
Ionone Is More than a Violet’s Fragrance: A Review





Abstract
1. Chemical Structure, Physicochemical Properties and Natural Occurrence
2. Synthesis
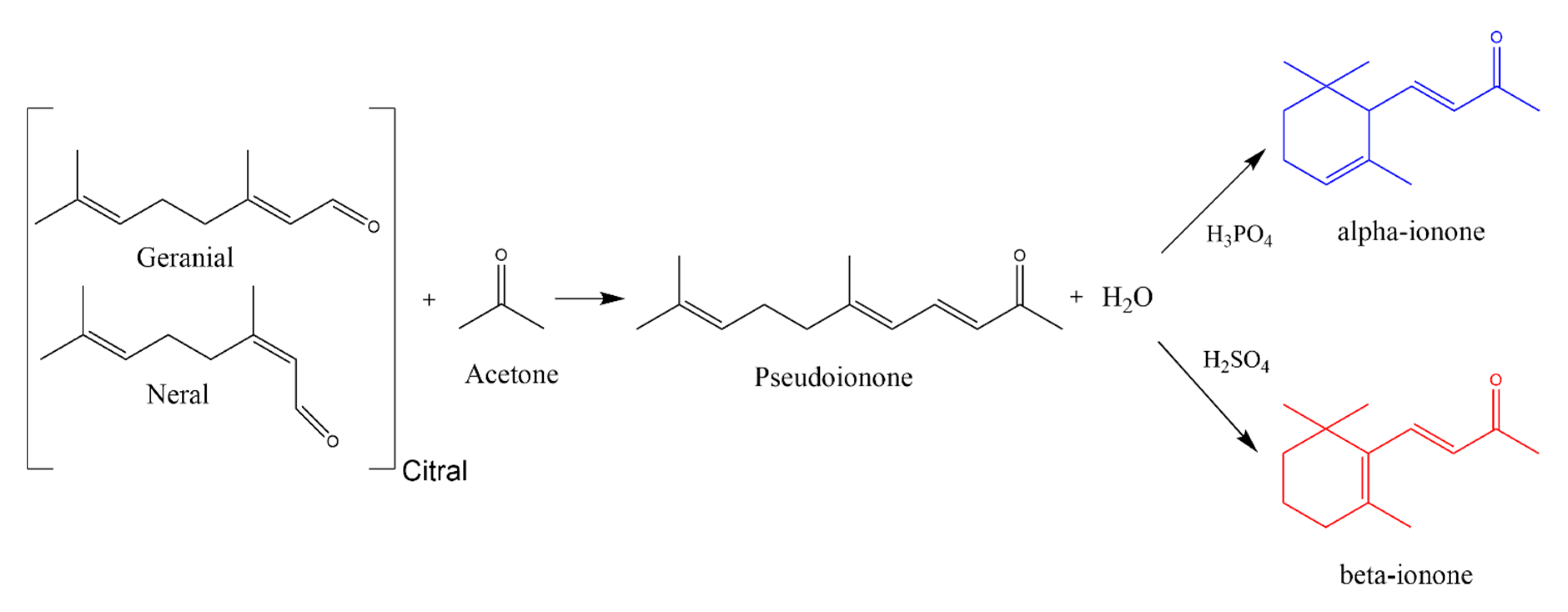
2.1. Chemical Synthesis
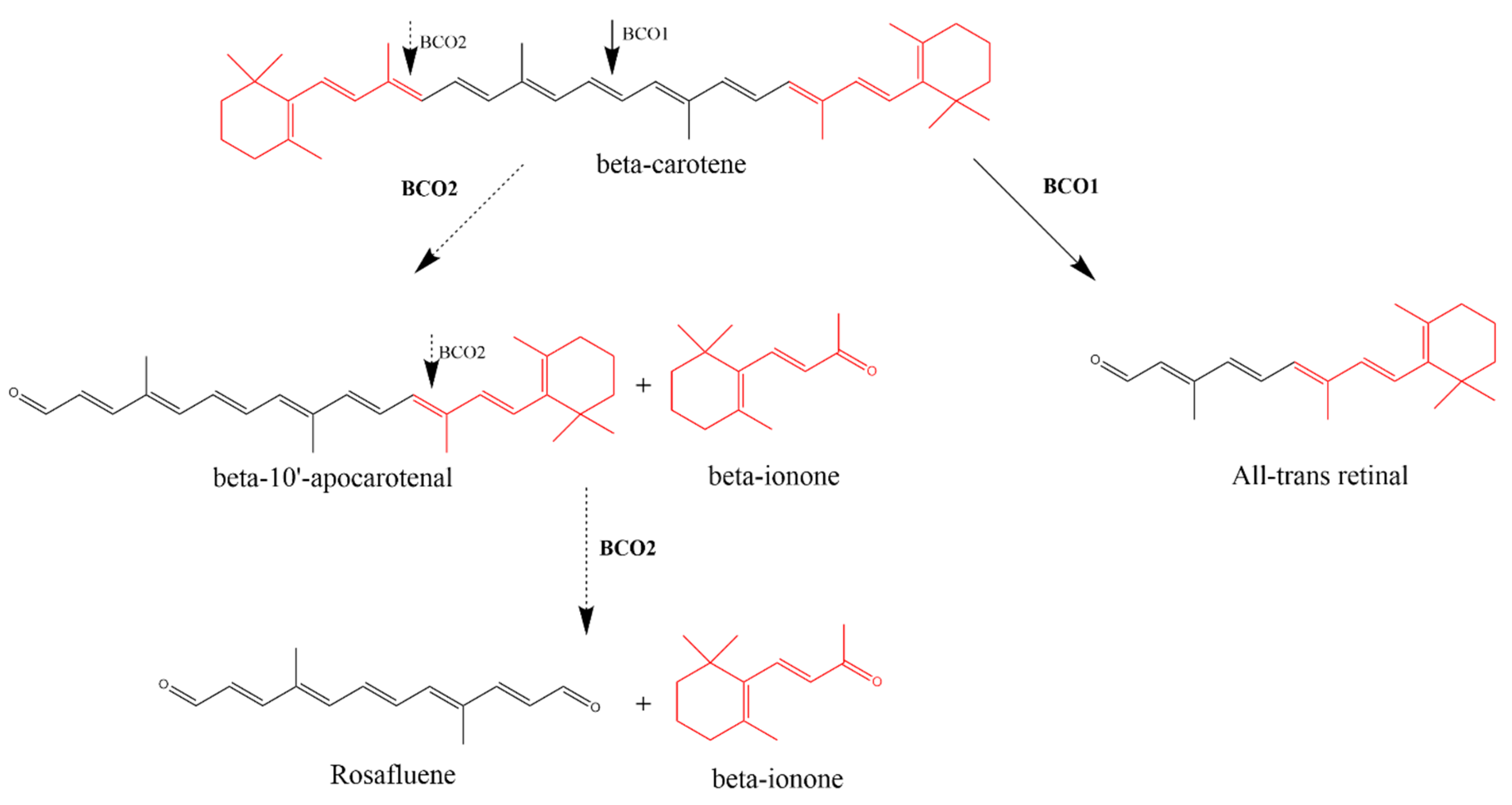
2.2. Endogenous Synthesis
2.2.1. β-Carotene Oxygenase 1 (BCO1)
2.2.2. β-Carotene Oxygenase 2 (BCO2)-Mediated Synthesis
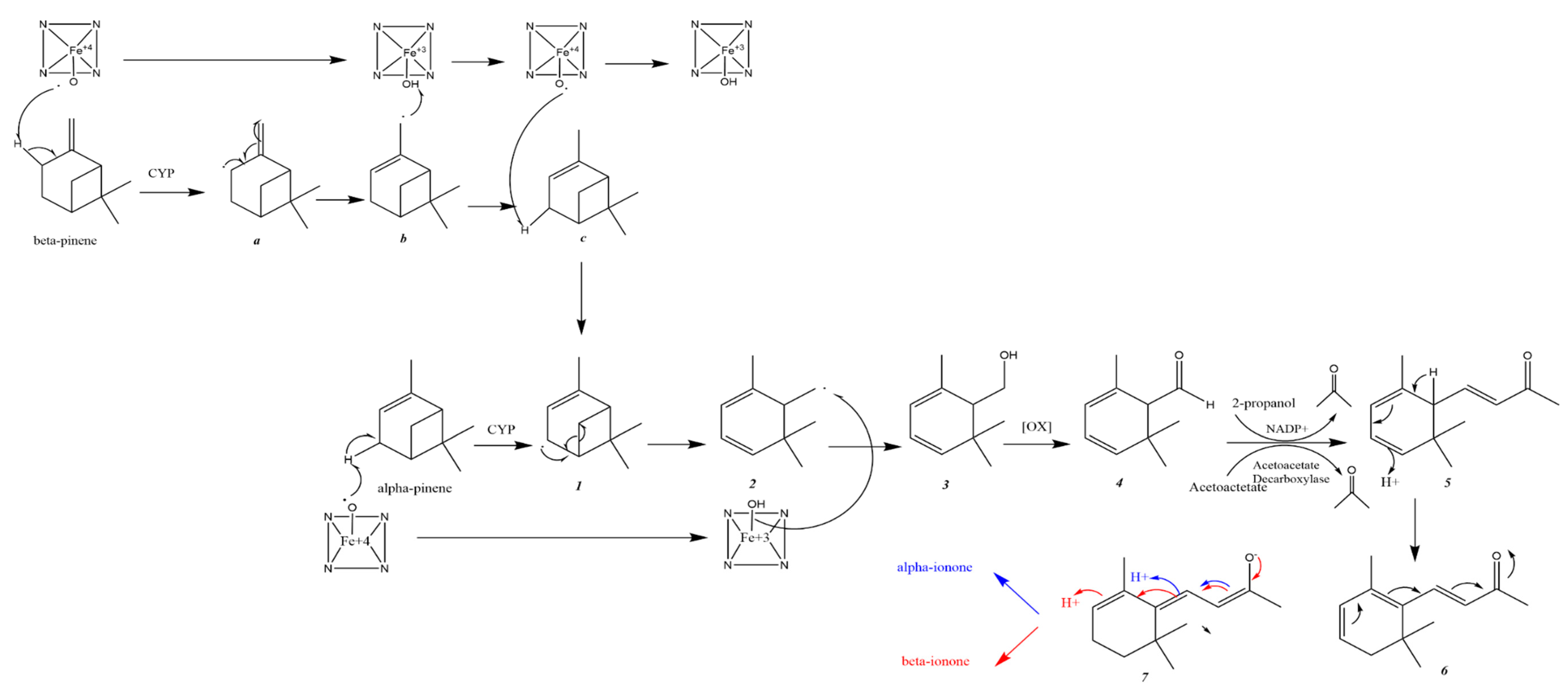
2.3. Possible Ionone Synthesis from Pinene
3. Olfactory Receptors
4. Ionone Effects
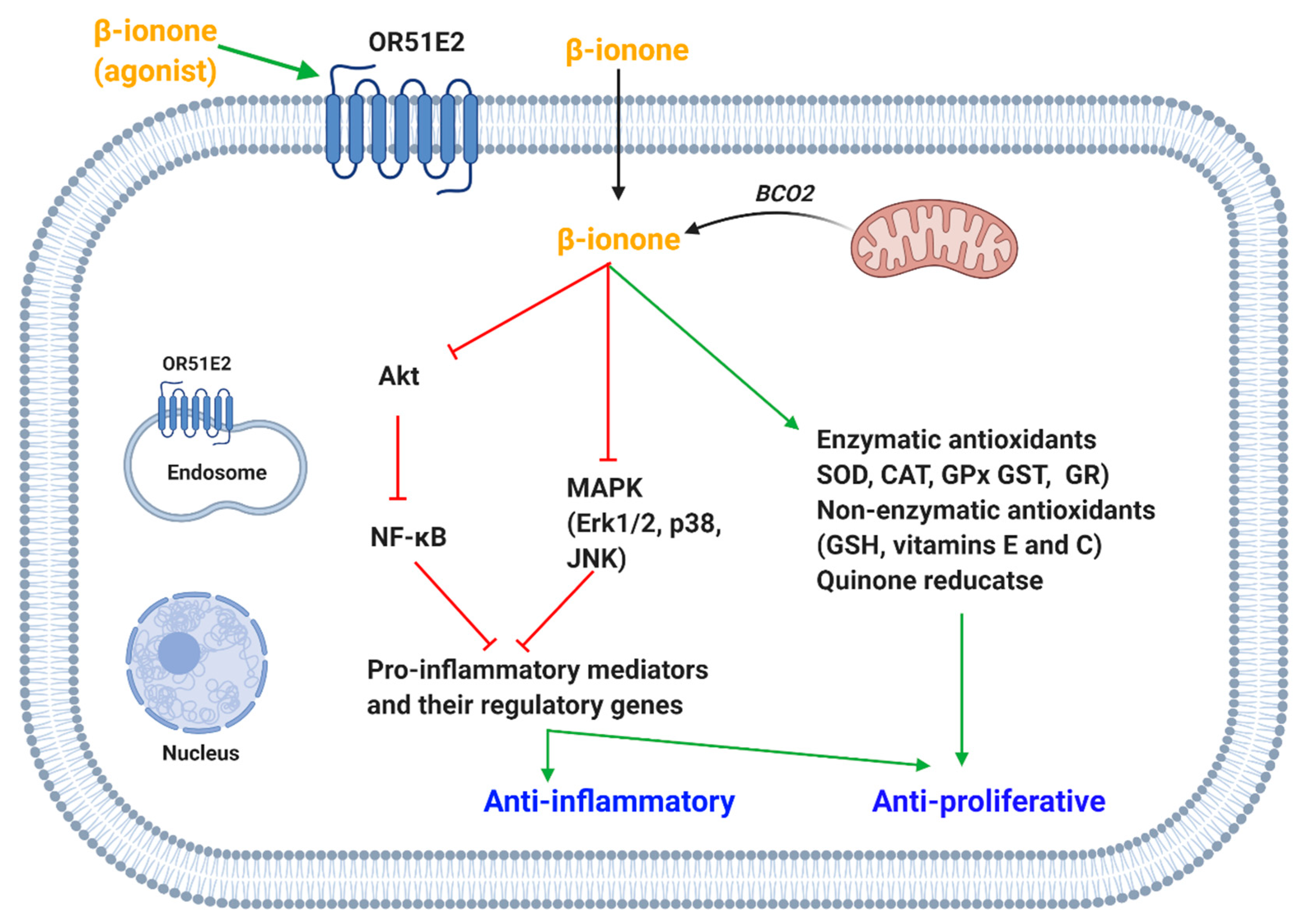
4.1. β-Ionone
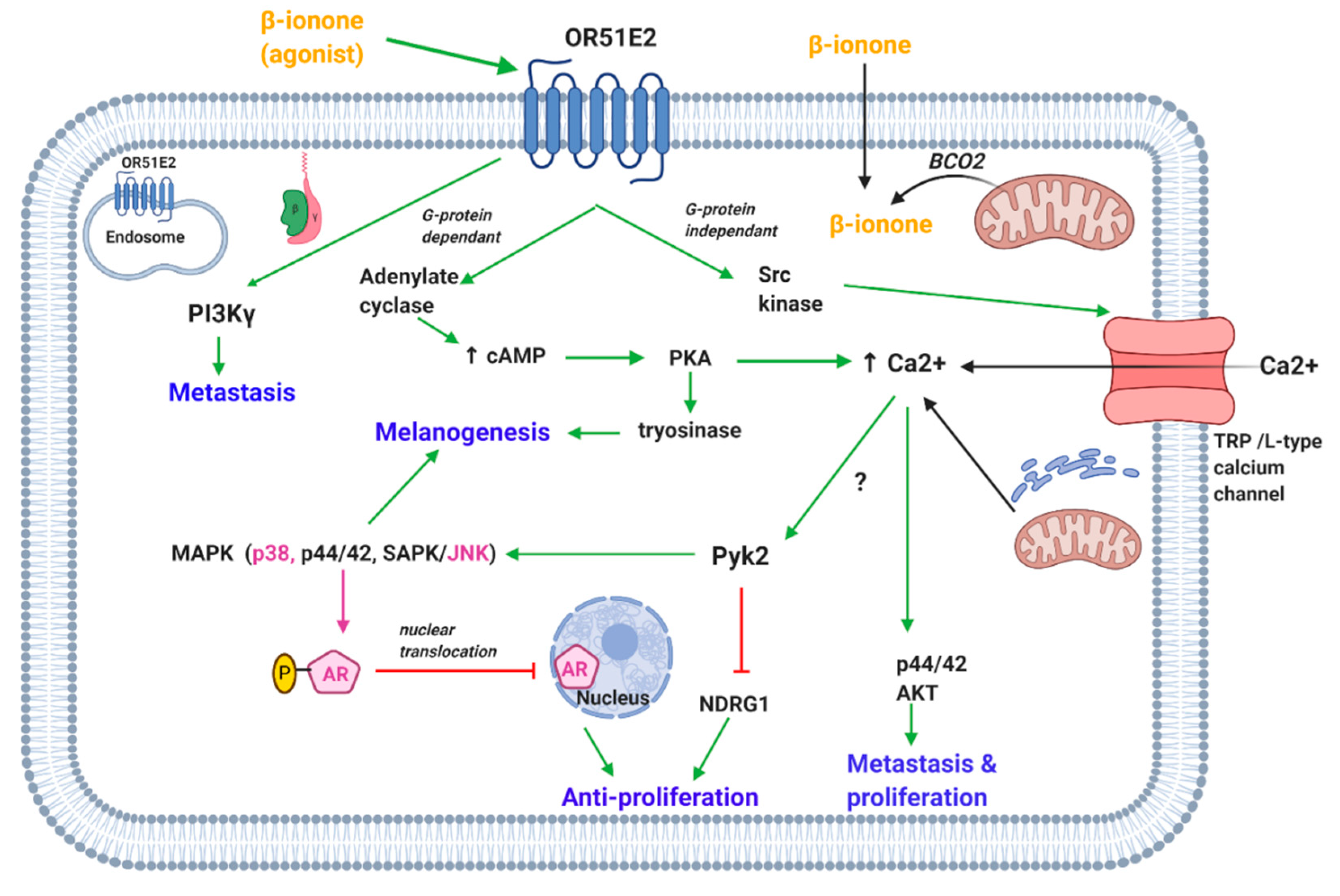
4.1.1. OR51E2-Mediated Effects
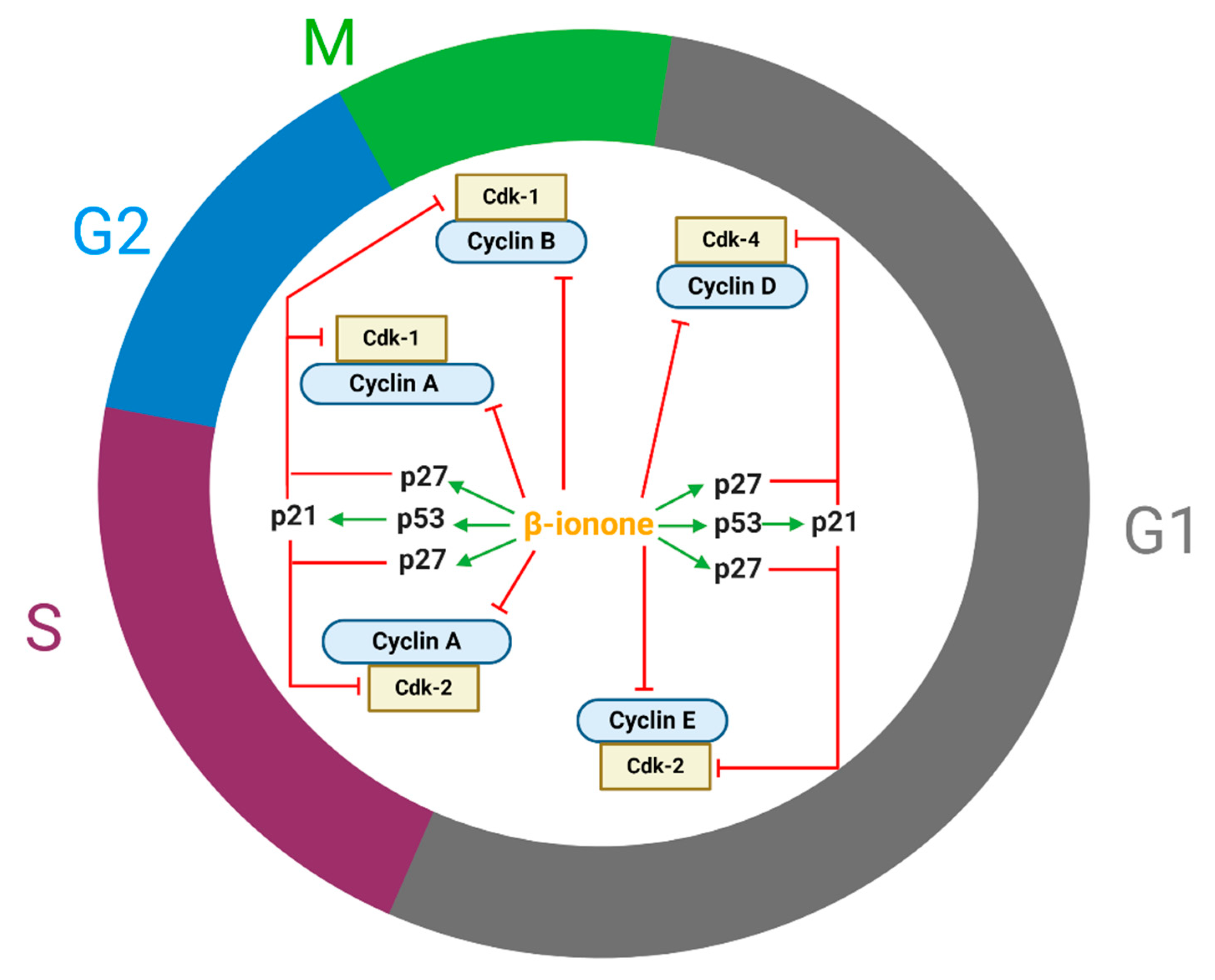
4.1.2. Cell Cycle Regulatory Protein-Mediated Effects
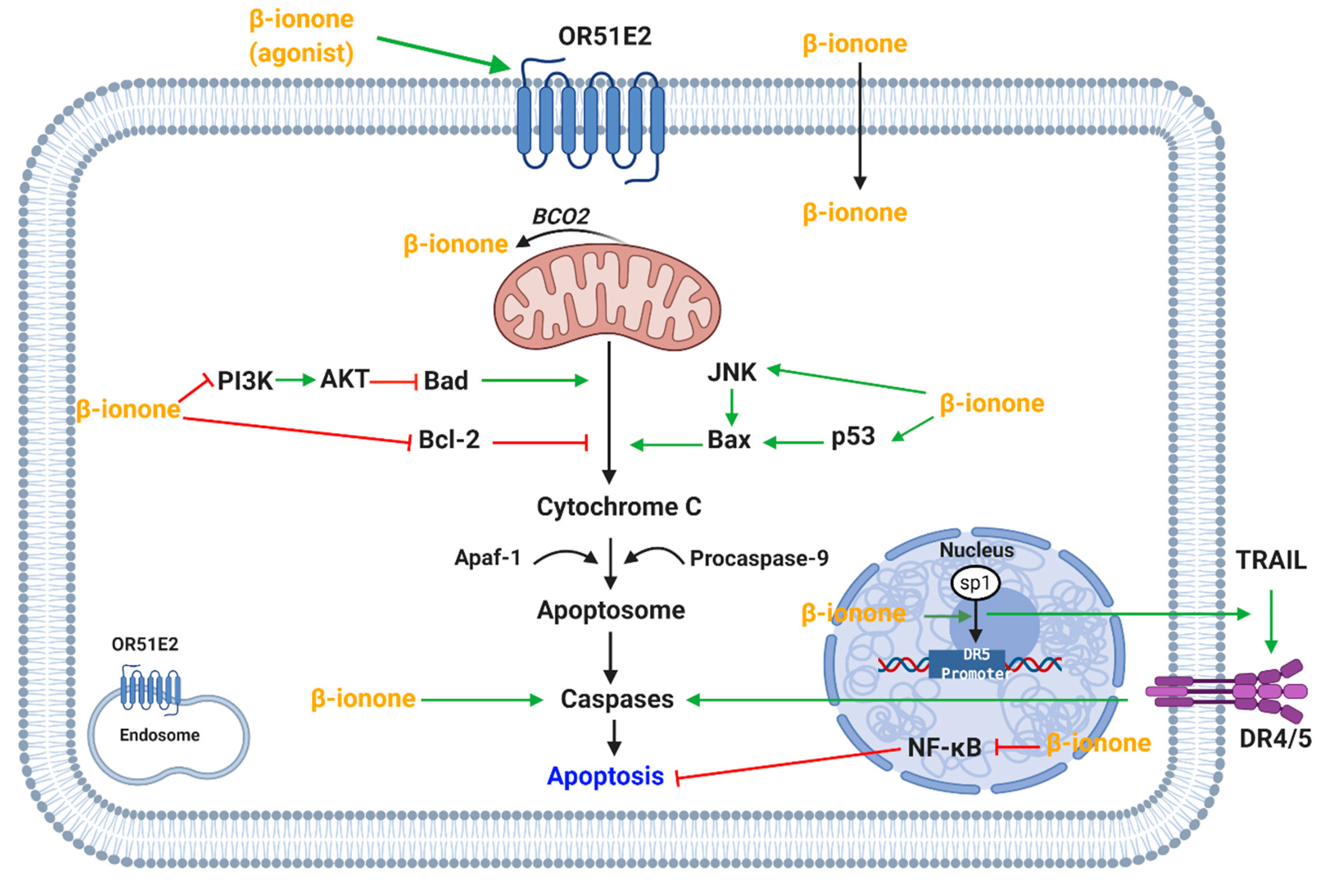
4.1.3. Apoptotic Effect
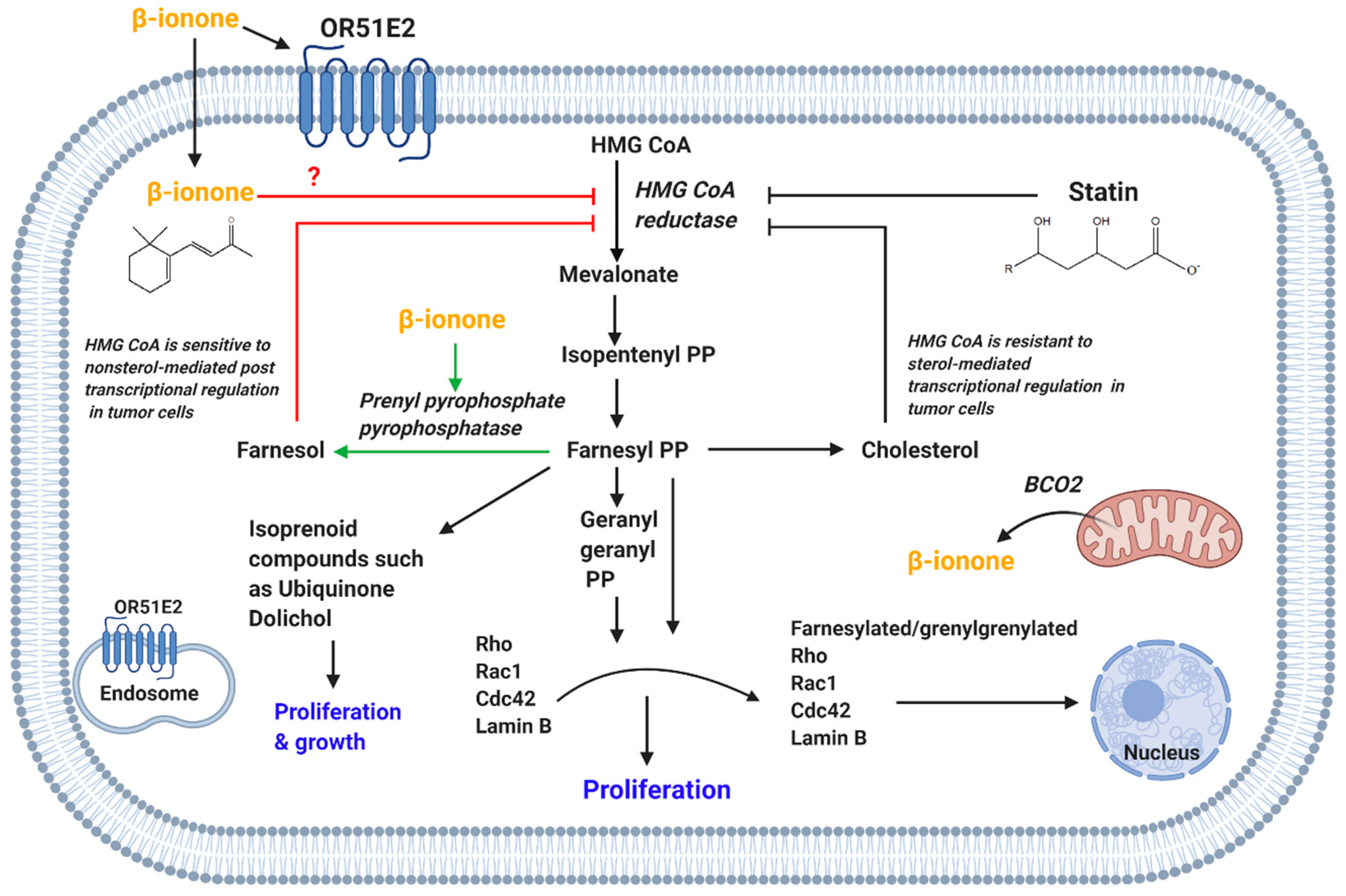
4.1.4. HMG CoA Reductase-Mediated Effect
4.1.5. Antioxidant-Mediated Effect
4.1.6. Pro-Inflammatory Molecules-Mediated Effects
4.1.7. Antimicrobial Effects
4.2. α-Ionone
4.3. Ionone Derivatives
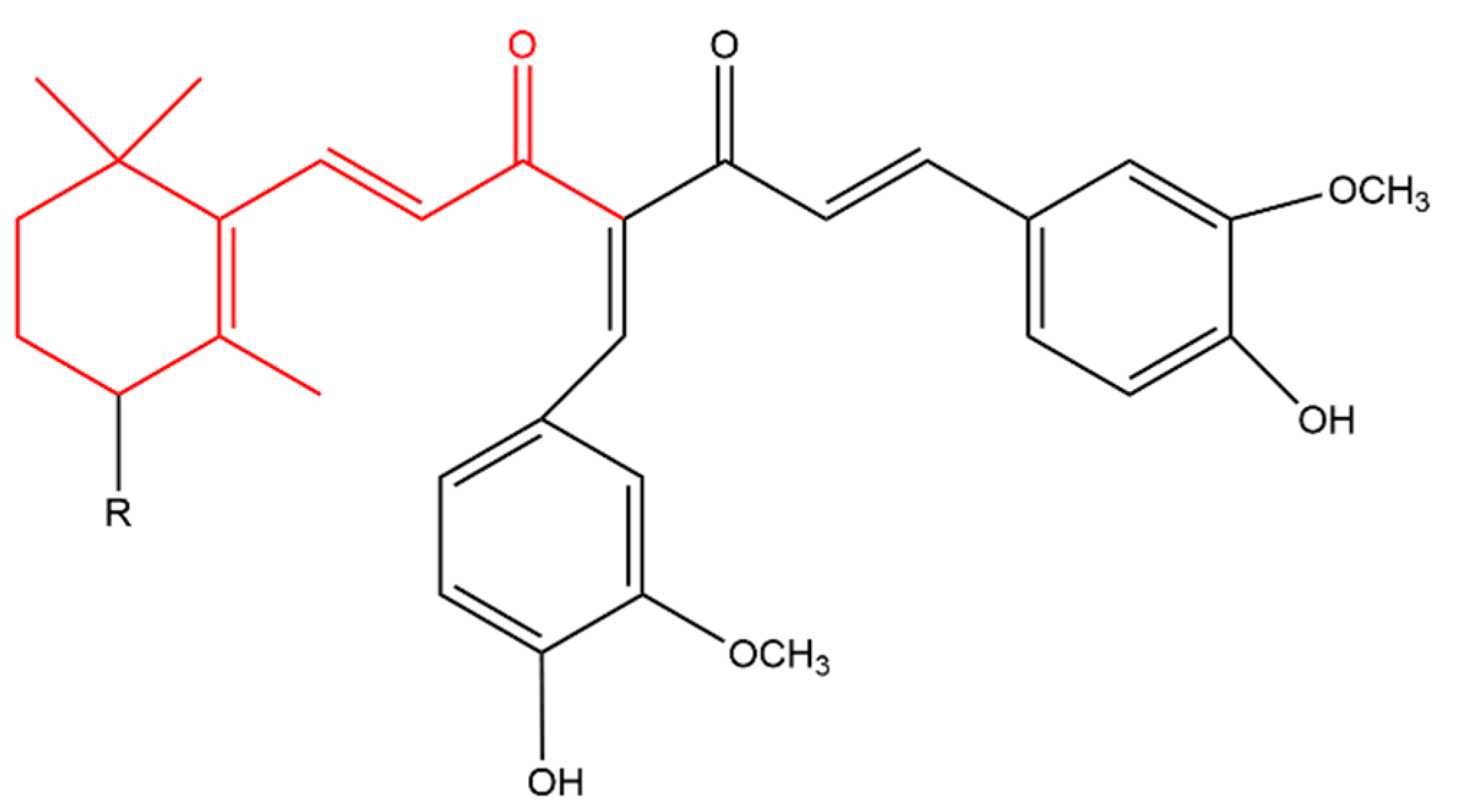
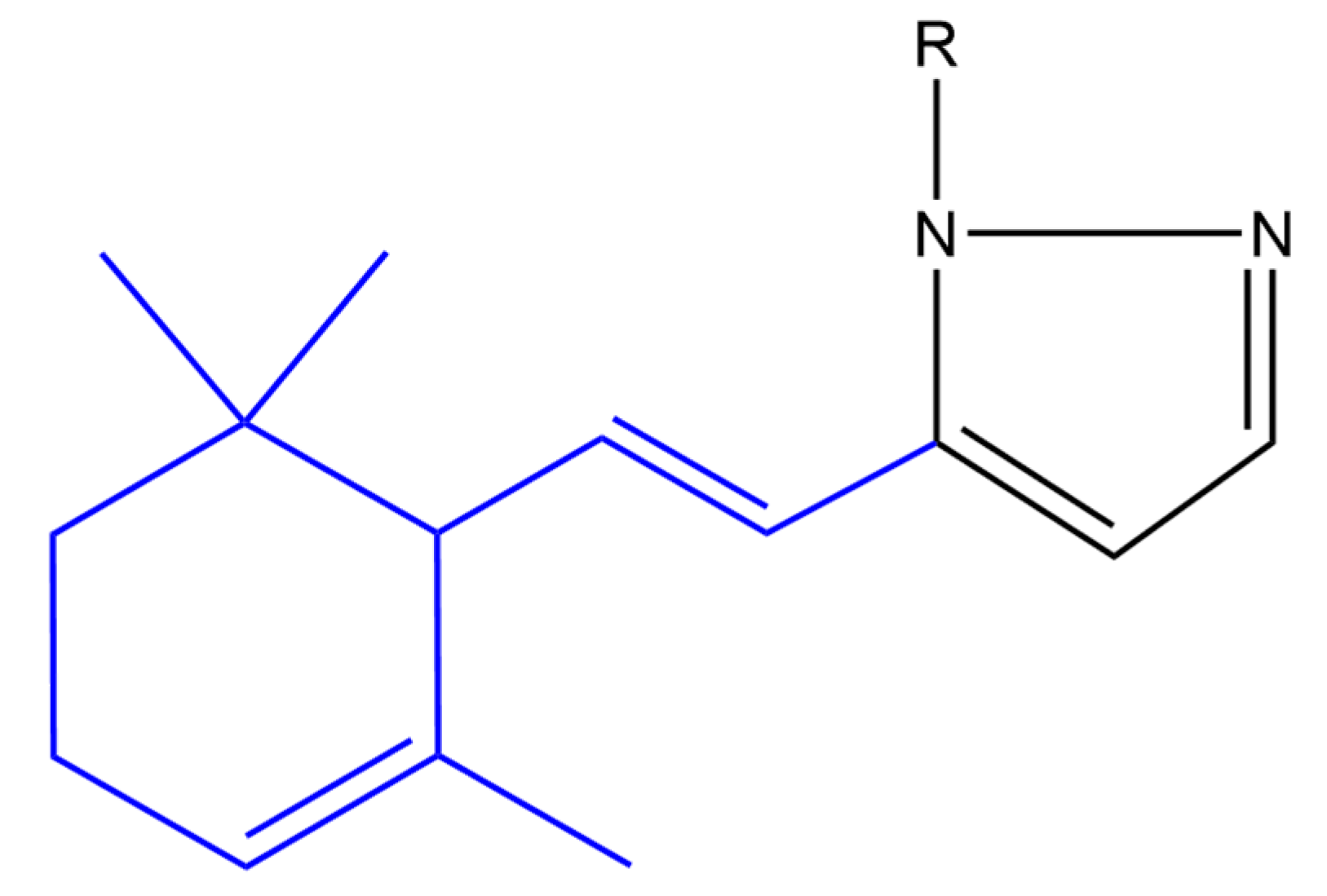
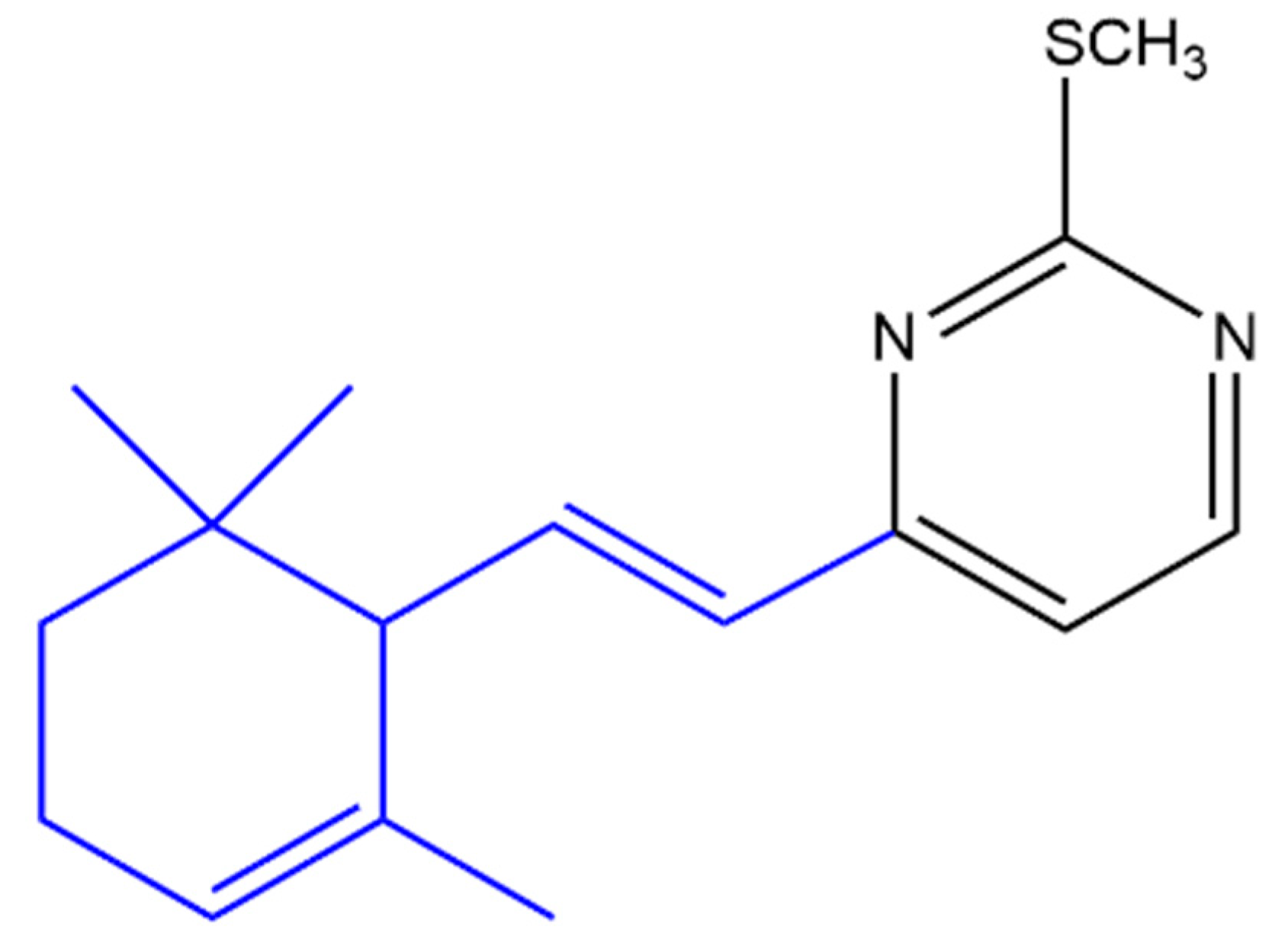
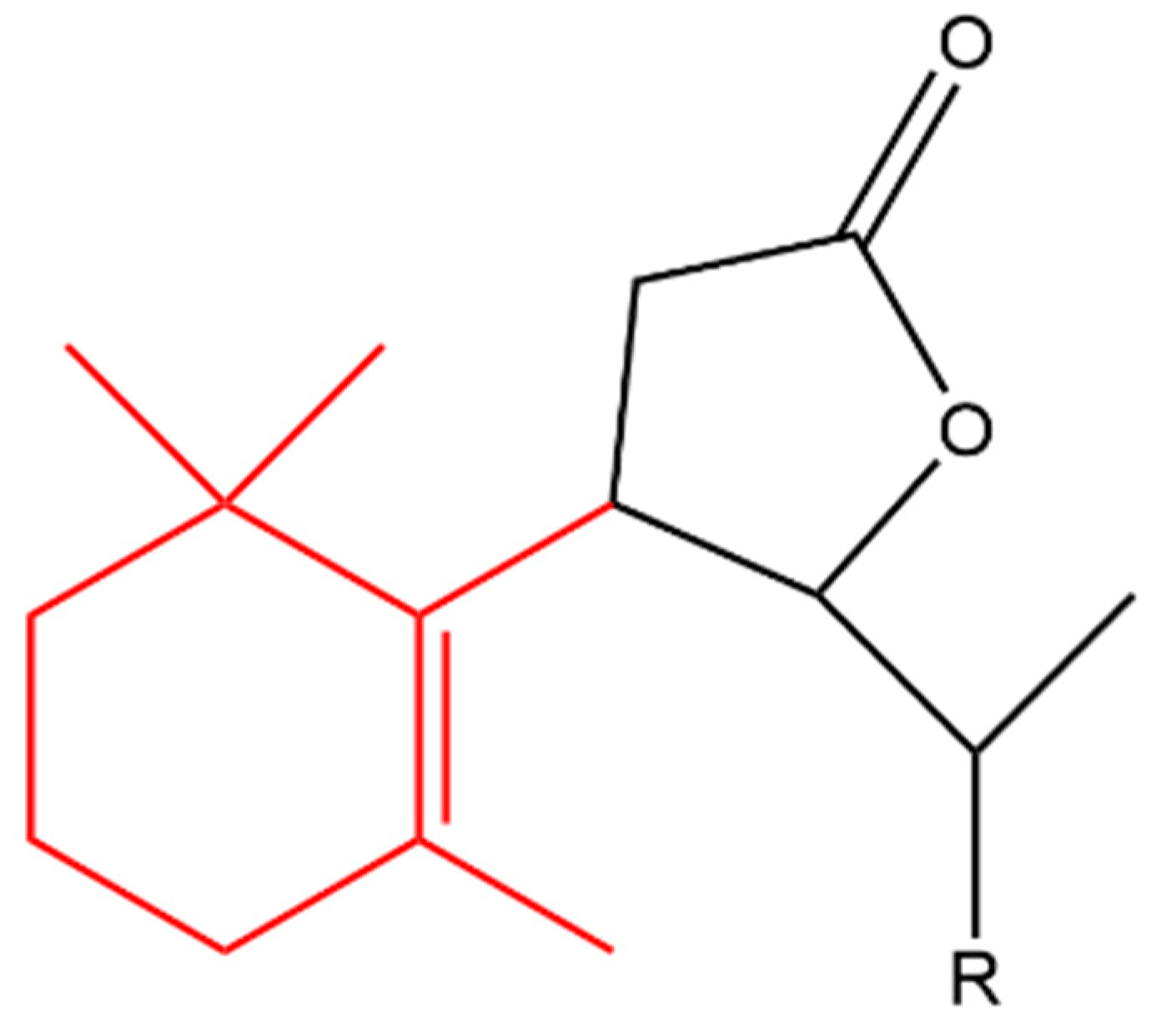
4.3.1. Anti-Cancer
4.3.2. Anti-Inflammatory
4.3.3. Antimicrobial Effect
5. Conclusions and Outlook
Funding
Acknowledgments
Conflicts of Interest
References
- Pinto, F.C.M.; De-Carvalho, R.R.; De-Oliveira, A.; Delgado, I.F.; Paumgartten, F.J.R. Study on the developmental toxicity of beta-ionone in the rat. Regul. Toxicol. Pharmacol. 2018, 97, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Petroianu, G.A.; Stegmeier-Petroianu, A.; Lorke, D.E. Cleopatra: From turpentine and juniper to ionone and irone. Pharmazie 2018, 73, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Mo, H.; Elson, C.E. Apoptosis and cell-cycle arrest in human and murine tumor cells are initiated by isoprenoids. J. Nutr. 1999, 129, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.L.; Chichester, C.O. Metabolism and nutritional significance of carotenoids. Annu. Rev. Nutr. 1981, 1, 351–374. [Google Scholar] [CrossRef]
- Cooper, C.M.; Davies, N.W.; Menary, R.C. C-27 apocarotenoids in the flowers of Boronia megastigma (Nees). J. Agric. Food Chem. 2003, 51, 2384–2389. [Google Scholar] [CrossRef]
- Simkin, A.J.; Underwood, B.A.; Auldridge, M.; Loucas, H.M.; Shibuya, K.; Schmelz, E.; Clark, D.G.; Klee, H.J. Circadian regulation of the PhCCD1 carotenoid cleavage dioxygenase controls emission of beta-ionone, a fragrance volatile of petunia flowers. Plant Physiol. 2004, 136, 3504–3514. [Google Scholar] [CrossRef]
- Ansari, M.; Emami, S. Beta-Ionone and its analogs as promising anticancer agents. Eur. J. Med. Chem. 2016, 123, 141–154. [Google Scholar] [CrossRef]
- Elson, C.E.; Yu, S.G. The chemoprevention of cancer by mevalonate-derived constituents of fruits and vegetables. J. Nutr. 1994, 124, 607–614. [Google Scholar] [CrossRef]
- Liu, Q.; Dong, H.W.; Sun, W.G.; Liu, M.; Ibla, J.C.; Liu, L.X.; Parry, J.W.; Han, X.H.; Li, M.S.; Liu, J.R. Apoptosis initiation of beta-ionone in SGC-7901 gastric carcinoma cancer cells via a PI3K-AKT pathway. Arch. Toxicol. 2013, 87, 481–490. [Google Scholar] [CrossRef]
- Lalko, J.; Lapczynski, A.; McGinty, D.; Bhatia, S.; Letizia, C.S.; Api, A.M. Fragrance material review on beta-ionone. Food Chem. Toxicol. 2007, 45 (Suppl. 1), S241–S247. [Google Scholar] [CrossRef]
- Lalko, J.; Lapczynski, A.; Politano, V.T.; McGinty, D.; Bhatia, S.; Letizia, C.S.; Api, A.M. Fragrance material review on alpha-ionone. Food Chem. Toxicol. 2007, 45 (Suppl. 1), S235–S240. [Google Scholar] [CrossRef]
- O’Neil, M.J. The Merck index: An encyclopedia of chemicals, drugs, and biologicals. In Encyclopedia of Chemicals, Drugs, and Biologicals, 15th ed.; Royal Society of Chemistry: Cambridge, UK, 2013; p. 940. [Google Scholar]
- Brenna, E.; Fuganti, C.; Serra, S.; Kraft, P. Optically Active Ionones and Derivatives: Preparation and Olfactory Properties. Eur. J. Org. Chem. 2002, 2002, 967–978. [Google Scholar] [CrossRef]
- Terpenoids, S.C. Kirk-Othmer Encyclopedia of Chemical Technology, 4th ed.; Kirk-Othmer, Ed.; John Wiley & Sons: New York, NY, USA, 2007. [Google Scholar]
- Showing Compound Alpha-Ionone (FDB014484). Available online: https://foodb.ca/compounds/FDB014484 (accessed on 10 November 2020).
- Lide, D.R.; Milne, G.W.A. Handbook of Data on Organic Compounds, 3rd ed.; CRC Press: Boca Raton, FL, USA, 1994; Volume I. [Google Scholar]
- Etzweiler, F.; Senn, E.; Schmidt, H.W.H. Method for measuring aqueous solubilities of organic compounds. Anal. Chem. 1995, 67, 655–658. [Google Scholar] [CrossRef]
- Fichan, I.; Larroche, C.; Gros, J.B. Water solubility, vapor pressure, and activity coefficients of terpenes and terpenoids. J. Chem. Eng. Data 1999, 44, 56–62. [Google Scholar] [CrossRef]
- Von Soden, H. Über ätherische Öle, welche durch Extraktion frischer Blüten mit flüchtigen Lösungsmitteln gewonnen werden (ätherische Blütenextraktöle). J. Für Prakt. Chem. 1904, 69, 256–272. [Google Scholar] [CrossRef]
- Gautschi, M.; Bajgrowicz, J.A.; Kraft, P. Fragrance chemistry—milestones and perspectives. CHIMIA Int. J. Chem. 2001, 55, 379–387. [Google Scholar]
- Tiemann, F.; Krüger, P. Ueber veilchenaroma. Ber. Der Dtsch. Chem. Ges. 1893, 26, 2675–2708. [Google Scholar] [CrossRef]
- Krishna, H.J.; Joshi, B. Notes-Note on preparation of β-Ionone. J. Org. Chem. 1957, 22, 224–226. [Google Scholar] [CrossRef]
- Cataldo, V.F.; López, J.; Cárcamo, M.; Agosin, E. Chemical vs. biotechnological synthesis of C13-apocarotenoids: Current methods, applications and perspectives. Appl. Microbiol. Biotechnol. 2016, 100, 5703–5718. [Google Scholar] [CrossRef]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef]
- Romanchik, J.E.; Morel, D.W.; Harrison, E.H. Distributions of carotenoids and α-tocopherol among lipoproteins do not change when human plasma is incubated in vitro. J. Nutr. 1995, 125, 2610–2617. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.A.; Hayaishi, O. The enzymatic cleavage of beta-carotene into vitamin A by soluble enzymes of rat liver and intestine. Proc. Natl. Acad. Sci. USA 1965, 54, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, G.M.; Bornemann, C.; Giger, A.; Müller, R.K.; Schneider, H.; Schlotterbeck, G.; Schiefer, G.; Woggon, W.-D. The Substrate Specificity of β,β-Carotene 15,15′-Monooxygenase. Helv. Chim. Acta 2001, 84, 2301–2315. [Google Scholar] [CrossRef]
- Lindqvist, A.; Andersson, S. Biochemical properties of purified recombinant human beta-carotene 15,15′-monooxygenase. J. Biol. Chem. 2002, 277, 23942–23948. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, C.; Hessel, S.; Lampert, J.M.; Vogt, K.; Lederer, M.O.; Breithaupt, D.E.; von Lintig, J. Identification and characterization of a mammalian enzyme catalyzing the asymmetric oxidative cleavage of provitamin A. J. Biol. Chem. 2001, 276, 14110–14116. [Google Scholar] [CrossRef] [PubMed]
- Petroianu, G.A. Endogenous ionone. Commentary on “Study on the developmental toxicity of β-ionone in the rat”. Regul. Toxicol. Pharmacol. 2019, 101, 194–195. [Google Scholar] [CrossRef]
- Amengual, J.; Lobo, G.P.; Golczak, M.; Li, H.N.; Klimova, T.; Hoppel, C.L.; Wyss, A.; Palczewski, K.; von Lintig, J. A mitochondrial enzyme degrades carotenoids and protects against oxidative stress. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Guo, X.; Wang, W.; Medeiros, D.M.; Clarke, S.L.; Lucas, E.A.; Smith, B.J.; Lin, D. Molecular aspects of β, β-carotene-9′, 10′-oxygenase 2 in carotenoid metabolism and diseases. Exp. Biol. Med. 2016, 241, 1879–1887. [Google Scholar] [CrossRef]
- Palczewski, G.; Amengual, J.; Hoppel, C.L.; von Lintig, J. Evidence for compartmentalization of mammalian carotenoid metabolism. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 4457–4469. [Google Scholar] [CrossRef]
- Wang, X.D.; Russell, R.M.; Liu, C.; Stickel, F.; Smith, D.E.; Krinsky, N.I. Beta-oxidation in rabbit liver in vitro and in the perfused ferret liver contributes to retinoic acid biosynthesis from beta-apocarotenoic acids. J. Biol. Chem. 1996, 271, 26490–26498. [Google Scholar] [CrossRef]
- Hessel, S.; Eichinger, A.; Isken, A.; Amengual, J.; Hunzelmann, S.; Hoeller, U.; Elste, V.; Hunziker, W.; Goralczyk, R.; Oberhauser, V.; et al. CMO1 deficiency abolishes vitamin A production from beta-carotene and alters lipid metabolism in mice. J. Biol. Chem. 2007, 282, 33553–33561. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, A.; He, Y.G.; Andersson, S. Cell type-specific expression of beta-carotene 9’,10’-monooxygenase in human tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2005, 53, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Jovancevic, N.; Khalfaoui, S.; Weinrich, M.; Weidinger, D.; Simon, A.; Kalbe, B.; Kernt, M.; Kampik, A.; Gisselmann, G.; Gelis, L.; et al. Odorant receptor 51E2 agonist β-ionone regulates RPE cell migration and proliferation. Front. Physiol. 2017, 8, 888. [Google Scholar] [CrossRef] [PubMed]
- Våge, D.I.; Boman, I.A. A nonsense mutation in the beta-carotene oxygenase 2 (BCO2) gene is tightly associated with accumulation of carotenoids in adipose tissue in sheep (Ovis aries). BMC Genet. 2010, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Strychalski, J.; Gugołek, A.; Brym, P.; Antoszkiewicz, Z.; Iwona, C.-S. Polymorphism of the BCO2 gene and the content of carotenoids, retinol, and α-tocopherol in the liver and fat of rabbits. Rev. Bras. Zootec. 2019, 48. [Google Scholar] [CrossRef]
- Lobo, G.P.; Isken, A.; Hoff, S.; Babino, D.; von Lintig, J. BCDO2 acts as a carotenoid scavenger and gatekeeper for the mitochondrial apoptotic pathway. Development 2012, 139, 2966–2977. [Google Scholar] [CrossRef]
- Tatman, D.; Mo, H. Volatile isoprenoid constituents of fruits, vegetables and herbs cumulatively suppress the proliferation of murine B16 melanoma and human HL-60 leukemia cells. Cancer Lett. 2002, 175, 129–139. [Google Scholar] [CrossRef]
- Lobo, G.P.; Amengual, J.; Palczewski, G.; Babino, D.; von Lintig, J. Mammalian carotenoid-oxygenases: Key players for carotenoid function and homeostasis. Biochim. Biophys. Acta 2012, 1821, 78–87. [Google Scholar] [CrossRef]
- Al-Tel, T.; Tarazi, H.; Aloum, L.; Lorke, D.; Petroianu, G. Possible metabolic conversion of pinene to ionone. Die Pharm. 2020, 75, 360–363. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Maßberg, D.; Hatt, H. Human olfactory receptors: Novel cellular functions outside of the nose. Physiol. Rev. 2018, 98, 1739–1763. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Reed, R.R. Golf: An olfactory neuron specific-G protein involved in odorant signal transduction. Science 1989, 244, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Sklar, P.B.; Anholt, R.R.; Snyder, S.H. The odorant-sensitive adenylate cyclase of olfactory receptor cells. Differential stimulation by distinct classes of odorants. J. Biol. Chem. 1986, 261, 15538–15543. [Google Scholar] [PubMed]
- Dhallan, R.S.; Yau, K.W.; Schrader, K.A.; Reed, R.R. Primary structure and functional expression of a cyclic nucleotide-activated channel from olfactory neurons. Nature 1990, 347, 184–187. [Google Scholar] [CrossRef]
- Gelis, L.; Jovancevic, N.; Veitinger, S.; Mandal, B.; Arndt, H.D.; Neuhaus, E.M.; Hatt, H. Functional characterization of the odorant receptor 51e2 in human melanocytes. J. Biol. Chem. 2016, 291, 17772–17786. [Google Scholar] [CrossRef]
- Xie, H.; Liu, T.; Chen, J.; Yang, Z.; Xu, S.; Fan, Y.; Zeng, J.; Chen, Y.; Ma, Z.; Gao, Y.; et al. Activation of PSGR with β-ionone suppresses prostate cancer progression by blocking androgen receptor nuclear translocation. Cancer Lett. 2019, 453, 193–205. [Google Scholar] [CrossRef]
- Parmentier, M.; Libert, F.; Schurmans, S.; Schiffmann, S.; Lefort, A.; Eggerickx, D.; Ledent, C.; Mollereau, C.; Gérard, C.; Perret, J.; et al. Expression of members of the putative olfactory receptor gene family in mammalian germ cells. Nature 1992, 355, 453–455. [Google Scholar] [CrossRef]
- Feldmesser, E.; Olender, T.; Khen, M.; Yanai, I.; Ophir, R.; Lancet, D. Widespread ectopic expression of olfactory receptor genes. BMC Genom. 2006, 7, 121. [Google Scholar] [CrossRef]
- De la Cruz, O.; Blekhman, R.; Zhang, X.; Nicolae, D.; Firestein, S.; Gilad, Y. A signature of evolutionary constraint on a subset of ectopically expressed olfactory receptor genes. Mol. Biol. Evol. 2009, 26, 491–494. [Google Scholar] [CrossRef]
- Flegel, C.; Manteniotis, S.; Osthold, S.; Hatt, H.; Gisselmann, G. Expression profile of ectopic olfactory receptors determined by deep sequencing. PLoS ONE 2013, 8, e55368. [Google Scholar] [CrossRef]
- Braun, T.; Voland, P.; Kunz, L.; Prinz, C.; Gratzl, M. Enterochromaffin cells of the human gut: Sensors for spices and odorants. Gastroenterology 2007, 132, 1890–1901. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, E.M.; Zhang, W.; Gelis, L.; Deng, Y.; Noldus, J.; Hatt, H. Activation of an olfactory receptor inhibits proliferation of prostate cancer cells. J. Biol. Chem. 2009, 284, 16218–16225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bedigian, A.V.; Wang, W.; Eggert, U.S. G protein-coupled receptors participate in cytokinesis. Cytoskeleton 2012, 69, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Zou, D.J.; Zhang, X.; Yan, Q.; Rodriguez-Gil, D.J.; Eisner, C.; Wells, E.; Greer, C.A.; Wang, T.; Firestein, S.; et al. Functional expression of the olfactory signaling system in the kidney. Proc. Natl. Acad. Sci. USA 2009, 106, 2059–2064. [Google Scholar] [CrossRef]
- Fukuda, N.; Touhara, K. Developmental expression patterns of testicular olfactory receptor genes during mouse spermatogenesis. Genes Cells Devoted Mol. Cell. Mech. 2006, 11, 71–81. [Google Scholar] [CrossRef]
- Fukuda, N.; Yomogida, K.; Okabe, M.; Touhara, K. Functional characterization of a mouse testicular olfactory receptor and its role in chemosensing and in regulation of sperm motility. J. Cell Sci. 2004, 117, 5835–5845. [Google Scholar] [CrossRef]
- Spehr, M.; Gisselmann, G.; Poplawski, A.; Riffell, J.A.; Wetzel, C.H.; Zimmer, R.K.; Hatt, H. Identification of a testicular odorant receptor mediating human sperm chemotaxis. Science 2003, 299, 2054–2058. [Google Scholar] [CrossRef]
- Spehr, M.; Schwane, K.; Heilmann, S.; Gisselmann, G.; Hummel, T.; Hatt, H. Dual capacity of a human olfactory receptor. Curr. Biol. CB 2004, 14, R832–R833. [Google Scholar] [CrossRef]
- Spehr, J.; Gelis, L.; Osterloh, M.; Oberland, S.; Hatt, H.; Spehr, M.; Neuhaus, E.M. G protein-coupled receptor signaling via Src kinase induces endogenous human transient receptor potential vanilloid type 6 (TRPV6) channel activation. J. Biol. Chem. 2011, 286, 13184–13192. [Google Scholar] [CrossRef]
- Sanz, G.; Leray, I.; Dewaele, A.; Sobilo, J.; Lerondel, S.; Bouet, S.; Grébert, D.; Monnerie, R.; Pajot-Augy, E.; Mir, L.M. Promotion of cancer cell invasiveness and metastasis emergence caused by olfactory receptor stimulation. PLoS ONE 2014, 9, e85110. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Li, F.; Yao, J.; Yu, J. Prostate specific G protein coupled receptor is associated with prostate cancer prognosis and affects cancer cell proliferation and invasion. BMC Cancer 2015, 15, 915. [Google Scholar] [CrossRef] [PubMed]
- Wiese, H.; Gelis, L.; Wiese, S.; Reichenbach, C.; Jovancevic, N.; Osterloh, M.; Meyer, H.E.; Neuhaus, E.M.; Hatt, H.H.; Radziwill, G.; et al. Quantitative phosphoproteomics reveals the protein tyrosine kinase Pyk2 as a central effector of olfactory receptor signaling in prostate cancer cells. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2015, 1854, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Gelis, L.; Jovancevic, N.; Bechara, F.G.; Neuhaus, E.M.; Hatt, H. Functional expression of olfactory receptors in human primary melanoma and melanoma metastasis. Exp. Dermatol. 2017, 26, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Stackhouse, B.G.; Florence, K.; Zhang, W.; Shanmugam, N.; Sesterhenn, I.A.; Zou, Z.; Srikantan, V.; Augustus, M.; Roschke, V.; et al. PSGR, a novel prostate-specific gene with homology to a G protein-coupled receptor, is overexpressed in prostate cancer. Cancer Res. 2000, 60, 6568–6572. [Google Scholar] [PubMed]
- Sanz, G.; Leray, I.; Grébert, D.; Antoine, S.; Acquistapace, A.; Muscat, A.; Boukadiri, A.; Mir, L.M. Structurally related odorant ligands of the olfactory receptor OR51E2 differentially promote metastasis emergence and tumor growth. Oncotarget 2017, 8, 4330–4341. [Google Scholar] [CrossRef] [PubMed]
- Maßberg, D.; Jovancevic, N.; Offermann, A.; Simon, A.; Baniahmad, A.; Perner, S.; Pungsrinont, T.; Luko, K.; Philippou, S.; Ubrig, B.; et al. The activation of OR51E1 causes growth suppression of human prostate cancer cells. Oncotarget 2016, 7, 48231–48249. [Google Scholar] [CrossRef]
- Duncan, R.E.; Lau, D.; El-Sohemy, A.; Archer, M.C. Geraniol and beta-ionone inhibit proliferation, cell cycle progression, and cyclin-dependent kinase 2 activity in MCF-7 breast cancer cells independent of effects on HMG-CoA reductase activity. Biochem. Pharmacol. 2004, 68, 1739–1747. [Google Scholar] [CrossRef]
- Dong, H.W.; Wang, K.; Chang, X.X.; Jin, F.F.; Wang, Q.; Jiang, X.F.; Liu, J.R.; Wu, Y.H.; Yang, C. Beta-ionone-inhibited proliferation of breast cancer cells by inhibited COX-2 activity. Arch. Toxicol. 2019, 93, 2993–3003. [Google Scholar] [CrossRef]
- Jones, S.; Fernandes, N.V.; Yeganehjoo, H.; Katuru, R.; Qu, H.; Yu, Z.; Mo, H. β-ionone induces cell cycle arrest and apoptosis in human prostate tumor cells. Nutr. Cancer 2013, 65, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Cooma, I.; Mohammed, A.; Steele, V.E.; Rao, C.V. Beta-ionone inhibits colonic aberrant crypt foci formation in rats, suppresses cell growth, and induces retinoid X receptor-alpha in human colon cancer cells. Mol. Cancer Ther. 2008, 7, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-R.; Sun, X.-R.; Dong, H.-W.; Sun, C.-H.; Sun, W.-G.; Chen, B.-Q.; Song, Y.-Q.; Yang, B.-F. β-Ionone suppresses mammary carcinogenesis, proliferative activity and induces apoptosis in the mammary gland of the Sprague-Dawley rat. Int. J. Cancer 2008, 122, 2689–2698. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.R.; Chen, B.Q.; Yang, B.F.; Dong, H.W.; Sun, C.H.; Wang, Q.; Song, G.; Song, Y.Q. Apoptosis of human gastric adenocarcinoma cells induced by beta-ionone. World J. Gastroenterol. 2004, 10, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liu, J.; Sun, W.; Tang, X. Beta-ionone induced apoptosis in SGC-7901 cells. J. Hyg. Res. 2007, 36, 667–670. [Google Scholar]
- Dong, H.W.; Zhang, S.; Sun, W.G.; Liu, Q.; Ibla, J.C.; Soriano, S.G.; Han, X.H.; Liu, L.X.; Li, M.S.; Liu, J.R. β-Ionone arrests cell cycle of gastric carcinoma cancer cells by a MAPK pathway. Arch. Toxicol. 2013, 87, 1797–1808. [Google Scholar] [CrossRef]
- Yu, S.G.; Abuirmeileh, N.M.; Qureshi, A.A.; Elson, C.E. Dietary beta.-ionone suppresses hepatic 3-hydroxy-3-methylglutaryl coenzyme A reductase activity. J. Agric. Food Chem. 1994, 42, 1493–1496. [Google Scholar] [CrossRef]
- Case, G.L.; He, L.; Mo, H.; Elson, C.E. Induction of geranyl pyrophosphate pyrophosphatase activity by cholesterol-suppressive isoprenoids. Lipids 1995, 30, 357–359. [Google Scholar] [CrossRef]
- De Moura Espíndola, R.; Mazzantini, R.P.; Ong, T.P.; de Conti, A.; Heidor, R.; Moreno, F.S. Geranylgeraniol and beta-ionone inhibit hepatic preneoplastic lesions, cell proliferation, total plasma cholesterol and DNA damage during the initial phases of hepatocarcinogenesis, but only the former inhibits NF-kappaB activation. Carcinogenesis 2005, 26, 1091–1099. [Google Scholar] [CrossRef]
- Kang, C.H.; Jayasooriya, R.G.; Choi, Y.H.; Moon, S.K.; Kim, W.J.; Kim, G.Y. β-Ionone attenuates LPS-induced pro-inflammatory mediators such as NO, PGE2 and TNF-α in BV2 microglial cells via suppression of the NF-κB and MAPK pathway. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2013, 27, 782–787. [Google Scholar] [CrossRef]
- Liu, J.R.; Yang, Y.M.; Dong, H.W.; Sun, X.R. Effect of beta-ionone in human mammary cancer cells (Er-) by MAPK pathway. J. Hyg. Res. 2005, 34, 706–709. [Google Scholar]
- Huang, C.S.; Lyu, S.C.; Hu, M.L. Synergistic effects of the combination of β-ionone and sorafenib on metastasis of human hepatoma SK-Hep-1 cells. Investig. New Drugs 2012, 30, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.R.; Yang, B.F.; Chen, B.Q.; Yang, Y.M.; Dong, H.W.; Song, Y.Q. Inhibition of beta-ionone on SGC-7901 cell proliferation and upregulation of metalloproteinases-1 and -2 expression. World J. Gastroenterol. 2004, 10, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Mescher, A. Junqueira’s Basic Histology Text & Atlas, 14th ed.; McGraw-Hill Education: New York, NY, USA, 2016. [Google Scholar]
- Scolastici, C.; de Conti, A.; Cardozo, M.T.; Ong, T.P.; Purgatto, E.; Horst, M.A.; Heidor, R.; Furtado, K.S.; Bassoli, B.K.; Moreno, F.S. β-ionone inhibits persistent preneoplastic lesions during the early promotion phase of rat hepatocarcinogenesis: TGF-α, NF-κB, and p53 as cellular targets. Nutr. Cancer 2014, 66, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef] [PubMed]
- Abbastabar, M.; Kheyrollah, M.; Azizian, K.; Bagherlou, N.; Tehrani, S.S.; Maniati, M.; Karimian, A. Multiple functions of p27 in cell cycle, apoptosis, epigenetic modification and transcriptional regulation for the control of cell growth: A double-edged sword protein. DNA Repair 2018, 69, 63–72. [Google Scholar] [CrossRef]
- Kim, M.O.; Moon, D.O.; Kang, C.H.; Kwon, T.K.; Choi, Y.H.; Kim, G.Y. beta-Ionone enhances TRAIL-induced apoptosis in hepatocellular carcinoma cells through Sp1-dependent upregulation of DR5 and downregulation of NF-kappaB activity. Mol. Cancer Ther. 2010, 9, 833–843. [Google Scholar] [CrossRef]
- Mao, G.E.; Reuter, V.E.; Cordon-Cardo, C.; Dalbagni, G.; Scher, H.I.; DeKernion, J.B.; Zhang, Z.F.; Rao, J. Decreased retinoid X receptor-alpha protein expression in basal cells occurs in the early stage of human prostate cancer development. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2004, 13, 383–390. [Google Scholar]
- Vieira, A.; Heidor, R.; Cardozo, M.T.; Scolastici, C.; Purgatto, E.; Shiga, T.M.; Barbisan, L.F.; Ong, T.P.; Moreno, F.S. Efficacy of geraniol but not of β-ionone or their combination for the chemoprevention of rat colon carcinogenesis. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 2011, 44, 538–545. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nature 1990, 343, 425–430. [Google Scholar] [CrossRef]
- Wang, M.; Casey, P.J. Protein prenylation: Unique fats make their mark on biology. Nat. Rev. Mol. Cell Biol. 2016, 17, 110–122. [Google Scholar] [CrossRef]
- Mo, H.; Jeter, R.; Bachmann, A.; Yount, S.T.; Shen, C.L.; Yeganehjoo, H. The potential of isoprenoids in adjuvant cancer therapy to reduce adverse effects of statins. Front. Pharmacol. 2018, 9, 1515. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Kim, H.S.; Kim, J.H.; Lee, J. The Effect of statin added to systemic anticancer therapy: A meta-analysis of randomized, controlled trials. J. Clin. Med. 2018, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, M.A.M.; Malhotra, N.; Mukherjee, S.D.; Sanger, S.; Dhesy-Thind, S.K.; Ellis, P.; Leong, D.P. Statin therapy in the treatment of active cancer: A systematic review and meta-analysis of randomized controlled trials. PLoS ONE 2018, 13, e0209486. [Google Scholar] [CrossRef] [PubMed]
- Elson, C.E.; Peffley, D.M.; Hentosh, P.; Mo, H. Isoprenoid-mediated inhibition of mevalonate synthesis: Potential application to cancer. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1999, 221, 294–311. [Google Scholar] [CrossRef]
- Mo, H.; Elson, C.E. Studies of the isoprenoid-mediated inhibition of mevalonate synthesis applied to cancer chemotherapy and chemoprevention. Exp. Biol. Med. 2004, 229, 567–585. [Google Scholar] [CrossRef]
- Elson, C.E. Suppression of mevalonate pathway activities by dietary isoprenoids: Protective roles in cancer and cardiovascular disease. J. Nutr. 1995, 125, 1666s–1672s. [Google Scholar] [CrossRef]
- Panini, S.; Schnitzer-Polokoff, R.; Spencer, T.; Sinensky, M. Sterol-independent regulation of 3-hydroxy-3-methylglutaryl-CoA reductase by mevalonate in Chinese hamster ovary cells. Magnitude and specificity. J. Biol. Chem. 1989, 264, 11044–11052. [Google Scholar]
- Bennis, F.; Favre, G.; Le Gaillard, F.; Soula, G. Importance of mevalonate-derived products in the control of HMG-CoA reductase activity and growth of human lung adenocarcinoma cell line A549. Int. J. Cancer 1993, 55, 640–645. [Google Scholar] [CrossRef]
- Kawata, S.; Takaishi, K.; Nagase, T.; Ito, N.; Matsuda, Y.; Tamura, S.; Matsuzawa, Y.; Tarui, S. Increase in the active form of 3-hydroxy-3-methylglutaryl coenzyme A reductase in human hepatocellular carcinoma: Possible mechanism for alteration of cholesterol biosynthesis. Cancer Res. 1990, 50, 3270–3273. [Google Scholar]
- El-Sohemy, A.; Archer, M.C. Inhibition of N-methyl-N-nitrosourea- and 7,12-dimethylbenz[a] anthracene-induced rat mammary tumorigenesis by dietary cholesterol is independent of Ha-Ras mutations. Carcinogenesis 2000, 21, 827–831. [Google Scholar] [CrossRef][Green Version]
- Harwood, H.J., Jr.; Alvarez, I.M.; Noyes, W.D.; Stacpoole, P.W. In vivo regulation of human leukocyte 3-hydroxy-3-methylglutaryl coenzyme A reductase: Increased enzyme protein concentration and catalytic efficiency in human leukemia and lymphoma. J. Lipid Res. 1991, 32, 1237–1252. [Google Scholar] [PubMed]
- Caruso, M.G.; Notarnicola, M.; Cavallini, A.; Di Leo, A. 3-Hydroxy-3-methylglutaryl coenzyme A reductase activity and low-density lipoprotein receptor expression in diffuse-type and intestinal-type human gastric cancer. J. Gastroenterol. 2002, 37, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Moreno, F.S.; Rossiello, M.R.; Manjeshwar, S.; Nath, R.; Rao, P.M.; Rajalakshmi, S.; Sarma, D.S. Effect of beta-carotene on the expression of 3-hydroxy-3-methylglutaryl coenzyme A reductase in rat liver. Cancer Lett. 1995, 96, 201–208. [Google Scholar] [CrossRef]
- Yu, S.G.; Anderson, P.J.; Elson, C.E. Efficacy of beta.-Ionone in the Chemoprevention of Rat Mammary Carcinogenesis. J. Agric. Food Chem. 1995, 43, 2144–2147. [Google Scholar] [CrossRef]
- Liu, J.R.; Dong, H.W.; Sun, X.R.; Wang, Q.; Sun, W.G.; Parry, J.W.; Liu, Q.; Han, X.H.; Sun, C.H.; Chen, B.Q.; et al. Effects of beta-ionone on mammary carcinogenesis and antioxidant status in rats treated with DMBA. Nutr. Cancer 2010, 62, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Correll, C.C.; Ng, L.; Edwards, P.A. Identification of farnesol as the non-sterol derivative of mevalonic acid required for the accelerated degradation of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Biol. Chem. 1994, 269, 17390–17393. [Google Scholar] [PubMed]
- McFarland, A.J.; Anoopkumar-Dukie, S.; Arora, D.S.; Grant, G.D.; McDermott, C.M.; Perkins, A.V.; Davey, A.K. Molecular mechanisms underlying the effects of statins in the central nervous system. Int. J. Mol. Sci. 2014, 15, 20607–20637. [Google Scholar] [CrossRef]
- Perchellet, J.-P.H.; Perchellet, E.M.; Crow, K.R.; Buszek, K.R.; Brown, N.; Ellappan, S.; Gao, G.; Luo, D.; Minatoya, M.; Lushington, G.H. Novel synthetic inhibitors of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase activity that inhibit tumor cell proliferation and are structurally unrelated to existing statins. Int. J. Mol. Med. 2009, 24, 633–643. [Google Scholar] [CrossRef]
- Cardozo, M.T.; de Conti, A.; Ong, T.P.; Scolastici, C.; Purgatto, E.; Horst, M.A.; Bassoli, B.K.; Moreno, F.S. Chemopreventive effects of β-ionone and geraniol during rat hepatocarcinogenesis promotion: Distinct actions on cell proliferation, apoptosis, HMGCoA reductase, and RhoA. J. Nutr. Biochem. 2011, 22, 130–135. [Google Scholar] [CrossRef]
- Ong, T.P.; Heidor, R.; de Conti, A.; Dagli, M.L.; Moreno, F.S. Farnesol and geraniol chemopreventive activities during the initial phases of hepatocarcinogenesis involve similar actions on cell proliferation and DNA damage, but distinct actions on apoptosis, plasma cholesterol and HMGCoA reductase. Carcinogenesis 2006, 27, 1194–1203. [Google Scholar] [CrossRef]
- Asokkumar, S.; Naveenkumar, C.; Raghunandhakumar, S.; Kamaraj, S.; Anandakumar, P.; Jagan, S.; Devaki, T. Antiproliferative and antioxidant potential of beta-ionone against benzo(a)pyrene-induced lung carcinogenesis in Swiss albino mice. Mol. Cell. Biochem. 2012, 363, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Lei, W.; Mandlekar, S.; Weber, M.; Der, C.; Wu, J.; Kong, A. Role of a Mitogen-activated protein kinase pathway in the induction of phase II detoxifying enzymes by chemicals. J. Biol. Chem. 1999, 274, 27545–27552. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.G.; Wyllie, S.G.; Markham, J.L.; Leach, D.N. The role of structure and molecular properties of terpenoids in determining their antimicrobial activity. Flavour Fragr. J. 1999, 14, 322–332. [Google Scholar] [CrossRef]
- Wilson, D.M.; Gueldner, R.C.; McKinney, J.K.; Lievsay, R.H.; Evans, B.D.; Hill, R.A. Effect of β-lonone onaspergillus flavus andaspergillus parasiticus growth, sporulation, morphology and aflatoxin production. J. Am. Oil Chem. Soc. 1981, 58, A959–A961. [Google Scholar] [CrossRef]
- Luttrell, L.M.; Maudsley, S.; Bohn, L.M. Fulfilling the Promise of “Biased“ G Protein-Coupled Receptor Agonism. Mol. Pharmacol. 2015, 88, 579–588. [Google Scholar] [CrossRef]
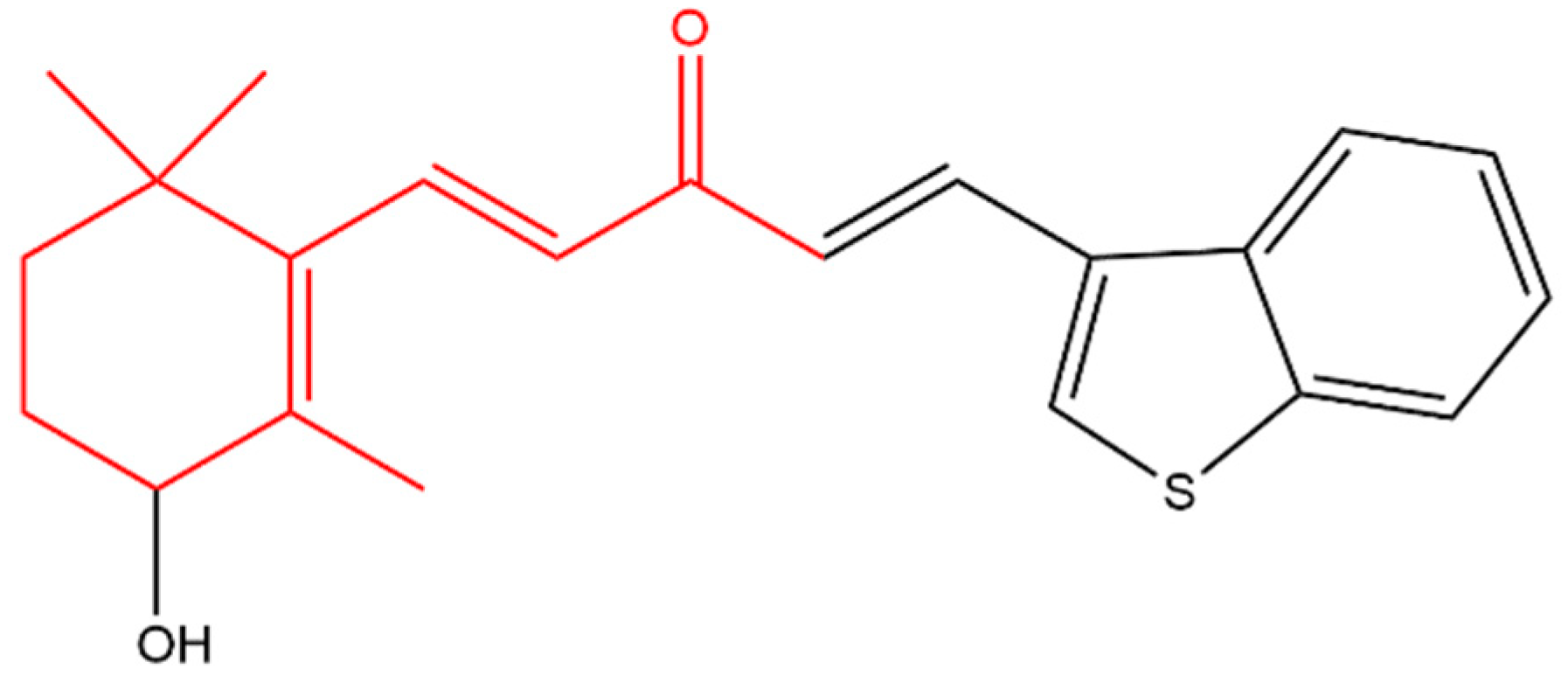
- Luetragoon, T.; Pankla Sranujit, R.; Noysang, C.; Thongsri, Y.; Potup, P.; Suphrom, N.; Nuengchamnong, N.; Usuwanthim, K. Anti-Cancer effect of 3-Hydroxy-β-Ionone identified from moringa oleifera Lam. Leaf on human squamous cell carcinoma 15 cell line. Molecules 2020, 25, 3563. [Google Scholar] [CrossRef]
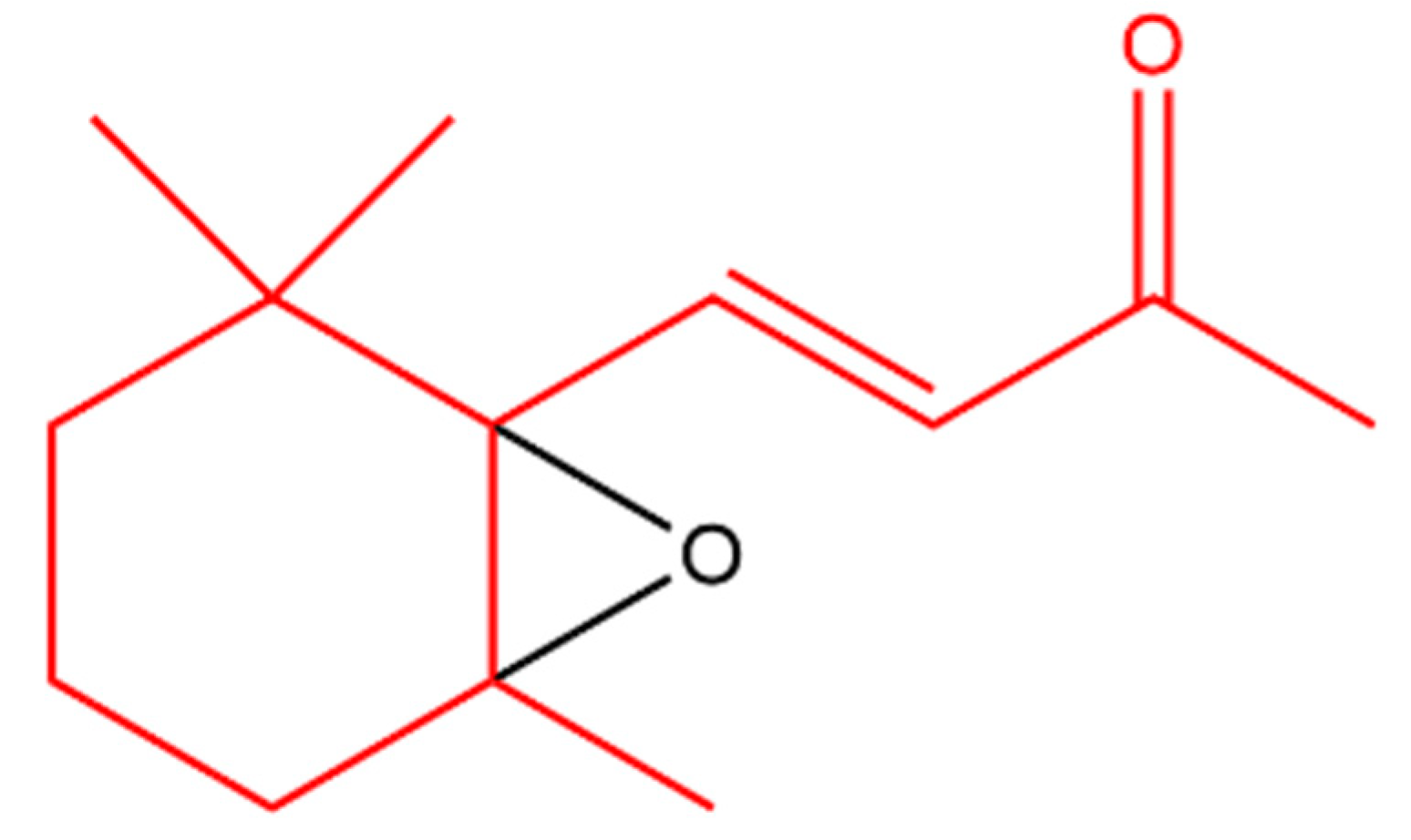
- Wertz, P.W.; Kensler, T.W.; Mueller, G.C. Inhibition of phorbol ester action in lymphocytes by 5,6-epoxy-β-ionone. Biochem. Biophys. Res. Commun. 1978, 83, 138–143. [Google Scholar] [CrossRef]
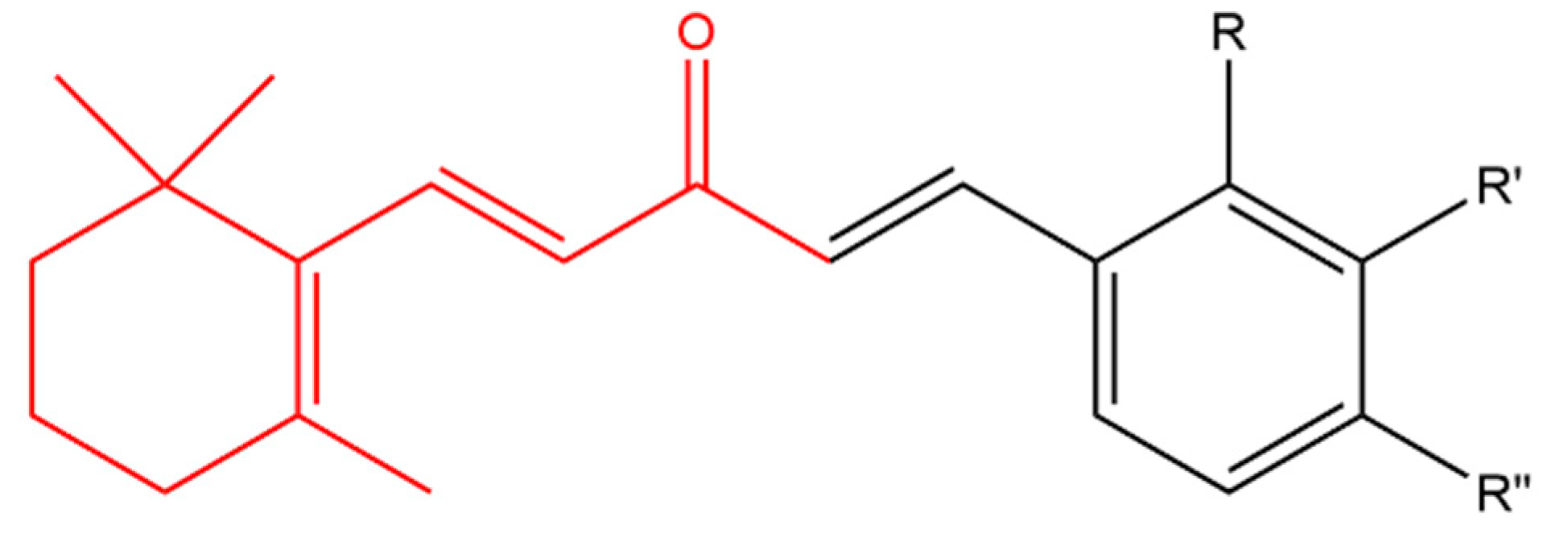
- Sharma, V.; Chaudhary, A.; Arora, S.; Saxena, A.K.; Ishar, M.P.S. β-Ionone derived chalcones as potent antiproliferative agents. Eur. J. Med. Chem. 2013, 69, 310–315. [Google Scholar] [CrossRef]
- Zhou, J.; Geng, G.; Batist, G.; Wu, J.H. Syntheses and potential anti-prostate cancer activities of ionone-based chalcones. Bioorganic Med. Chem. Lett. 2009, 19, 1183–1186. [Google Scholar] [CrossRef]
- Zhou, J.; Geng, G.; Shi, Q.; Sauriol, F.; Wu, J.H. Design and synthesis of androgen receptor antagonists with bulky side chains for overcoming antiandrogen resistance. J. Med. Chem. 2009, 52, 5546–5550. [Google Scholar] [CrossRef]
- Zhou, J.; Geng, G.; Wu, J.H. Synthesis and in vitro characterization of ionone-based chalcones as novel antiandrogens effective against multiple clinically relevant androgen receptor mutants. Investig. New Drugs 2010, 28, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, J.; Geng, G.; Lin, R.; Wu, J.H. Synthesis and in vitro characterization of ionone-based compounds as dual inhibitors of the androgen receptor and NF-κB. Investig. New Drugs 2014, 32, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Balbi, A.; Anzaldi, M.; Mazzei, M.; Miele, M.; Bertolotto, M.; Ottonello, L.; Dallegri, F. Synthesis and biological evaluation of novel heterocyclic ionone-like derivatives as anti-inflammatory agents. Bioorganic Med. Chem. 2006, 14, 5152–5160. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Cai, M.; Qi, D.; Ying, X.; Huang, C.; Xing, C. β-Ionone-Derived Curcumin Analogs as Potent Anti-Inflammatory Agents. Pharm. Chem. J. 2018, 51, 902–906. [Google Scholar] [CrossRef]
- Anzaldi, M.; Sottofattori, E.; Rizzetto, R.; Granello di Casaleto, B.; Balbi, A. Synthesis and antimicrobial activity of heterocyclic ionone-like derivatives. Eur. J. Med. Chem. 1999, 34, 837–842. [Google Scholar] [CrossRef]
- Sharma, V.; Singh, G.; Kaur, H.; Saxena, A.K.; Ishar, M.P. Synthesis of β-ionone derived chalcones as potent antimicrobial agents. Bioorganic Med. Chem. Lett. 2012, 22, 6343–6346. [Google Scholar] [CrossRef]
- Grabarczyk, M.; Wińska, K.; Mączka, W.; Żarowska, B.; Maciejewska, G.; Dancewicz, K.; Gabryś, B.; Anioł, M. Synthesis, biotransformation and biological activity of halolactones obtained from β-ionone. Tetrahedron 2016, 72, 637–644. [Google Scholar] [CrossRef]
- Suryawanshi, S.N.; Bhat, B.A.; Pandey, S.; Chandra, N.; Gupta, S. Chemotherapy of leishmaniasis. Part VII: Synthesis and bioevaluation of substituted terpenyl pyrimidines. Eur. J. Med. Chem. 2007, 42, 1211–1217. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-Ionone | β-Ionone | Reference | |
|---|---|---|---|
| Chemical structure depiction |  |  | [10,11] |
| β-Carotene Oxygenase 1 (BCO1) | β-Carotene Oxygenase 2 (BCO2) | |
|---|---|---|
| Site of Cleavage | Symmetric–central | Asymmetric–eccentric |
| Products of the cleavage of β-carotene | Two entities of all-trans retinal | β-ionone and β-apo-10′-carotenal |
| Substrate specificity | Pro-vitamin carotenoids | Pro-vitamin and non-pro-vitamin carotenoids |
| Cellular compartmentalization | Cytoplasm | Inner mitochondrial membrane |
| Human tissue expression | Tissues that are expressing BCO1 were also found to express BCO2 however only BCO2 was detected in endometrial connective tissue, endocrine pancreas and cardiac and skeletal muscle | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aloum, L.; Alefishat, E.; Adem, A.; Petroianu, G. Ionone Is More than a Violet’s Fragrance: A Review. Molecules 2020, 25, 5822. https://doi.org/10.3390/molecules25245822
Aloum L, Alefishat E, Adem A, Petroianu G. Ionone Is More than a Violet’s Fragrance: A Review. Molecules. 2020; 25(24):5822. https://doi.org/10.3390/molecules25245822
Chicago/Turabian StyleAloum, Lujain, Eman Alefishat, Abdu Adem, and Georg Petroianu. 2020. "Ionone Is More than a Violet’s Fragrance: A Review" Molecules 25, no. 24: 5822. https://doi.org/10.3390/molecules25245822
APA StyleAloum, L., Alefishat, E., Adem, A., & Petroianu, G. (2020). Ionone Is More than a Violet’s Fragrance: A Review. Molecules, 25(24), 5822. https://doi.org/10.3390/molecules25245822