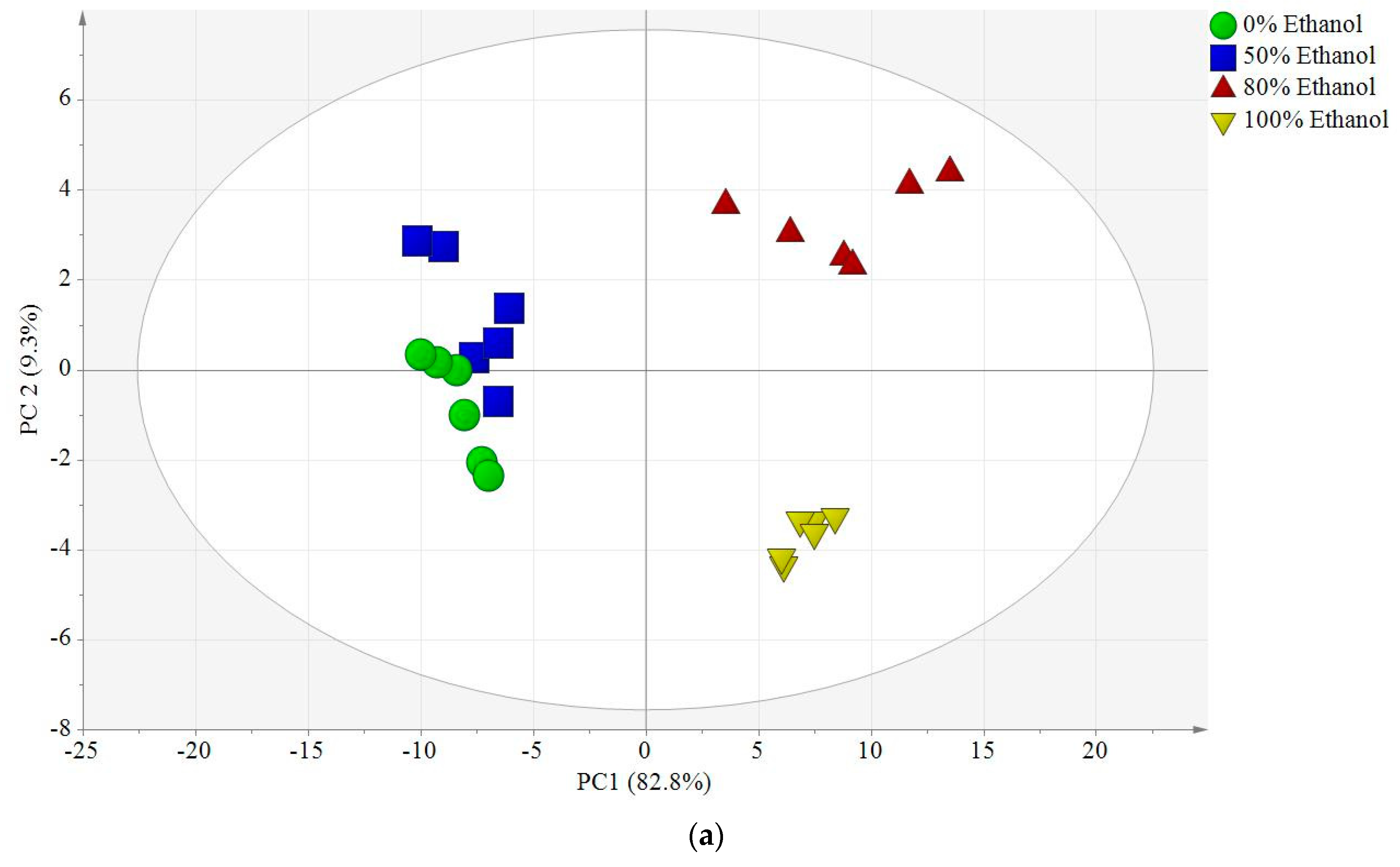
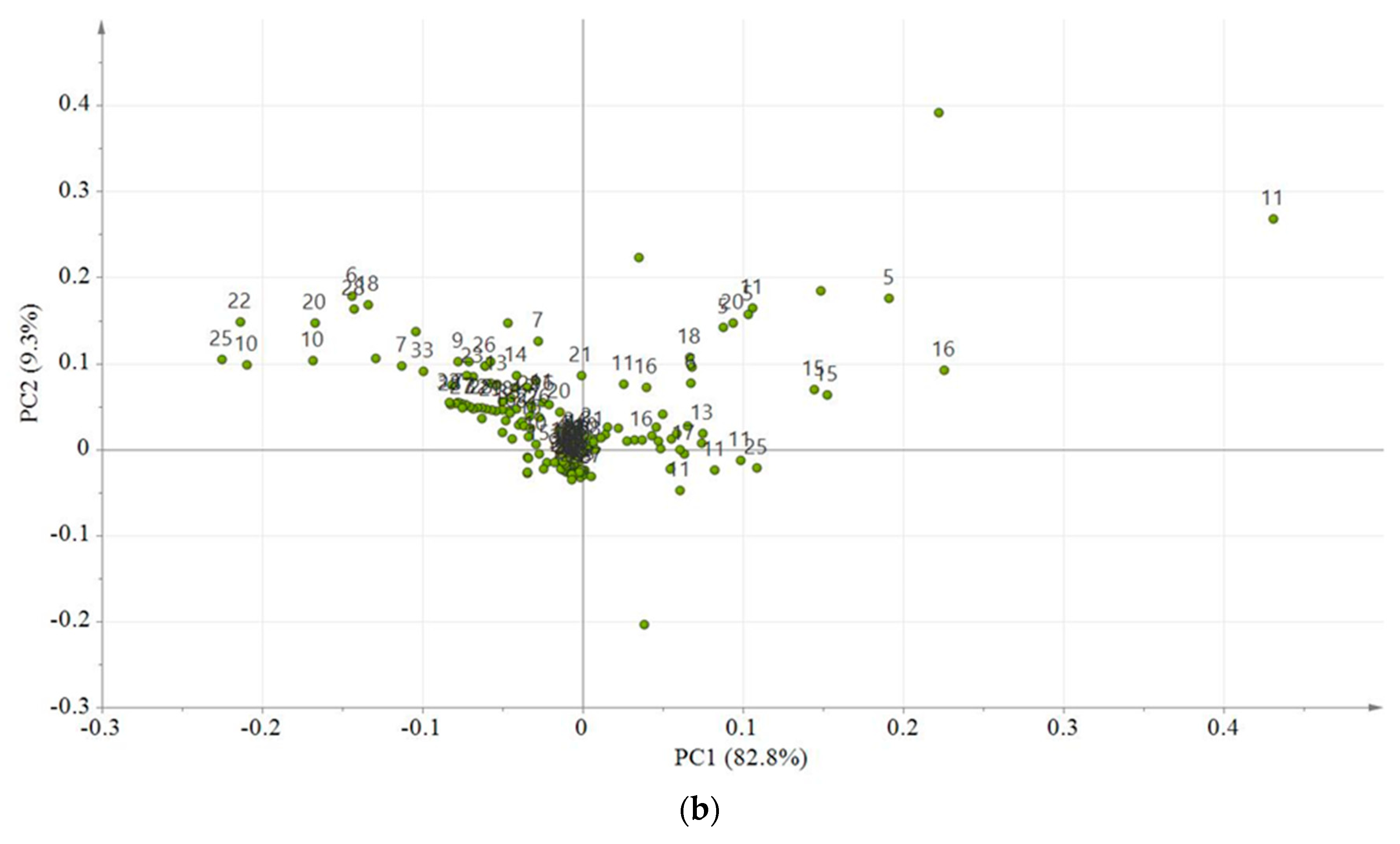
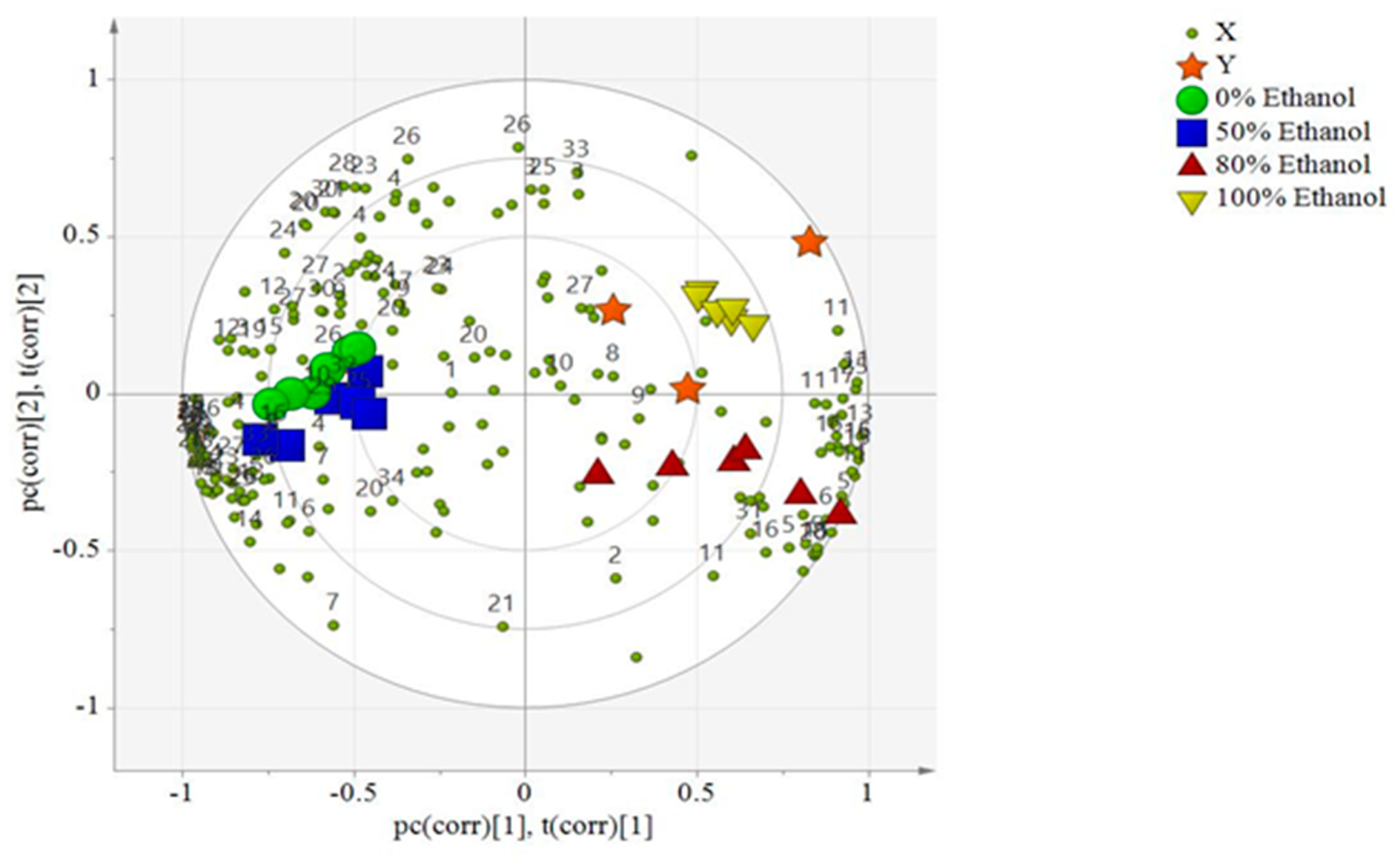
2.6. Multivariate Data Analyses
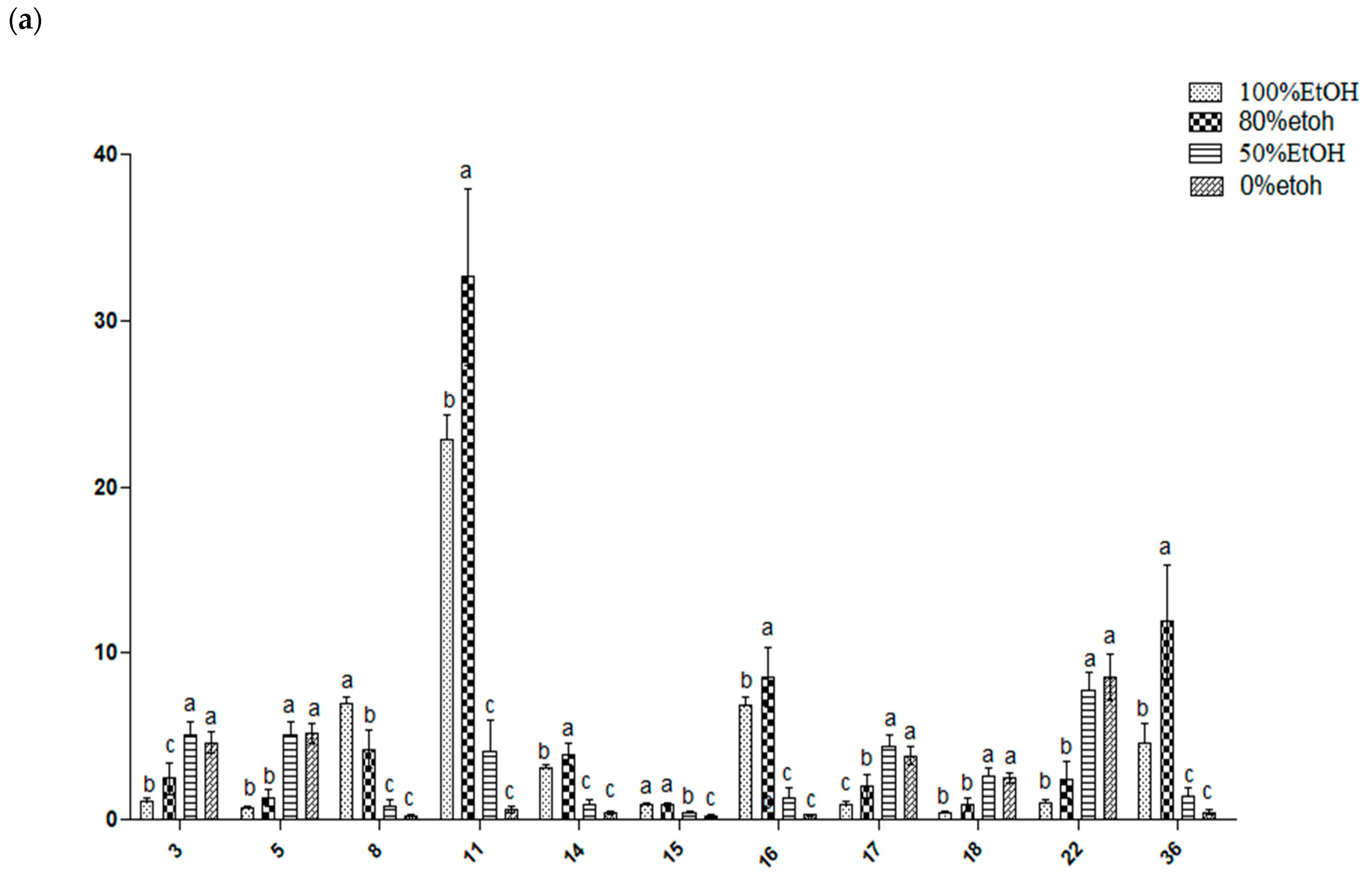
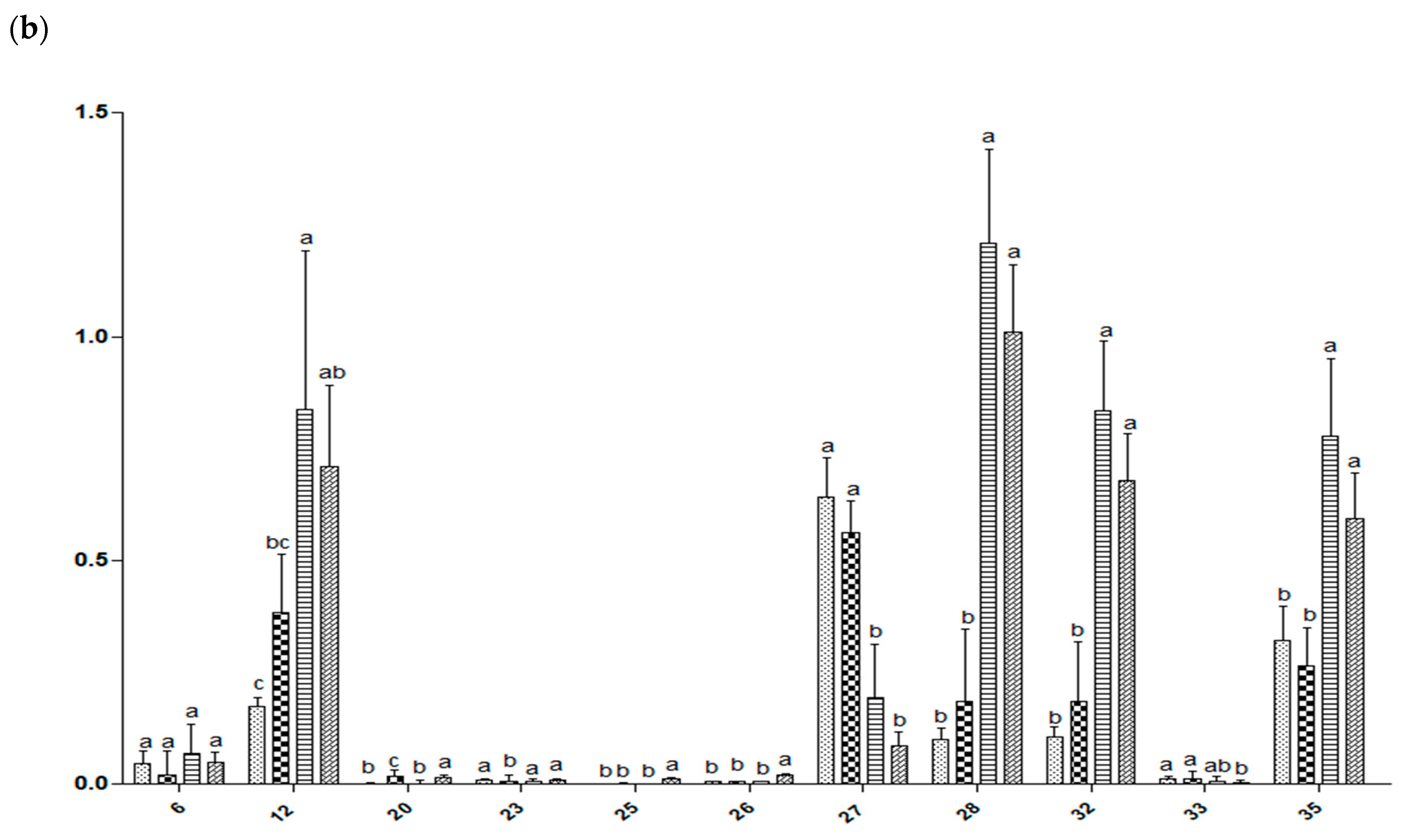
2.7. Relative Quantification of Metabolites
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Materials
3.3. Plant Extraction
3.4. Total Phenolic Content (TPC)
3.5. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Assay
3.6. α-Glucosidase Inhibitory Activity
3.7. Nitric Oxide Inhibitory Activity
3.8. Nuclear Magnetic Resonance Measurement
3.9. Multivariate Data Analysis
3.10. Statistics Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Patel, P.; Harde, P.; Pillai, J.; Darji, N.; Patel, B. Antidiabetic herbal drugs a review. Pharmacophore 2012, 3, 18–29. [Google Scholar]
- Ziani, B.E.; Calhelha, R.C.; Barreira, J.C.; Barros, L.; Hazzit, M.; Ferreira, I.C. Bioactive properties of medicinal plants from the Algerian flora: Selecting the species with the highest potential in view of application purposes. Ind. Crop Prod. 2015, 77, 582–589. [Google Scholar] [CrossRef]
- Ababou, A.; Chouieb, M.; Bouthiba, A.; Saidi, D.; Bouzina, M.M.; Mederbal, K. Spatial pattern analysis of Peganum harmala on the salted lower Chelif plain, Algeria. Turk. J. Botany 2013, 37, 111–121. [Google Scholar]
- Monsef, H.R.; Ghobadi, A.; Iranshahi, M.; Abdollahi, M. Antinociceptive effects of Peganum harmala L. alkaloid extract on mouse formalin test. J. Pharm. Pharm. Sci. 2004, 7, 65–69. [Google Scholar] [PubMed]
- Zaker, F.; Oody, A.; Arjmand, A. A study on the antitumoral and differentiation effects of Peganum harmala derivatives in combination with ATRA on leukaemic cells. Arch. Pharm. Res. 2007, 30, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Chaturvedi, J.P.; Narender, T.; Srivastava, A.K. Preliminary studies on the hypoglycemic effect of Peganum harmala L. seeds ethanol extract on normal and streptozotocin induced diabetic rats. Indian J. Clin. Biochem. 2008, 23, 391–393. [Google Scholar] [CrossRef][Green Version]
- El Ghoul, J.; Ghanem-Boughanmi, N.; Ben-Attia, M. Biochemical study on the protective effect of ethanolic extract of Zygophyllum album on streptozotocin-induced oxidative stress and toxicity in mice. Biomed. Prev. Nutr. 2011, 1, 79–83. [Google Scholar] [CrossRef]
- Mnafgui, K.; Kchaou, M.; Ben Salah, H.; Hajji, R.; Khabbabi, G.; Elfeki, A.; Allouche, N.; Gharsallah, N. Essential oil of Zygophyllum album inhibits key-digestive enzymes related to diabetes and hypertension and attenuates symptoms of diarrhea in alloxan-induced diabetic rats. Pharm. Biol. 2016, 54, 1326–1333. [Google Scholar] [CrossRef]
- Tadjeddine, A.L.; Kambouche, N.; Medjdoub, H.; Meddah, B.; Dicko, A.; Saidi, S.; Derdour, A. Antidiabetic effect of Anacyclus valentinus L. aqueous extract in normoglycemic and streptozotocin induced-diabetic rats. Am. J. Phytomed. Clin. Ther. 2013, 1, 424–431. [Google Scholar]
- Magnan, C.; Ktorza, A. Production et sécrétion de l’insuline par la cellule β pancréatique. EMC-Endocrinologie 2005, 2, 241–264. [Google Scholar] [CrossRef]
- Muckensturm, B.; Diyani, F.; Le Nouën, D.; Fkih-Tetouani, S.; Reduron, J.P. Ammolactone, a guaianolide from a medicinal plant, Ammodaucus leucotrichus. Phytochemistry 1997, 44, 907–910. [Google Scholar] [CrossRef]
- Román, R.R.; Alarcón-Aguilar, F.R.; Lara-Lemus, A.L.; Flores-Saenz, J.L. Hypoglycemic effect of plants used in Mexico as antidiabetics. Arch. Med. Res. 1992, 23, 59–64. [Google Scholar]
- Sahpaz, S.; Garbacki, N.; Tits, M.; Bailleul, F. Isolation and pharmacological activity of phenylpropanoid esters from Marrubium vulgare. J. Ethnopharmacol. 2002, 79, 389–392. [Google Scholar] [CrossRef]
- Amer, M.M. Constituents of the aerial parts of Marrubium vulgare L. J. Pharm Sci. 1993, 9, 92–98. [Google Scholar]
- Grieve, M. A Modern Herbal; Courier Corporation: North Chelmsford, MA, USA, 1971; Volume 1, p. 514. [Google Scholar]
- Huyghe, C. White lupin (Lupinus albus L.). Field Crops Res. 1997, 53, 147–160. [Google Scholar] [CrossRef]
- Villano, D.; Fernández-Pachón, M.S.; Moyá, M.L.; Troncoso, A.M.; García-Parrilla, M.C. Radical scavenging ability of polyphenolic compounds towards DPPH free radical. Talanta 2007, 71, 230–235. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Bioch. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- Mohsen, S.M.; Ammar, A.S. Total phenolic contents and antioxidant activity of corn tassel extracts. Food Chem. 2009, 112, 595–598. [Google Scholar] [CrossRef]
- Stankovic, M.S. Total phenolic content, flavonoid concentration and antioxidant activity of Marrubium peregrinum L. extracts. Kragujevac J. Sci. 2011, 33, 63–72. [Google Scholar]
- Amri, B.; Martino, E.; Vitulo, F.; Corana, F.; Kaâb, L.B.; Rui, M.; Rossi, D.; Mori, M.; Rossi, S.; Collina, S. Marrubium vulgare L. Leave extract: Phytochemical composition, antioxidant and wound healing properties. Molecules 2017, 22, 1851. [Google Scholar] [CrossRef]
- Miguel, M.; Bouchamaa, N.; Aazza, S.; Gaamoussi, F.; Lyoussi, B. Antioxidant, anti-inflammatory and anti-acetylcholinesterase activities of eleven extracts of Moroccan plants. Fresen. Environ. Bull. 2014, 23, 1–4. [Google Scholar]
- McDermott, J.H. Antioxidant nutrients: Current dietary recommendations and research update. J. Am. Pharm. Assoc. (1996) 2000, 40, 785–799. [Google Scholar]
- Namiki, M. Antioxidants/antimutagens in food. Crit. Rev. Food Sci. Nutr. 1990, 29, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Sebaa, A.; Marouf, A.; Kambouche, N.; Derdour, A. Phytochemical composition, antioxidant and antimicrobial activities of Ammodaucus leucotrichus fruit from Algerian Sahara. Orient. J. Chem. 2018, 34, 519. [Google Scholar] [CrossRef]
- Dahmane, D.; Dob, T.; Krimat, S.; Nouasri, A.; Metidji, H.; Ksouri, A. Chemical composition, antioxidant and antibacterial activities of the essential oils of medicinal plant Ammodaucus leucotrichus from Algeria. J. Essent. Oil Res. 2017, 29, 48–55. [Google Scholar] [CrossRef]
- Pukalskas, A.; Venskutonis, P.R.; Salido, S.; de Waard, P.; van Beek, T.A. Isolation, identification and activity of natural antioxidants from horehound (Marrubium vulgare L.) cultivated in Lithuania. Food Chem. 2012, 130, 695–701. [Google Scholar] [CrossRef]
- Aliboudhar, H.; Tigrine-Kordjani, N.; Hanifi, N.; Meklati, B.Y. Volatiles profiling and antioxidant activity evaluation of different parts of a medicinal plant: Anacyclus clavatus. J. Herbs Spices Med. Plants. 2013, 19, 33–47. [Google Scholar] [CrossRef]
- Selles, C.; Dib, M.E.; Allali, H.; Tabti, B. Evaluation of antimicrobial and antioxidant activities of solvent extracts of Anacyclus pyrethrum L. from Algeria. Mediterr. J. Chem. 2012, 2, 408–415. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Ismail, A.; Marjan, Z.M.; Foong, C.W. Total antioxidant activity and phenolic content in selected vegetables. Food Chem. 2004, 87, 581–586. [Google Scholar] [CrossRef]
- Awin, T.; Mediani, A.; Shaari, K.; Faudzi, S.M.; Sukari, M.A.; Lajis, N.H.; Abas, F. Phytochemical profiles and biological activities of Curcuma species subjected to different drying methods and solvent systems: NMR-based metabolomics approach. Ind. Crop. Prod. 2016, 94, 342–352. [Google Scholar] [CrossRef]
- Piślewska, M.; Bednarek, P.; Stobiecki, M.; Zieliśska, M.; Wojtaszek, P. Cell wall-associated isoflavonoids and β-glucosidase activity in Lupinus albus plants responding to environmental stimuli. Plant. Cell Environ. 2002, 25, 20–40. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Murtijaya, J. Antioxidant properties of Phyllanthus amarus extracts as affected by different drying methods. LWT-Food Sci. Technol. 2007, 40, 1664–1669. [Google Scholar] [CrossRef]
- Gholamhoseinian, A.; Fallah, H.; Sharifi-far, F.; Mirtajaddini, M. The inhibitory effect of some Iranian plants extracts on the alpha glucosidase. Iran. J. Basic Med. Sci. 2008, 11, 1–9. [Google Scholar]
- Hammami, S.; Li, Z.; Huang, M.; El Mokni, R.; Dhaouadi, H.; Yin, S. New bioactive labdane diterpenoids from Marrubium aschersonii. J. Nat. Prod. 2016, 30, 2142–2148. [Google Scholar] [CrossRef] [PubMed]
- Tahara, S.; Ingham, J.L.; Nakahara, S.; Mizutani, J.; Harborne, J.B. Fungitoxic dihydrofurano-isoflavones and related compounds in white lupin, Lupinus albus. Phytochemistry 1984, 23, 1889–1900. [Google Scholar] [CrossRef]
- Tahara, S.; Orihara, S.; Ingham, J.L.; Mizutani, J. Seventeen isoflavonoids from Lupinus albus roots. Phytochemistry 1989, 28, 901–911. [Google Scholar] [CrossRef]
- Abu Bakar Sajak, A.; Abas, F.; Ismail, A.; Khatib, A. Effect of different drying treatments and solvent ratios on phytochemical constituents of Ipomoea aquatica and correlation with α-glucosidase inhibitory activity. Int. J. Food Prop. 2016, 19, 2817–2831. [Google Scholar] [CrossRef]
- Erbaş, M.; Certel, M.; Uslu, M.K. Some chemical properties of white lupin seeds (Lupinus albus L.). Food Chem. 2005, 89, 341–345. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Rayas-Duarte, P. Composition of Lupinus albus. Cereal Chem. 1995, 72, 643–647. [Google Scholar]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant. Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Fernández-Tomé, S.; Hernández-Ledesma, B. Functionality of Soybean Compounds in the Oxidative Stress-Related Disorders. In Gastrointestinal Tissue; Academic Press: Cambridge, MA, USA, 2017; pp. 339–353. [Google Scholar]
- Miller, N.J.; Rice-Evans, C.A. The relative contributions of ascorbic acid and phenolic antioxidants to the total antioxidant activity of orange and apple fruit juices and blackcurrant drink. Food Chem. 1997, 60, 331–337. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Appelqvist, L.Å. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.; Sousa, J.L.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzym. Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef]
- Banerjee, P.; Ghosh, S.; Dutta, M.; Subramani, E.; Khalpada, J.; Roy Choudhury, S.; Chakravarty, B.; Chaudhury, K. Identification of key contributory factors responsible for vascular dysfunction in idiopathic recurrent spontaneous miscarriage. PLoS ONE 2013, 8, e80940. [Google Scholar] [CrossRef]
- Lee, S.Y.; Mediani, A.; Nur Ashikin, A.H.; Azliana, A.B.; Abas, F. Antioxidant and α-glucosidase inhibitory activities of the leaf and stem of selected traditional medicinal plants. Int. Food Res. J. 2014, 21, 379–386. [Google Scholar]
- Wan, C.; Yuan, T.; Cirello, A.L.; Seeram, N.P. Antioxidant and α-glucosidase inhibitory phenolics isolated from highbush blueberry flowers. Food Chem. 2012, 135, 1929–1937. [Google Scholar] [CrossRef]
- Maulidiani, M.; Sheikh, B.Y.; Mediani, A.; Wei, L.S.; Ismail, I.S.; Abas, F.; Lajis, N.H. Differentiation of Nigella sativa seeds from four different origins and their bioactivity correlations based on NMR-metabolomics approach. Phytochem. Lett. 2015, 13, 308–318. [Google Scholar] [CrossRef]
- Abas, F.; Lajis, N.H.; Israf, D.A.; Khozirah, S.; Kalsom, Y.U. Antioxidant and nitric oxide inhibition activities of selected Malay traditional vegetables. Food Chem. 2006, 95, 566–573. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536. [Google Scholar] [CrossRef]
Sample Availability: Not available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Ethanol Ratio | DPPH Activity | α-Glucosidase Inhibition Activity | Nitric Oxide Inhibition Activity | |||
|---|---|---|---|---|---|---|---|
| % Inhibition (5000 μg/mL) | IC50 μg/mL | % Inhibition (5000 μg/mL) | IC50 | % Inhibition μg/mL | Cell Viability (5000 μg/mL) | ||
| Peganum harmala L. | 100 | 10.89 ± 2.62 De | ND | ND | ND | 6.44 ± 0.76 De | 83.63 ± 0.48 |
| 80 | 15.42 ± 1.82 Cf | ND | ND | ND | 8.65 ± 0.93 Ce | 90.65 ± 0.65 | |
| 50 | 28.13 ± 1.56 Bc | ND | ND | ND | 11.82 ± 1.22 Bd | 97.43 ± 0.78 | |
| 0 | 44.72 ± 2.56 Ab | ND | ND | ND | 39.73 ± 0.94 Aa | 91.34 ± 0.36 | |
| Anacyclus valentinus | 100 | 45.99 ± 1.02 Bb | ND | 8. 13 ± 1.07 BCd | ND | 28.87 ± 1.33 Cc | 67.29 ± 0.38 |
| 80 | 65.57 ± 1.32 Ab | 25.17 ± 1.62 | 47.01 ± 2.88 Ab | ND | 40.29 ± 1.14 Ab | 93.11 ± 0.40 | |
| 50 | 31.89 ± 1.12 Cb | ND | 10.13 ± 1.26 Bb | ND | 32.12 ± 1.57 Bb | 97.35 ± 0.61 | |
| 0 | 25.79 ± 1.24 Dc | ND | 7.67 ± 0.07 Cd | ND | 22.06 ± 1.93 Db | 94.28 ± 0.44 | |
| Zygophyllum album | 100 | 21.44 ± 1.27 Bd | ND | ND | ND | 27.95 ± 1.34 Ac | 38.79 ± 0.55 |
| 80 | 19.26 ± 0.88 Ce | ND | ND | ND | 18.37 ± 1.55 ABd | 96.48 ± 0.63 | |
| 50 | 15.89 ± 0.72 Dd | ND | ND | ND | 13.77 ± 2.04 Bd | 95.81 ± 0.75 | |
| 0 | 48.10 ± 0.66 Aa | ND | ND | ND | 38.28 ± 0.84 ABb | 67.76 ± 0.71 | |
| Ammodaucus leucotrichus | 100 | 81.60 ± 0.56 Aa | 26.26 ± 1.22 | 16.66 ± 1.62 Bc | ND | 50.53 ± 0.62 Aa | 34.52 ± 0.68 |
| 80 | 50.39 ± 1.76 Cc | ND | 26.60 ± 1.87 Ac | ND | 41.12 ± 1.78 Cb | 99.09 ± 0.22 | |
| 50 | 43.73 ± 2.10 Ba | ND | 4.85 ± 1.15 Cc | ND | 44.94 ± 1.58 Ba | 90.24 ± 0.56 | |
| 0 | 49.80 ± 1.84 Da | ND | 16.27 ± 0.86 Bc | ND | 39.67 ± 1.88 Ca | 78.55 ± 0.46 | |
| Marrubium vulgare | 100 | 75.84 ± 2.95 Aa | 19.67 ± 1.33 | 31.93 ± 2.39 Cb | ND | 48.55 ± 1.46 Ab | 41.74 ± 0.38 |
| 80 | 78.57 ± 1.88 Aa | 24.08 ± 2.62 | 68.06 ± 2.15 Aa | 12.66 ± 2.30 | 49.87 ± 1.32 Aa | 99.63 ± 0.40 | |
| 50 | 35.55 ± 2.38 Bb | ND | 43.43 ± 0.10 Ba | ND | 21.76 ± 0.93 Bc | 98.54 ± 0.39 | |
| 0 | 14.21 ± 1.78 Cd | ND | 20.00 ± 3.00 Db | ND | 7.77 ± 0.84 Cc | 99.56 ± 0.29 | |
| Lupinus albus | 100 | 39.59 ± 2.49 Ac | ND | 96.78 ± 1.75 Aa | 6.45 ± 1.30 | 22.56 ± 0.92 ABd | 41.74 ± 0.38 |
| 80 | 36.23 ± 1.67 Bd | ND | 64.57 ± 1.96 Ba | 8.66 ± 1.88 | 20.44 ± 1.06 BCc | 99.63 ± 0.40 | |
| 50 | 39.87 ± 1.55 Cc | ND | 45.59 ± 2.76 Ca | ND | 23.04 ± 1.23 Ac | 98.54 ± 0.39 | |
| 0 | 25.70 ± 1.89 Cc | ND | 43.59 ± 2.44 Ca | ND | 19.67 ± 1.56 Cb | 92.43 ± 0.77 | |
| Curcumin (IC50) | - | - | - | - | - | 10.97 ± 0.81 | - |
| Quercetin | - | 74.84 ± 0.67 | 12.27 ±1.20 | 60.80 ± 1.21 | 8.47 ± 1.57 | - | - |
| Compounds | Chemical Shift | References |
|---|---|---|
| (1) Formic acid | 8.45 (s) | Erbaş et al., 2005 |
| (2) Adenosine | 8.37 (s), 8.23 (s) | - |
| (3) Hydroxybutyrate | 3.58 (m), 2.26 (m), 1.78 (m) | - |
| (4) Asparagine | 2.96 (m), 2.88 (m), 4.02 (dd, 7.5, 4) | - |
| (5) Proline | 4.11 (m), 3.35 (m), 3.30 (m), 2.06 (m), 1.98 (m) | - |
| (6) Thiamine | 9.47 (s), 8.03 (s), 5.47 (s), 3.87 (t, 5.8), 3.20 (t, 5.5), 2.55 (d, 15.5) | - |
| (7) Epicatechin | 7.05 (br s), 6.96 (m), 6.12 (d, 1.5), 6.08 (d,1.5) | - |
| (8) Caprate | 2.15 (br s), 1.46 (br s), 1.25 (br s), 0.86 (br s) | - |
| (9) Kaempferol | 8.02 (d, 8.0), 6.92 (d, 8.0), 6.35 (d, 2.0), 6.20 (d, 2.0) | - |
| (10) Methoxy flavone | 7.16 (d, 2.0), 6.90 (d, 8.5) | - |
| (11) α-Tocopherol | 1.24 (s), 2.05 (m), 1.80 (m), 1.65 (m), 1.52 (m), 1.40 (m), 1.17 (m), 0.88 (d, 6.5) | - |
| (12) Stearic acid | 0.93 (t, 6.5), 1.79 (tt, 7.0), 2.43 (t, 7.5) | - |
| (13) Palmitic acid | 2.37 (t, 4.0), 1.70 (m), 1.47 (m), 0.89 (t, 18.0) | - |
| (14) Oleic acid | 5.35 (m), 2.40 (t, 4.0), 2.03 (m), 1.60 (m), 0.89 (t, 20) | - |
| (15) Linoleic acid | 5.44 (m), 2.91 (t, 6.0), 2.28 (t, 7.0), 2.02 (m), 1.70 (m), 1. 45 (m), 0.93 (t, 6.5) | - |
| (16) Oleanolic acid | 5.14 (br s), 2.89 (m), 2.73 (m), 1.95 (m), 1.84 (d, 6.0), 1.47 (m), 1.32 (m), 1.09 (m), 0.86 (t,12.5) | - |
| (17) Betulinic acid | 4.94 (s), 3.55 (m), 2.75 (m), 2.27 (m), 1.78 (m) | - |
| (18) Lupeol | 1.64 (s), 4.55 (s), 4.66 (s), 2.37 (m), 3.2 (m), 2.22 (d, 5.5) | - |
| (19) Gallic acid | 7.00 (s) | - |
| (20) Hydroxyiso lupalbigenin | 1.65 (s), 1.78 (s), 1.86 (s), 6.68 (s), 8.29 (s), 12.50 (s), 6.56 (d, 8.30), 5.32 (br t,7.0) | - |
| (21) Wighteon | 1.65 (s), 1.78 (s), 6.49 (s), 5.28 (br t), 3.37 (br d) | - |
| (22) Luteone | 1.78 (s), 1.65 (s), 5.32 (br t, 7.0), 3.44 (d, 7.0) | - |
| (23) Lupisoflavone | 3.88 (m), 7.27 (d, 2.0), 6.88 (d, 8.2) | - |
| (24) Lupinisoflavone A | 6.63 (s), 13.30 (s), 8.35 (s), 7.24 (d, 9.0) | - |
| (25) Lupinoisolone A | 1.25 (br s), 1.36 (s), 1.70 (br s), 6.33 (s), 8.18 (s), 13.40 (s), 3.89 (m), 6.88 (d), (5.33 br t, 7.0), 3.38 (br d, 6.8) | - |
| (26) Lupinoisolone C | 1.28 (s), 1.62 (s), 1.77 (br s), 7. 94 (s), 13.41 (s), 6.41 (d,8.0), 6.44 (br s), 7.03 (d, 8.2), 5.31 (br t, 7.1), 3.75 (br t, 7.0), 3.35 (br d, 7.0) | |
| (27) Lupinisol A | 1.72 (s), 1.89 (s), 6.45 (s), 7.33 (s), 8.12 (s), 13.58 (s), 4.72 (brs), 4.95 (br s), 3.00 (m), 4.4 (m), 7.29 (dd, 8.3, 2.0), 6.89 (d, 8.3), 5.35 (br t,7.0), 3.35 (br d, 7.0) | Tahara et al., 1984 |
| (28) Lupinisol B | 6.52 (s), 8.18 (s), 13.00 (s), 4.77 (br s), 4.94 (br s), 6.95 (d, 8.0), 5.32 (br t, 7.0), 4.48 (dd, 7.0,4.0), 3.48(br d,5.0) | |
| (29) Lipinisol C | 8.14 (s), 6.56 s, 13.01 (s), 4.85 (br s), 5.04 (br s), 7.05 (d, 7.0), 5.28 (br t, 7.3), 4.37 (br d, 5.0), 3.38 (br d, 5.0) | - |
| (30) Chandalone | 1.47 (s), 6.32 (s), 8.14 (s), 13.46 (s),1.25 (br s), 7.35 (d, 2.0), 7.29 (dd, 2.0, 7.0) 6.88 (d, 2.0), 6.69 (d, 9.0), 5.79 (d, 10.0), 5.33 (br t), 3.38 (br, 5.5) | |
| (31) Isoderrone | 1.41 (s), 8.25 (s), 13.00 (s), 7.29 (d, 2.0), 6.48 (d, 2.0), 6.44 (d, 1.5) 6.31 (d, 1.5), 5.77 (d, 5.0) | - |
| (32) Lupinalbin F | 1.72 (s), 1.89 (s), 7.33 (s), 8.12 (s), 13.58 (s), 4.72 (br s), 4.95 (br s), 3.00 (m), 4.40 (m), 7.29 (dd, 8.3, 2.0), 6.89 (d, 8.3), 6.45 (s), 5.35 (br t, 7.0), 3.35 (br d, 7.0) | |
| (33) Lupinoisoflavone G | 1.25 (s), 1.29 (s), 6.37 (s), 8.14 (s), 1.73 (br s), 7.35 (d, 2.0), 7.29 (dd, 8.0,2.0). 5.33 (br t, 7.1), 3.18 (br d, 8.0) | - |
| (34) Genistein | 6.39 (s), 7.69 (s), 7.32 (d, 7.0), 6.33 (s), 6.95 (d, 7.5) | |
| (35) Unknown | 1.24 (s) | - |
| (36) Unknown | 5.22 (s) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hellal, K.; Maulidiani, M.; Ismail, I.S.; Tan, C.P.; Abas, F. Antioxidant, α-Glucosidase, and Nitric Oxide Inhibitory Activities of Six Algerian Traditional Medicinal Plant Extracts and 1H-NMR-Based Metabolomics Study of the Active Extract. Molecules 2020, 25, 1247. https://doi.org/10.3390/molecules25051247
Hellal K, Maulidiani M, Ismail IS, Tan CP, Abas F. Antioxidant, α-Glucosidase, and Nitric Oxide Inhibitory Activities of Six Algerian Traditional Medicinal Plant Extracts and 1H-NMR-Based Metabolomics Study of the Active Extract. Molecules. 2020; 25(5):1247. https://doi.org/10.3390/molecules25051247
Chicago/Turabian StyleHellal, Khaoula, M. Maulidiani, Intan Safinar Ismail, Chin Ping Tan, and Faridah Abas. 2020. "Antioxidant, α-Glucosidase, and Nitric Oxide Inhibitory Activities of Six Algerian Traditional Medicinal Plant Extracts and 1H-NMR-Based Metabolomics Study of the Active Extract" Molecules 25, no. 5: 1247. https://doi.org/10.3390/molecules25051247
APA StyleHellal, K., Maulidiani, M., Ismail, I. S., Tan, C. P., & Abas, F. (2020). Antioxidant, α-Glucosidase, and Nitric Oxide Inhibitory Activities of Six Algerian Traditional Medicinal Plant Extracts and 1H-NMR-Based Metabolomics Study of the Active Extract. Molecules, 25(5), 1247. https://doi.org/10.3390/molecules25051247