Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass




Abstract
1. Introduction
2. Results
2.1. Solid-State Fermentation Kinetics
2.2. Optimization of the SSF Steps
2.2.1. Effect of pH and Moisture Level
2.2.2. Enzymatic Hydrolysis
2.3. Kinetics of SSF with Optimized Conditions
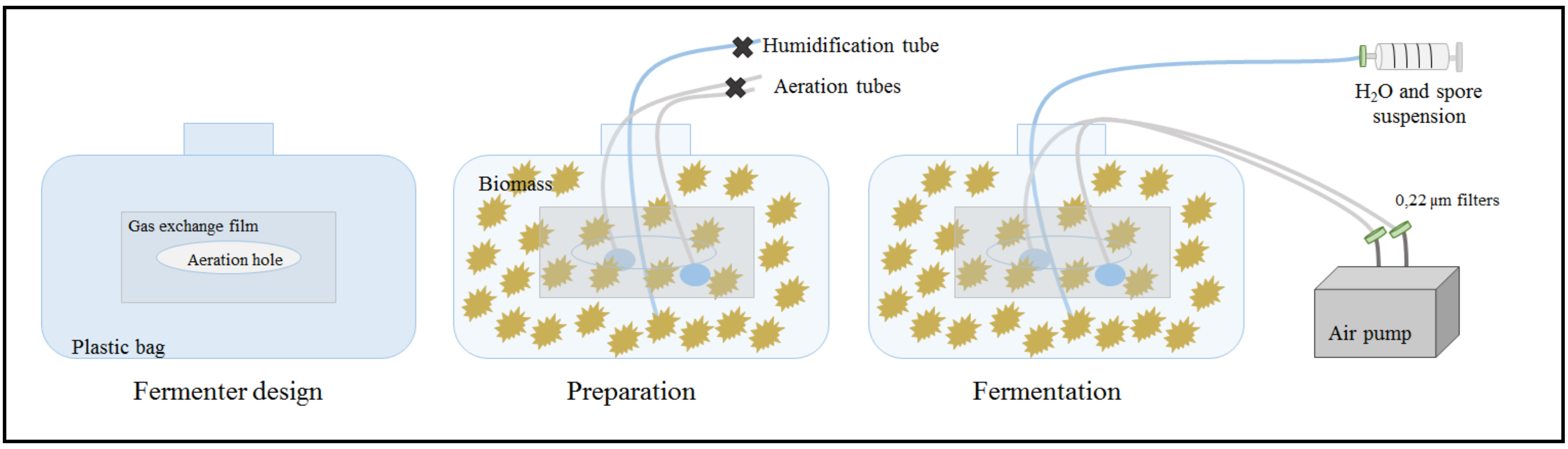
2.4. Larger Scale Fermentation
3. Discussion
4. Materials and Methods
4.1. Feedstock and Microorganisms
4.2. Initial SSF Step
4.2.1. pH and Humidity Level Optimization
4.2.2. Enzyme Production for Biomass Hydrolysis
4.3. Scale-Up Steps
4.4. Analytical Procedure
4.4.1. Mycelial Growth (Protein Assays)
4.4.2. Organic Acid Assays
4.4.3. Enzyme Activity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pandey, A. Solid-state Fermentation in Biotechnology: Fundamentals and Applications; Asiatech Publishers: New Delhi, India, 2001. [Google Scholar]
- Hölker, U.; Lenz, J. Solid-state fermentation—Are there any biotechnological advantages? Curr. Opin. Microbiol. 2005, 8, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Hölker, U.; Höfer, M.; Lenz, J. Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Appl. Microbiol. Biotechnol. 2004, 64, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Dashtban, M.; Schraft, H.; Qin, W. Fungal bioconversion of lignocellulosic residues; opportunities & perspectives. Int. J. Biol. Sci. 2009, 5, 578–595. [Google Scholar] [CrossRef] [PubMed]
- Krishna, C. Solid-state fermentation systems-an overview. Crit. Rev. Biotechnol. 2005, 25, 1–30. [Google Scholar] [CrossRef]
- Rodriguez-Leon, J.A.; Soccol, C.R.; Pandey, A.; Rodriguez, D.E. Factors Affecting Solid-state Fermentation. In Current Developments in Solid-State Fermentation; Pandey, A., Soccol, C.R., Larroche, C., Eds.; Springer: New York, NY, USA, 2008; pp. 26–47. [Google Scholar] [CrossRef]
- Singh nee’ Nigam, P.; Pandey, A. (Eds.) Solid-State Fermentation Technology for Bioconversion of Biomass and Agricultural Residues. In Biotechnology for Agro-Industrial Residues Utilisation; Springer: Dordrecht, Netherlands, 2009; pp. 197–221. [Google Scholar] [CrossRef]
- Christensen, C.H.; Rass-Hansen, J.; Marsden, C.C.; Taarning, E.; Egeblad, K. The Renewable Chemicals Industry. Chemsuschem. 2008, 1, 283–289. [Google Scholar] [CrossRef]
- Bozell, J.J. Feedstocks for the Future—Biorefinery Production of Chemicals from Renewable Carbon. CLEAN Soil Air Water 2008, 36, 641–647. [Google Scholar] [CrossRef]
- Willke, T.; Vorlop, K.D. Industrial bioconversion of renewable resources as an alternative to conventional chemistry. Appl. Microbiol. Biotechnol. 2004, 66, 131–142. [Google Scholar] [CrossRef]
- Gallezot, P. Conversion of biomass to selected chemical products. Chem. Soc. Rev. 2012, 41, 1538–1558. [Google Scholar] [CrossRef]
- Werpy, T.; Holladay, J.; White, J. Top Value Added Chemicals From Biomass: I. Results of Screening for Potential Candidates from Sugars and Synthesis Gas. DOE Scientific and Technical Information.; Pacific Northwest National Lab: Richland, WA, USA, 2004; 419907. [Google Scholar] [CrossRef]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—the US Department of Energy’s ‘Top 10′ revisited. Green Chem. 2010, 12, 539. [Google Scholar] [CrossRef]
- Magnuson, J.K.; Lasure, L.L. Organic Acid Production by Filamentous Fungi. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Tkacz, J.S., Lange, L., Eds.; Springer: New York, NY, USA, 2004; pp. 307–340. [Google Scholar] [CrossRef]
- Goldberg, I.; Rokem, J.S.; Pines, O. Organic acids: Old metabolites, new themes. J. Chem. Technol. Biotechnol. 2006, 81, 1601–1611. [Google Scholar] [CrossRef]
- Tsao, G.T.; Cao, N.J.; Du, J.; Gong, C.S. Production of multifunctional organic acids from renewable resources. Adv. Biochem. Eng. Biotechnol. 1999, 65, 243–280. [Google Scholar] [CrossRef] [PubMed]
- Liaud, N.; Giniés, C.; Navarro, D.; Fabre, N.; Crapart, S.; Gimbert, I.H.; Levasseur, A.; Raouche, S.; Sigoillot, J.C. Exploring fungal biodiversity: organic acid production by 66 strains of filamentous fungi. Fungal Biol. Biotechnol. 2014, 1, 1. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, S.; Huang, H.; Wen, J. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol. Adv. 2012, 30, 1685–1696. [Google Scholar] [CrossRef]
- Yahiro, K.; Shibata, S.; Jia, S.R.; Park, W.; Okabe, M. Efficient itaconic acid production from raw corn starch. J. Ferment. Bioeng. 1997, 84, 375–377. [Google Scholar] [CrossRef]
- Hevekerl, A.; Kuenz, A.; Vorlop, K.D. Filamentous fungi in microtiter plates—An easy way to optimize itaconic acid production with Aspergillus terreus. Appl. Microbiol. Biotechnol. 2014, 98, 6983–6989. [Google Scholar] [CrossRef]
- Willke, T.; Vorlop, K.D. Biotechnological production of itaconic acid. Appl. Microbiol. Biotechnol. 2001, 56, 289–295. [Google Scholar] [CrossRef]
- Kanamasa, S.; Dwiarti, L.; Okabe, M.; Park, E.Y. Cloning and functional characterization of the cis-aconitic acid decarboxylase (CAD) gene from Aspergillus terreus. Appl. Microbiol. Biotechnol. 2008, 80, 223–229. [Google Scholar] [CrossRef]
- Okabe, M.; Lies, D.; Kanamasa, S.; Park, E.Y. Biotechnological production of itaconic acid and its biosynthesis in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2009, 84, 597–606. [Google Scholar] [CrossRef]
- Begum, M.F.; Alimon, A.R. Bioconversion and saccharification of some lignocellulosic wastes by Aspergillus oryzae ITCC-4857.01 for fermentable sugar production. Electron. J. Biotechnol. 2011, 14, 5. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Huang, M.C.; Lin, S.F.; Su, Y.C. Method for the production of itaconic acid using Aspergillus terreus solid state fermentation. U.S. Patent 6171831, 09 January 2001. [Google Scholar]
- West, T.P. Fumaric acid production by Rhizopus oryzae on corn distillers’ grains with solubles. Res. J. Microbiol. 2008, 3, 35–40. [Google Scholar] [CrossRef]
- Jiménez-Quero, A.; Pollet, E.; Zhao, M.; Marchioni, E.; Averous, L.; Phalip, V. Itaconic and fumaric acid production from biomass hydrolysates by Aspergillus strains. J. Microbiol. Biotechnol. 2016, 26, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Quero, A.; Pollet, E.; Zhao, M.; Marchioni, E.; Averous, L.; Phalip, V. Fungal fermentation of lignocellulosic biomass for itaconic and fumaric acid production. J. Microbiol. Biotechnol. 2017, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Te Biesebeke, R.; Ruijter, G.; Rahardjo, Y.S.P.; Hoogschagen, M.J.; Heerikhuisen, M.; Levin, A.; van Driel, K.G.A.; Schutyser, M.A.I.; Dijksterhuis, J.; Zhu, Y.; et al. Aspergillus oryzae in solid-state and submerged fermentations. Progress report on a multi-disciplinary project. Fems Yeast Res. 2002, 2, 245–248. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ummalyma, S.B.; Supriya, R.D.; Sindhu, R.; Binod, P.; Nair, R.B.; Pandey, A.; Gnansounou, E. Biological pretreatment of lignocellulosic biomass—Current trends and future perspectives. In Second and Third Generation of Feedstocks; Elsevier: Alpharetta, GA, USA, 2019; pp. 197–212. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of lignocellulosic biomass: biochemical and molecular perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef]
- Gervais, P.; Molin, P. The role of water in solid-state fermentation. Biochem. Eng. J. 2003, 13, 85–101. [Google Scholar] [CrossRef]
- Mondala, A.H. Direct fungal fermentation of lignocellulosic biomass into itaconic, fumaric, and malic acids: current and future prospects. J. Ind. Microbiol. Biotechnol. 2015, 42, 487–506. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Chio, C.; Yang, C.; Su, E.; Jin, Y.; Qin, W. Delignification overmatches hemicellulose removal for improving hydrolysis of wheat straw using the enzyme cocktail from Aspergillus niger. Bioresour. Technol. 2019, 274, 459–467. [Google Scholar] [CrossRef]
- Sandhya, C.; Sumantha, J.; Szakacs, G.; Pandey, A. Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Process Biochem. 2005, 40, 2689–2694. [Google Scholar] [CrossRef]
- Koser, S.; Anwar, Z.; Iqbal, Z.; Anjum, A.; Aqil, T.; Mehmood, S.; Irshad, M. Utilization of Aspergillus oryzae to produce pectin lyase from various agro-industrial residues. J. Radiat. Res. Appl. Sci. 2014, 7, 327–332. [Google Scholar] [CrossRef]
- Viniegra-González, G.; Favela-Torres, E.; Aguilar, C.N.; de Rómero-Gomez, S.J.; Díaz-Godínez, G.; Augur, C. Advantages of fungal enzyme production in solid state over liquid fermentation systems. Biochem. Eng. J. 2003, 13, 157–167. [Google Scholar] [CrossRef]
- Ayyachamy, M.; Gupta, V.K.; Cliffe, F.E.; Tuohy, M.G. Enzymatic Saccharification of Lignocellulosic Biomass. In Laboratory Protocols in Fungal Biology; Gupta, V.K., Tuohy, M.G., Ayyachamy, M., Turner, K.M., O’Donovan, A., Eds.; Springer: New York, NY, USA, 2013; pp. 475–481. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, S.; Fang, H. Consolidated bioprocessing of lignocellulosic biomass to itaconic acid by metabolically engineering Neurospora crassa. Appl. Microbiol. Biotechnol. 2018, 102, 9577–9584. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, H.H.; Tharmasothirajan, A.; Track, E.; Blank, L.M.; Wierckx, N. Engineering the morphology and metabolism of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Metab. Eng. 2019, 54, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Nemestóthy, N.; Bakonyi, P.; Komáromy, P.; Bélafi-Bakó, K. Evaluating aeration and stirring effects to improve itaconic acid production from glucose using Aspergillus terreus. Biotechnol. Lett. 2019, 41, 1383–1389. [Google Scholar] [CrossRef]
- Molnar, A.P.; Németh, Z.; Kollath, I.S.; Fekete, E.; Flipphi, M.; Ag, N.; Karaffa, L. High oxygen tension increases itaconic acid accumulation, glucose consumption, and the expression and activity of alternative oxidase in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2018, 102, 8799–8808. [Google Scholar] [CrossRef]
- Chenyu Du, A.A. Fermentative Itaconic Acid Production. J. Biodivers. Bioprospecting Dev. 2014, 1, 2. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-Amylase activity | 18.10 |
| Cellulase activity (cellulose) | 4.69 |
| Endoxylanase activity | 70.30 |
| Cellulase activity (xyloglucan) | 10.11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Quero, A.; Pollet, E.; Avérous, L.; Phalip, V. Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass. Molecules 2020, 25, 1070. https://doi.org/10.3390/molecules25051070
Jiménez-Quero A, Pollet E, Avérous L, Phalip V. Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass. Molecules. 2020; 25(5):1070. https://doi.org/10.3390/molecules25051070
Chicago/Turabian StyleJiménez-Quero, Amparo, Eric Pollet, Luc Avérous, and Vincent Phalip. 2020. "Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass" Molecules 25, no. 5: 1070. https://doi.org/10.3390/molecules25051070
APA StyleJiménez-Quero, A., Pollet, E., Avérous, L., & Phalip, V. (2020). Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass. Molecules, 25(5), 1070. https://doi.org/10.3390/molecules25051070