
Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus

 , ,
, , 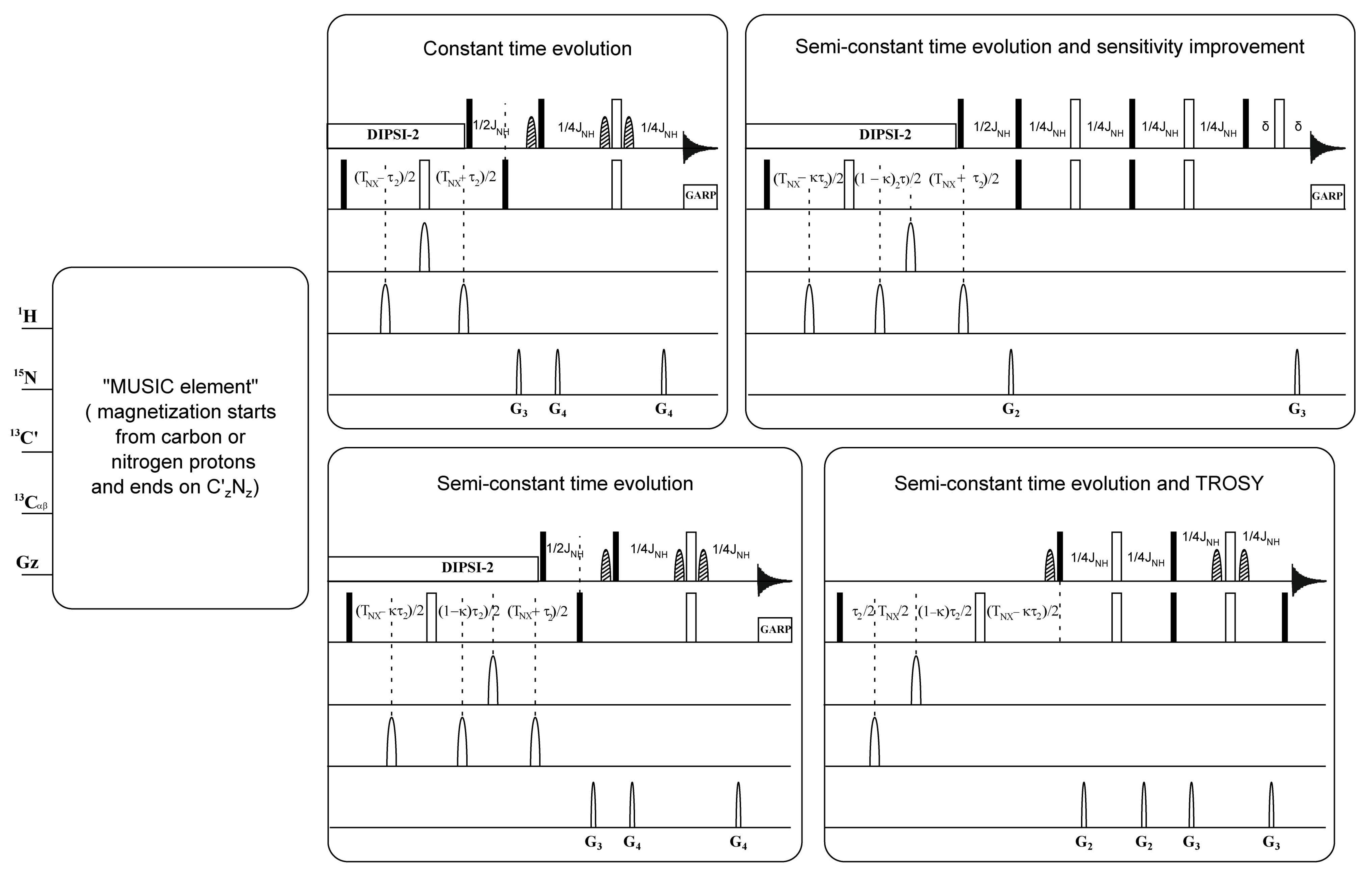{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Purification
2.2. NMR Spectroscopy
2.3. Preparation of Samples Used for NMR Assignment Experiments
2.4. NMR Experiments
2.5. MUSIC Experiments
3. Results
3.1. Selective Detection of Different Types of Amino Acids in vHVD + GdmCl Spectra
3.2. Proline Assignment
3.3. Assignment of the 1H and 15N in vHVD by Back-Titration
4. Discussion
4.1. Chemical Shift Assignment Strategy
4.2. Secondary Structure of Native vHDV
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jensen:, M.R.; Ruigrok, R.W.H.; Blackledge, M. Describing intrinsically disordered proteins at atomic resolution by NMR. Curr. Opin. Struct. Biol. 2013, 23, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Parigi, G.; Rezaei-Ghaleh, N.; Giachetti, A.; Becker, S.; Fernandez, C.; Blackledge, M.; Griesinger, C.; Zweckstetter, M.; Luchinat, C. Long-Range Correlated Dynamics in Intrinsically Disordered Proteins. J. Am. Chem. Soc. 2014, 136, 16201–16209. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Huang, J.R.; Yao, M.; Communie, G.; Ozenne, V.; Mollica, L.; Salmon, L.; Jensen, M.R.; Blackledge, M. Towards a robust description of intrinsic protein disorder using nuclear magnetic resonance spectroscopy. Mol. Biosyst. 2012, 8, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Felli, I.C.; Pierattelli, R. Novel methods based on C-13 detection to study intrinsically disordered proteins. J. Magn. Reson. 2014, 241, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.C.; Usher, G.A.; Showalter, S.A. The Use of C-13 Direct-Detect NMR to Characterize Flexible and Disordered Proteins. Method Enzym. 2018, 611, 81–100. [Google Scholar] [CrossRef]
- Gibbs, E.B.; Cook, E.C.; Showalter, S.A. Application of NMR to studies of intrinsically disordered proteins. Arch. Biochem. Biophys. 2017, 628, 57–70. [Google Scholar] [CrossRef]
- Bermel, W.; Bertini, I.; Felli, I.C.; Piccioli, M.; Pierattelli, R. C-13-detected protonless NMR spectroscopy of proteins in solution. Prog. Nucl. Mag. Res. Spectrosc. 2006, 48, 25–45. [Google Scholar] [CrossRef]
- Sahu, D.; Bastidas, M.; Showalter, S.A. Generating NMR chemical shift assignments of intrinsically disordered proteins using carbon-detected NMR methods. Anal. Biochem. 2014, 449, 17–25. [Google Scholar] [CrossRef]
- Schubert, M.; Labudde, D.; Leitner, D.; Oschkinat, H.; Schmieder, P. A modified strategy for sequence specific assignment of protein NMR spectra based on amino acid type selective experiments. J. Biomol. NMR 2005, 31, 115–127. [Google Scholar] [CrossRef]
- Sun, Z.Y.J.; Frueh, D.P.; Selenko, P.; Hoch, J.C.; Wagner, G. Fast assignment of N-15-HSQC peaks using high-resolution 3D HNcocaNH experiments with non-uniform sampling. J. Biomol. NMR 2005, 33, 43–50. [Google Scholar] [CrossRef]
- Pervushin, K.; Riek, R.; Wider, G.; Wuthrich, K. Attenuated T2 relaxation by mutual cancellation of dipole-dipole coupling and chemical shift anisotropy indicates an avenue to NMR structures of very large biological macromolecules in solution. Proc. Natl. Acad. Sci. USA 1997, 94, 12366–12371. [Google Scholar] [CrossRef] [PubMed]
- Sattler, M.; Schleucher, J.; Griesinger, C. Heteronuclear multidimensional NMR experiments for the structure determination of proteins in solution employing pulsed field gradients. Prog. Nucl. Mag. Res. Spectrosc. 1999, 34, 93–158. [Google Scholar] [CrossRef]
- Panova, S.; Cliff, M.J.; Macek, P.; Blackledge, M.; Jensen, M.R.; Nissink, J.W.M.; Embrey, K.J.; Davies, R.; Waltho, J.P. Mapping Hidden Residual Structure within the Myc bHLH-LZ Domain Using Chemical Denaturant Titration. Structure 2019, 27, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef] [PubMed]
- Foy, N.J.; Akhrymuk, M.; Shustov, A.V.; Frolova, E.I.; Frolov, I. Hypervariable domain of nonstructural protein nsP3 of Venezuelan equine encephalitis virus determines cell-specific mode of virus replication. J. Virol. 2013, 87, 7569–7584. [Google Scholar] [CrossRef] [PubMed]
- Foy, N.J.; Akhrymuk, M.; Akhrymuk, I.; Atasheva, S.; Bopda-Waffo, A.; Frolov, I.; Frolova, E.I. Hypervariable domains of nsP3 proteins of New World and Old World alphaviruses mediate formation of distinct, virus-specific protein complexes. J. Virol. 2013, 87, 1997–2010. [Google Scholar] [CrossRef]
- Kim, D.Y.; Reynaud, J.M.; Rasalouskaya, A.; Akhrymuk, I.; Mobley, J.A.; Frolov, I.; Frolova, E.I. New World and Old World Alphaviruses Have Evolved to Exploit Different Components of Stress Granules, FXR and G3BP Proteins, for Assembly of Viral Replication Complexes. PLoS Pathog. 2016, 12, e1005810. [Google Scholar] [CrossRef]
- Meshram, C.D.; Phillips, A.T.; Lukash, T.; Shiliaev, N.; Frolova, E.I.; Frolov, I. Mutations in Hypervariable Domain of Venezuelan Equine Encephalitis Virus nsP3 Protein Differentially Affect Viral Replication. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Schulte-Herbruggen, T.; Sorensen, O.W. Clean TROSY: Compensation for relaxation-induced artifacts. J. Magn. Reson. 2000, 144, 123–128. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Nielsen, J.T.; Mulder, F.A.A. POTENCI: Prediction of temperature, neighbor and pH-corrected chemical shifts for intrinsically disordered proteins. J. Biomol. NMR 2018, 70, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Schwarzinger, S.; Kroon, G.J.; Foss, T.R.; Wright, P.E.; Dyson, H.J. Random coil chemical shifts in acidic 8 M urea: Implementation of random coil shift data in NMRView. J. Biomol. NMR 2000, 18, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Sykes, B.D.; Richards, F.M. The chemical shift index: A fast and simple method for the assignment of protein secondary structure through NMR spectroscopy. Biochemistry 1992, 31, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Markley, J.L.; Bax, A.; Arata, Y.; Hilbers, C.W.; Kaptein, R.; Sykes, B.D.; Wright, P.E.; Wuthrich, K. Recommendations for the presentation of NMR structures of proteins and nucleic acids--IUPAC-IUBMB-IUPAB Inter-Union Task Group on the standardization of data bases of protein and nucleic acid structures determined by NMR spectroscopy. Eur. J. Biochem. 1998, 256, 1–15. [Google Scholar] [CrossRef]
- Favier, A.; Brutscher, B. Recovering lost magnetization: Polarization enhancement in biomolecular NMR. J. Biomol. NMR 2011, 49, 9–15. [Google Scholar] [CrossRef]
- Lescop, E.; Schanda, P.; Brutscher, B. A set of BEST triple-resonance experiments for time-optimized protein resonance assignment. J. Magn. Reson. 2007, 187, 163–169. [Google Scholar] [CrossRef]
- Orekhov, V.; Jaravine, V.A. Analysis of non-uniformly sampled spectra with multi-dimensional decomposition. Prog. Nucl. Magn. Reson. Spectrosc. 2011, 59, 271–292. [Google Scholar] [CrossRef]
- Jaravine, V.A.; Zhuravleva, A.V.; Permi, P.; Ibraghimov, I.; Orekhov, V.Y. Hyperdimensional NMR spectroscopy with nonlinear sampling. J. Am. Chem. Soc. 2008, 130, 3927–3936. [Google Scholar] [CrossRef]
- Kazimierczuk, K.; Stanek, J.; Zawadzka-Kazimierczuk, A.; Kozminski, W. Random sampling in multidimensional NMR spectroscopy. Prog. Nucl. Magn. Reson. Spectrosc. 2010, 57, 420–434. [Google Scholar] [CrossRef]
- Agback, P.; Dominguez, F.; Pustovalova, Y.; Lukash, T.; Shiliaev, N.; Orekhov, V.Y.; Frolov, I.; Agback, T.; Frolova, E.I. Structural characterization and biological function of bivalent binding of CD2AP to intrinsically disordered domain of chikungunya virus nsP3 protein. Virology 2019, 537, 130–142. [Google Scholar] [CrossRef]
- Isaksson, L.; Mayzel, M.; Saline, M.; Pedersen, A.; Rosenlow, J.; Brutscher, B.; Karlsson, B.G.; Orekhov, V.Y. Highly Efficient NMR Assignment of Intrinsically Disordered Proteins: Application to B- and T Cell Receptor Domains. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Jaravine, V.A.; Orekhov, V.Y. Targeted acquisition for real-time NMR spectroscopy. J. Am. Chem. Soc. 2006, 128, 13421–13426. [Google Scholar] [CrossRef]
- Unnerstale, S.; Nowakowski, M.; Baraznenok, V.; Stenberg, G.; Lindberg, J.; Mayzel, M.; Orekhov, V.; Agback, T. Backbone Assignment of the MALT1 Paracaspase by Solution NMR. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Schmidt, E.; Guntert, P. A New Algorithm for Reliable and General NMR Resonance Assignment. J. Am. Chem. Soc. 2012, 134, 12817–12829. [Google Scholar] [CrossRef]
- Keller, R.L.J. The Computer Aided Resonance Assignment Tutorial; CANTINA Verlag: Goldau, Switzerland, 2004. [Google Scholar]
- Pustovalova, Y.; Mayzel, M.; Orekhov, V.Y. XLSY: Extra-Large NMR Spectroscopy. Angew. Chem. Int. Ed. 2018, 57, 14043–14045. [Google Scholar] [CrossRef]
- Tossavainen, H.; Salovaara, S.; Hellman, M.; Ihalin, R.; Permi, P. Dispersion from C-alpha or N-H: 4D experiments for backbone resonance assignment of intrinsically disordered proteins. J. Biomol. NMR 2020, 74, 147–159. [Google Scholar] [CrossRef]
- Grudziaz, K.; Zawadzka-Kazimierczuk, A.; Kozminski, W. High-dimensional NMR methods for intrinsically disordered proteins studies. Methods 2018, 148, 81–87. [Google Scholar] [CrossRef]
- Zhang, H.; Neal, S.; Wishart, D.S. RefDB: A database of uniformly referenced protein chemical shifts. J. Biomol. NMR 2003, 25, 173–195. [Google Scholar] [CrossRef]
- Schwarzinger, S.; Kroon, G.J.; Foss, T.R.; Chung, J.; Wright, P.E.; Dyson, H.J. Sequence-dependent correction of random coil NMR chemical shifts. J. Am. Chem. Soc. 2001, 123, 2970–2978. [Google Scholar] [CrossRef]
- Marsh, J.A.; Singh, V.K.; Jia, Z.C.; Forman-Kay, J.D. Sensitivity of secondary structure propensities to sequence differences between alpha- and gamma-synuclein: Implications for fibrillation. Protein Sci. 2006, 15, 2795–2804. [Google Scholar] [CrossRef]
- Mielke, S.P.; Krishnan, V.V. Characterization of protein secondary structure from NMR chemical shifts. Prog. Nucl. Magn. Reson. Spectrosc. 2009, 54, 141–165. [Google Scholar] [CrossRef]
- Meshram, C.D.; Agback, P.; Shiliaev, N.; Urakova, N.; Mobley, J.A.; Agback, T.; Frolova, E.I.; Frolov, I. Multiple Host Factors Interact with Hypervariable Domain of Chikungunya Virus nsP3 and Determine Viral Replication in Cell-Specific Mode. J. Virol. 2018. [Google Scholar] [CrossRef]






Sample Availability: Samples and additional information are available from the authors upon reasonable request. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agback, P.; Shernyukov, A.; Dominguez, F.; Agback, T.; Frolova, E.I. Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus. Molecules 2020, 25, 5824. https://doi.org/10.3390/molecules25245824
Agback P, Shernyukov A, Dominguez F, Agback T, Frolova EI. Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus. Molecules. 2020; 25(24):5824. https://doi.org/10.3390/molecules25245824
Chicago/Turabian StyleAgback, Peter, Andrey Shernyukov, Francisco Dominguez, Tatiana Agback, and Elena I. Frolova. 2020. "Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus" Molecules 25, no. 24: 5824. https://doi.org/10.3390/molecules25245824
APA StyleAgback, P., Shernyukov, A., Dominguez, F., Agback, T., & Frolova, E. I. (2020). Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus. Molecules, 25(24), 5824. https://doi.org/10.3390/molecules25245824