
Assessing the Direct Binding of Ark-Like E3 RING Ligases to Ubiquitin and Its Implication on Their Protein Interaction Network

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
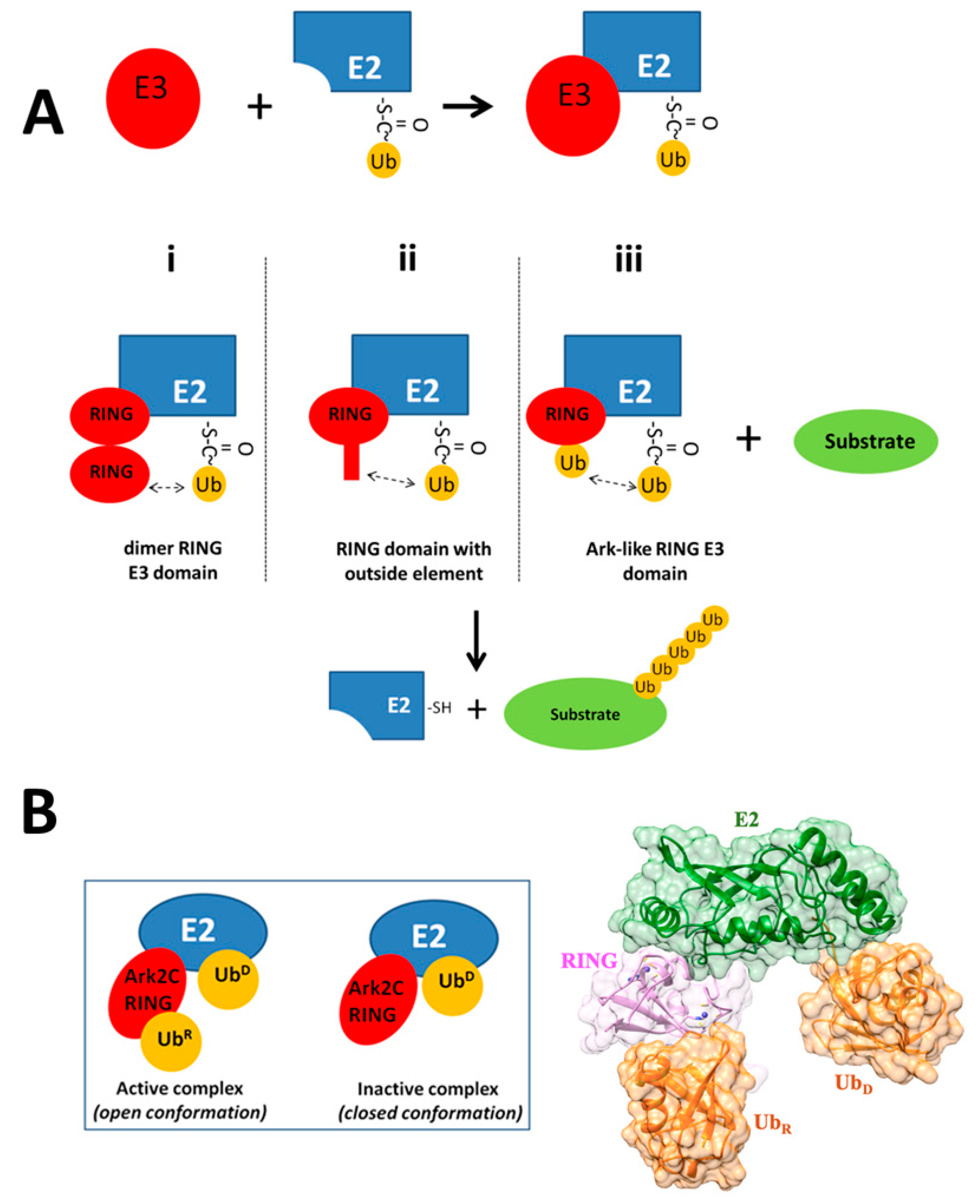
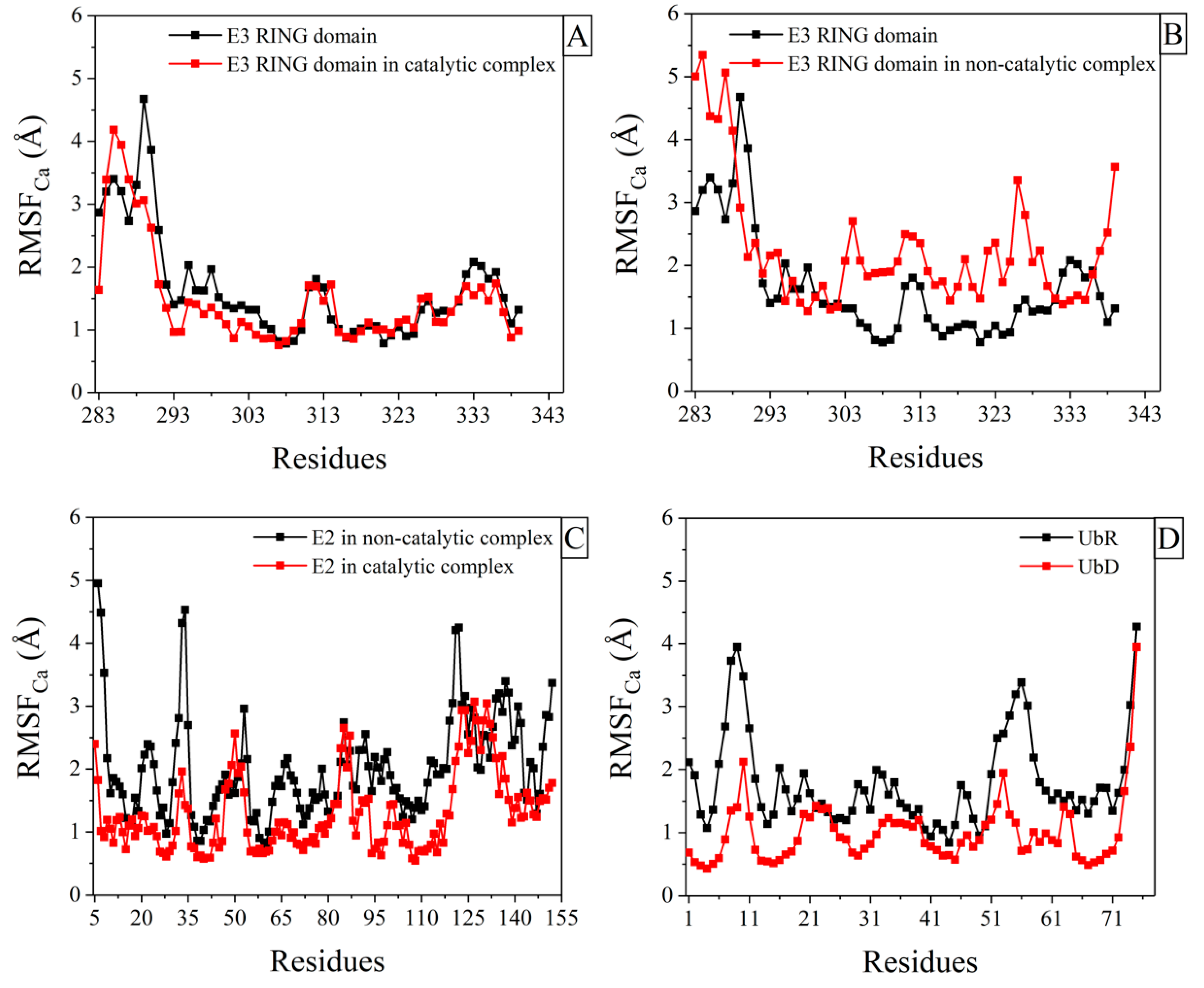
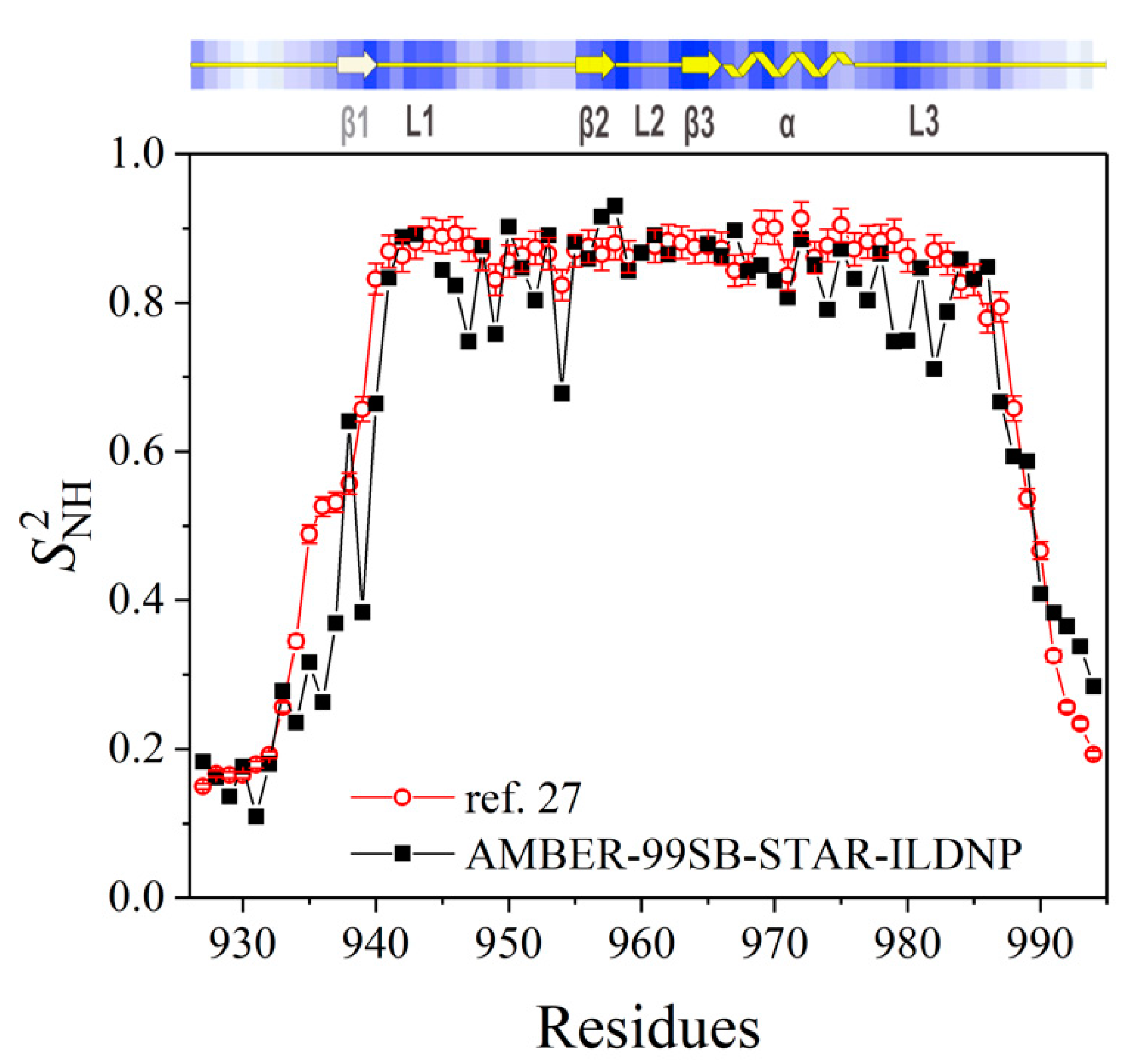
2.1. Structural Fluctuations in Ark2C RING Domain, after Binding to Ligands
2.2. Structural Fluctuations in E2 Enzyme and Ubs after Binding to Ligands
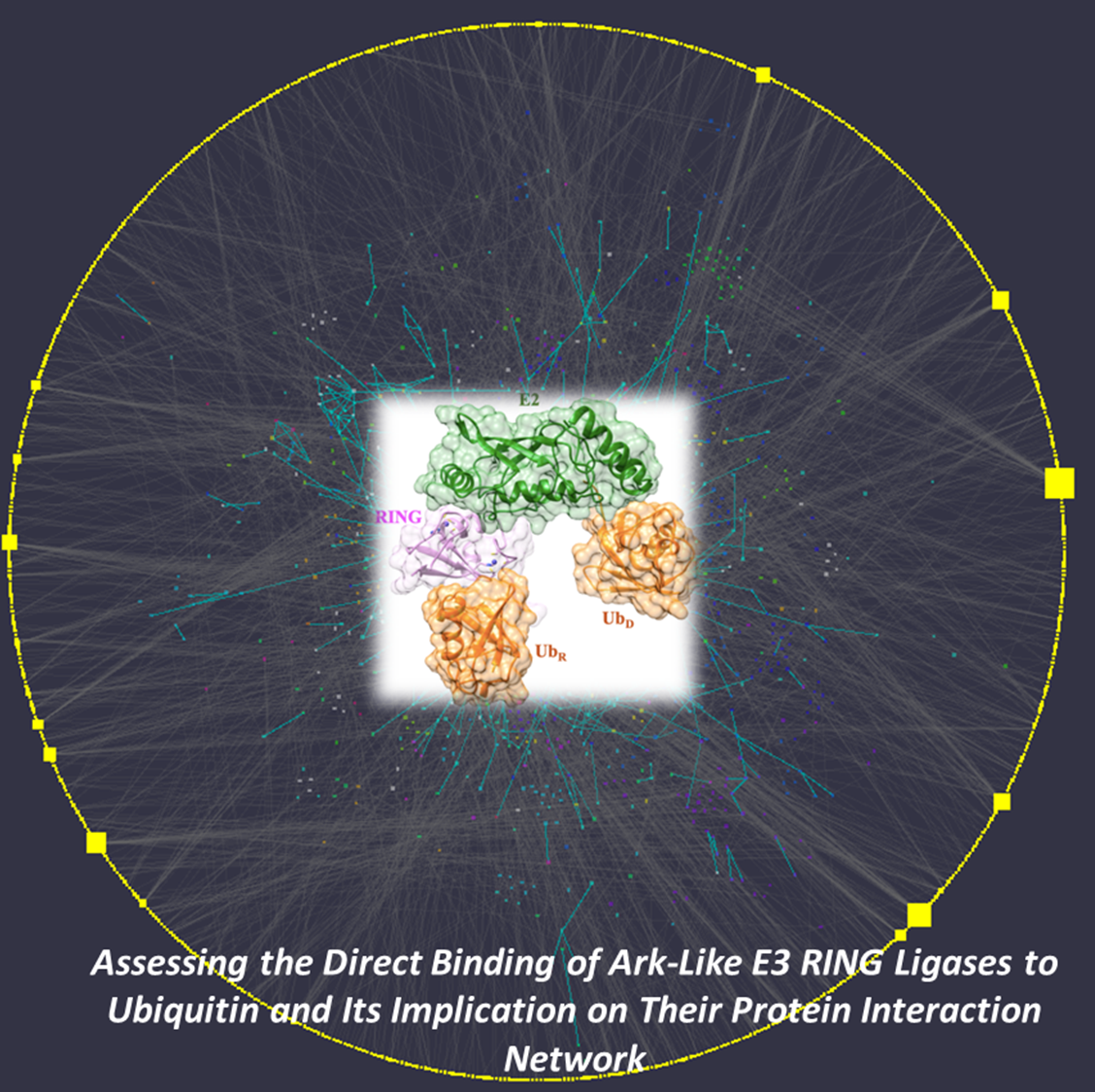
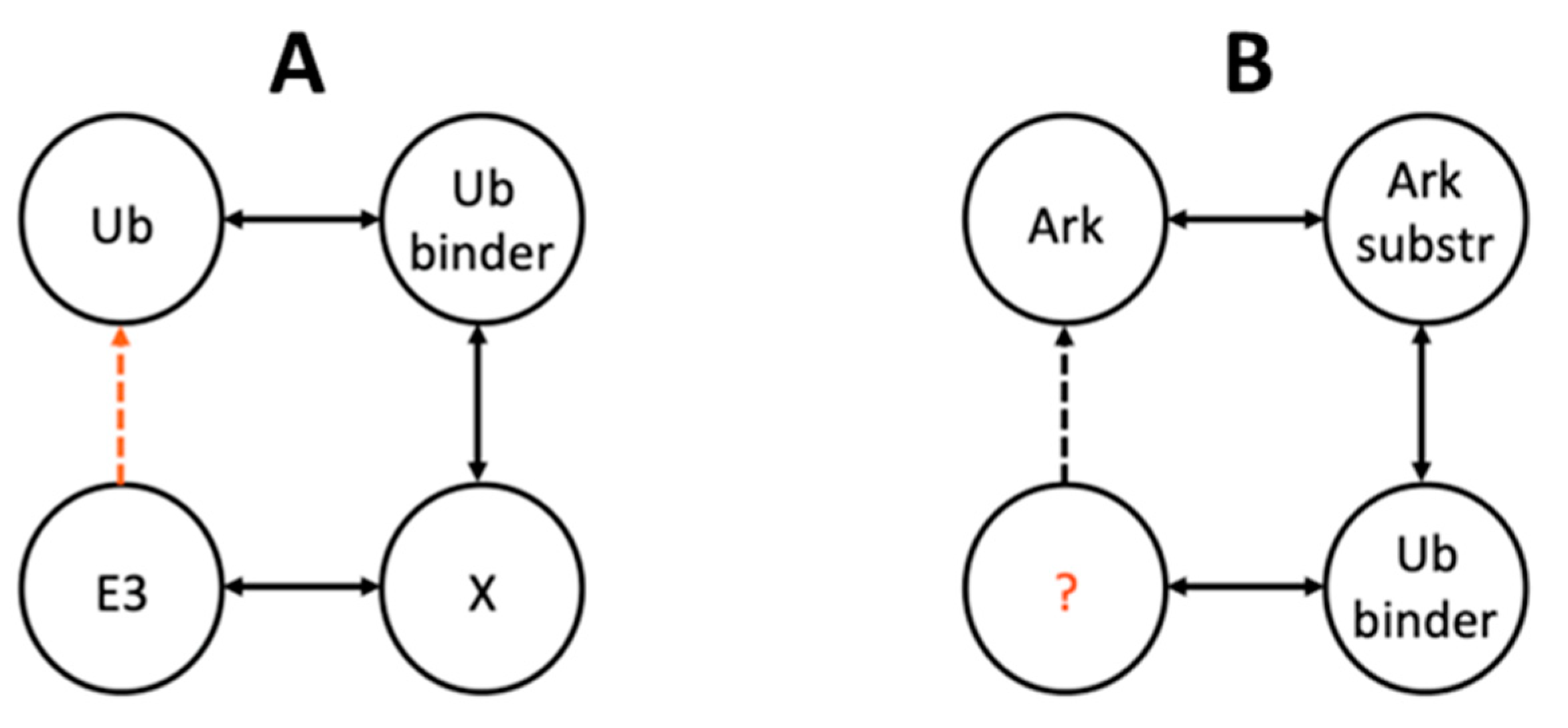
2.3. Identification of Putative Ub-Binding E3 RING Ligases
2.4. Network Analysis Reveals Potential Arkadia and Ark2C Interactors
3. Discussion
4. Materials and Methods
4.1. Molecular Dynamics
Force Field Selection
4.2. Network-Based Analysis
4.3. Functional Annotation, Subcellular Localization and Protein Structure Visualization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deshaies, R.J.; Joazeiro, C.A.P. RING Domain E3 Ubiquitin Ligases. Annu. Rev. Biochem. 2009, 99–434. [Google Scholar] [CrossRef] [PubMed]
- Schrader, E.K.; Harstad, K.G.; Matouschek, A. Targeting proteins for degradation. Nat. Chem. Biol. 2009, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y. E3 Ubiquitin Ligases as Cancer Targets and Biomarkers. Neoplasia 2006, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Bengtson, M.H.; Ulbrich, A.; Matsuda, A.; Reddy, V.A.; Orth, A.; Chanda, S.K.; Batalov, S.; Joazeiro, C.A.P. Genome-wide and functional annotation of human E3 ubiquitin ligases identifies MULAN, a mitochondrial E3 that regulates the organelle’s dynamics and signaling. PLoS ONE 2008, 3, e1487. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Spyroulias, G.A. RING finger E(3) ubiquitin ligases: Structure and drug discovery. Curr. Pharm. Des. 2009, 15, 3716–3731. [Google Scholar] [CrossRef]
- Birkou, M.; Chasapis, C.T.; Marousis, K.D.; Loutsidou, A.K.; Bentrop, D.; Lelli, M.; Herrmann, T.; Carthy, J.M.; Episkopou, V.; Spyroulias, G.A. A Residue Specific Insight into the Arkadia E3 Ubiquitin Ligase Activity and Conformational Plasticity. J. Mol. Biol. 2017, 429, 2373–2386. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Andreini, C.; Georgiopolou, A.K.; Stefanidou, M.E.; Vlamis-Gardikas, A. Identification of the zinc, copper and cadmium metalloproteome of the protozoon Tetrahymena thermophila by systematic bioinformatics. Arch. Microbiol. 2017, 199, 1141–1149. [Google Scholar] [CrossRef]
- Chasapis, C.T. Hierarchical core decomposition of RING structure as a method to capture novel functional residues within RING-type E3 ligases: A structural systems biology approach. Comput. Biol. Med. 2018, 100, 86–91. [Google Scholar] [CrossRef]
- Metzger, M.B.; Pruneda, J.N.; Klevit, R.E.; Weissman, A.M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochim. Biophys. Acta 2014, 1843, 47–60. [Google Scholar] [CrossRef]
- Plechanovová, A.; Jaffray, E.G.; Tatham, M.H.; Naismith, J.H.; Hay, R.T. Structure of a RING E3 ligase and ubiquitin-loaded E2 primed for catalysis. Nature 2012, 489, 115–120. [Google Scholar]
- Dou, H.; Buetow, L.; Hock, A.; Sibbet, G.J.; Vousden, K.H.; Huang, D.T. Structural basis for autoinhibition and phosphorylation-dependent activation of c-Cbl. Nat. Struct. Mol. Biol. 2012, 19, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, Y.; Kleffmann, T.; Linke, K.; Condon, S.M.; Hinds, M.G.; Day, C.L. Regulation of ubiquitin transfer by XIAP, a dimeric RING E3 ligase. Biochem. J. 2013, 450, 629–638. [Google Scholar] [CrossRef]
- Dou, H.; Buetow, L.; Sibbet, G.J.; Cameron, K.; Huang, D.T. Essentiality of a non-RING element in priming donor ubiquitin for catalysis by a monomeric E3. Nat. Struct. Mol. Biol. 2013, 20, 982–986. [Google Scholar] [CrossRef] [PubMed]
- Buetow, L.; Gabrielsen, M.; Anthony, N.G.; Dou, H.; Patel, A.; Aitkenhead, H.; Sibbet, G.J.; Smith, B.O.; Huang, D.T. Activation of a Primed RING E3-E2–Ubiquitin Complex by Non-Covalent Ubiquitin. Mol. Cell 2015, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.D.; Mace, P.D.; Day, C.L. Secondary ubiquitin-RING docking enhances Arkadia and Ark2C E3 ligase activity. Nat. Struct. Mol. Biol. 2016, 23, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.R.; Betarbet, R.; Gearing, M.; Gulcher, J.; Hicks, A.A.; Stefansson, K.; Lah, J.J.; Levey, A.I. PARK10 candidate RNF11 is expressed by vulnerable neurons and localizes to Lewy bodies in Parkinson disease brain. J. Neuropathol. Exp. Neurol. 2007, 66, 955–964. [Google Scholar] [CrossRef][Green Version]
- Subramaniam, V.; Li, H.; Wong, M.; Kitching, R.; Attisano, L.; Wrana, J.; Zubovits, J.; Burger, A.M.; Seth, A. The RING-H2 protein RNF11 is overexpressed in breast cancer and is a target of Smurf2 E3 ligase. Br. J. Cancer 2003, 89, 1538–1544. [Google Scholar] [CrossRef]
- Azmi, P.; Seth, A. RNF11 is a multifunctional modulator of growth factor receptor signalling and transcriptional regulation. Eur. J. Cancer. 2005, 41, 2549–2560. [Google Scholar] [CrossRef] [PubMed]
- Colland, F.; Daviet, L. Integrating a functional proteomic approach into the target discovery process. Biochimie 2004, 86, 625–632. [Google Scholar] [CrossRef]
- Li, H.; Seth, A. An RNF11: Smurf2 complex mediates ubiquitination of the AMSH protein. Oncogene 2004, 23, 1801–1808. [Google Scholar] [CrossRef]
- Shembade, N.; Parvatiyar, K.; Harhaj, N.S.; Harhaj, E.W. The ubiquitin-editing enzyme A20 requires RNF11 to downregulate NF-κB signalling. Embo J. 2009, 28, 513–522. [Google Scholar] [CrossRef]
- Nagano, Y.; Mavrakis, K.J.; Lee, K.L.; Fujii, T.; Koinuma, D.; Sase, H.; Yuki, K.; Isogaya, K.; Saitoh, M.; Imamura, T.; et al. Arkadia induces degradation of SnoN and c-Ski to enhance transforming growth factor-b signaling. J. Biol. Chem. 2007, 282, 20492–20501. [Google Scholar] [CrossRef]
- Le Scolan, E.; Zhu, Q.; Wang, L.; Bandyopadhyay, A.; Javelaud, D.; Mauviel, A.; Sun, L.; Luo, K. Transforming growth factor-b suppresses the ability of Ski to inhibit tumor metastasis by inducing its degradation. Cancer Res. 2008, 68, 3277–3328. [Google Scholar] [CrossRef] [PubMed]
- Nagano, Y.; Koinuma, D.; Miyazawa, K.; Miyazono, K. Context-dependent regulation of the expression of c-Ski protein by Arkadia in human cancer cells. J. Biochem. 2010, 147, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Briones-Orta, M.A.; Levy, L.; Madsen, C.D.; Das, D.; Erker, Y.; Sahai, E.; Hill, C.S. Arkadia Regulates Tumor Metastasis by Modulation of the T.G.F.-Pathway. Cancer Res. 2013, 1800–1810. [Google Scholar] [CrossRef]
- Mavrakis, K.J.; Andrew, R.L.; Lee, K.L.; Petropoulou, C.; Dixon, J.E.; Navaratnam, N.; Norris, D.P.; Episkopou, V. Arkadia Enhances Nodal/TGF-β Signaling by Coupling Phospho-Smad2/3 Activity and Turnover. PLoS Biol. 2007, e67. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Kandias, N.G.; Episkopou, V.; Bentrop, D.; Spyroulias, G.A. NMR-based insights into the conformational and interaction properties of Arkadia RING-H2 E3 Ub ligase. Proteins Struct. Funct. Bioinformatics 2012, 1484–1489. [Google Scholar] [CrossRef]
- Kandias, N.G.; Chasapis, C.T.; Bentrop, D.; Episkopou, V.; Spyroulias, G.A. High yield expression and NMR characterization of Arkadia E3 ubiquitin ligase RING-H2 finger domain. Biochem. Biophys. Res. Commun. 2009, 378, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Episkopou, V.; Arkell, R.; Timmons, P.M.; Walsh, J.J.; Andrew, R.L.; Swan, D. Induction of the mammalian node requires Arkadia function in the extraembryonic lineages. Nature 2001, 410, 825–830. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Loutsidou, A.K.; Orkoula, M.G.; Spyroulias, G.A. Zinc Binding Properties of Engineered RING Finger Domain of Arkadia E3 Ubiquitin Ligase. Bioinorg. Chem. Appl. 2010. [Google Scholar] [CrossRef]
- Nederveen, A.J.; Bonvin, A.M.J.J. NMR Relaxation and Internal Dynamics of Ubiquitin from a 0.2 μs MD Simulation. J. Chem. Theory Comput. 2005, 1, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Borden, K.L. RING domains: Master builders of molecular scaffolds? J. Mol. Biol. 2000, 295, 1103–1112. [Google Scholar] [CrossRef]
- Kovács, I.A.; Luck, K.; Spirohn, K.; Wang, Y.; Pollis, C.; Schlabach, S.; Bian, W.; Kim, D.K.; Kishore, N.; Hao, T.; et al. Network-based prediction of protein interactions. Nat. Commun. 2019, 10, 1240. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, L.S.; Stratakis, C.A. Structure of the Human Ubiquitin Fusion Gene Uba80 (RPS27a) and One of Its Pseudogenes. Biochem. Biophys. Res. Commun. 2000, 270, 1106–1110. [Google Scholar] [CrossRef]
- Salmerón, A.; Janzen, J.; Soneji, Y.; Bump, N.; Kamens, J.; Allen, H.; Ley, S.C. Direct phosphorylation of NF-kappaB1 p105 by the IkappaB kinase complex on serine 927 is essential for signal-induced p105 proteolysis. J. Biol. Chem. 2001, 276, 22215–22222. [Google Scholar] [CrossRef]
- Liu, L.; Wong, C.C.; Gong, B.; Yu, J. Functional significance and therapeutic implication of ring-type E3 ligases in colorectal cancer. Oncogene 2017, 37, 148–159. [Google Scholar] [CrossRef]
- Qu, F.; Xiang, Z.; Zhou, Y.; Qin, Y.; Yu, Z. Tumor necrosis factor receptor-associated factor 3 from Anodonta woodiana is an important factor in bivalve immune response to pathogen infection. Fish Shellfish Immunol. 2017, 71, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Shimbo, T.; Wade, P.A. Proteins That Read DNA Methylation. Adv. Exp. Med. Biol. 2016, 945, 303–320. [Google Scholar]
- Dove, K.K.; Klevit, R.E. RING-Between-RING E3 Ligases: Emerging Themes amid the Variations. J. Mol. Biol. 2017, 429, 3363–3375. [Google Scholar] [CrossRef]
- Chasapis, C.T. Building Bridges Between Structural and Network-Based Systems Biology. Mol. Biotechnol. 2019, 3, 221–229. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahlad, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Duan, Y.; Wu, C.; Chowdhury, S.; Lee, M.C.; Xiong, G.; Zhang, W.; Yang, R.; Cieplak, P.; Luo, R.; Lee, T.; et al. A point-charge force field for molecular mechanics simulations of proteins based on condensed-phase quantum mechanical calculations. J. Comput. Chem. 2003, 24, 1999–2012. [Google Scholar] [CrossRef] [PubMed]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins Struct. Funct. Bioinform. 2010. [Google Scholar] [CrossRef]
- Aliev, A.E.; Kulke, M.; Khaneja, H.S.; Chudasama, V.; Sheppard, T.D.; Lanigan, R.M. Motional timescale predictions by molecular dynamics simulations: Case study using proline and hydroxyproline sidechain dynamics. Proteins Struct. Funct. Bioinform. 2014, 8, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Peng, E.; Todorova, N.; Yarovsky, I. Effects of forcefield and sampling method in all-atom simulations of inherently disordered proteins: Application to conformational preferences of human amylin. PLoS ONE 2017, 12, e0186219. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D.; Rao, J.S.; Segelken, E.; Cruz, L. Force-Field Induced Bias in the Structure of Aβ21–30, A Comparison of OPLS, AMBER, CHARMM, and GROMOS Force Fields. J. Chem. Inf. Model. 2015, 55, 2587–2595. [Google Scholar] [CrossRef]
- Robustelli, P.; Piana, S.; Shaw, D.E. Developing a molecular dynamics force field for both folded and disordered protein states. Proc. Natl. Acad. Sci. USA 2018, 115, E4758. [Google Scholar] [CrossRef] [PubMed]
- Lindorff-Larsen, K.; Maragakis, P.; Piana, S.; Eastwood, M.P.; Dror, R.O.; Shaw, D.E. Systematic Validation of Protein Force Fields against Experimental Data. PLoS ONE 2012, 7, e32131. [Google Scholar] [CrossRef]
- Medvar, B.; Raghuram, V.; Pisitkun, T.; Sarkar, A.; Knepper, M.A. Comprehensive database of human E3 ubiquitin ligases: Application to aquaporin-2 regulation. Physiol. Genom. 2016, 48, 502. [Google Scholar] [CrossRef]
- Li, Y.; Xie, P.; Lu, L.; Wang, J.; Diao, L.; Liu, Z.; Guo, F.; He, Y.; Liu, Y.; Huang, Q.; et al. An integrated bioinformatics platform for investigating the human E3 ubiquitin ligase-substrate interaction network. Nat. Commun. 2017, 8, 347. [Google Scholar] [CrossRef] [PubMed]
- Iconomou, M.; Saunders, D.N. Systematic approaches to identify E3 ligase substrates. Biochem. J. 2016, 473, 4083–4101. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, Y.; Lin, S.; Guo, Y.; Deng, W.; Zhang, Y.; Guo, A.; Xue, Y. iUUCD 2.0: An update with rich annotations for ubiquitin and ubiquitin-like conjugations. Nucleic Acids Res. 2018, 46, D447–D453. [Google Scholar] [CrossRef] [PubMed]
- Oughtred, R.; Stark, C.; Breitkreutz, B.J.; Rust, J.; Boucher, L.; Chang, C.; Kolas, N.; O’Donnell, L.; Leung, G.; McAdam, R.; et al. The BioGRID interaction database: 2019 update. Nucleic Acids Res. 2019, 47, D529–D541. [Google Scholar] [CrossRef] [PubMed]
- Peana, M.; Chasapis, C.T.; Simula, G.; Medici, S.; Zoroddu, M.A. A Model for Manganese interaction with Deinococcus radiodurans proteome network involved in ROS response and defense. J. Trace Elem. Med. Biol. 2018, 50, 465–473. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Goddard, T.; Huang, C.; Ferrin, T. Software Extensions to UCSF Chimera for Interactive Visualization of Large Molecular Assemblies. Structure 2005, 13, 473–482. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E3 RING Ligase (UniProt ID) | Protein Name | Connectivity Degree with Ub-Binding Proteins |
|---|---|---|
| P22681 | E3 ubiquitin-protein ligase C.B.L. | 35 |
| Q00987 | E3 ubiquitin-protein ligase Mdm2 | 31 |
| O60260 | E3 ubiquitin-protein ligase parkin | 24 |
| Q8IUQ4 | E3 ubiquitin-protein ligase SIAH1 | 18 |
| Q86TM6 | E3 ubiquitin-protein ligase synoviolin | 15 |
| Q9Y4K3 | TNF receptor-associated factor 6 | 14 |
| O43255 | E3 ubiquitin-protein ligase SIAH2 | 13 |
| P38398 | Breast cancer type 1 susceptibility protein | 13 |
| P35226 | Polycomb complex protein BMI-1 | 12 |
| Q13490 | Baculoviral IAP repeat-containing protein 2 | 11 |
| Ub-Binding Protein (UniProt ID) | Connectivity Degree with Arkadia | Potential Arkadia Interactors | |
|---|---|---|---|
| Ark2C | Q96A37 | 7 | O60260, O60674, P00441, P00533, P0CG47, P0CG48, P51965, P60709, P62837, Q07666, Q13501, Q13618, Q92993 |
| Q8IYW5 | 6 | ||
| Q9UBN7 | 5 | ||
| Q8TF42 | 5 | ||
| P42566 | 5 | ||
| Arkadia | P03372 | 2 | P0CG48, P0CG47, P62979, P62987, O14920 |
| No | Protein Complex Description | PDB ID | Force Field | Size of Cubic Box, x (Å) | Simulation Time (ns) | |
|---|---|---|---|---|---|---|
| 1 | Arkadia E3 Ligase (RNF111) | SOLUTION NMR | 2KIZ | AMBER-03 | 60 | 150 |
| 2 | AMBER-99SB-ILDN-2010 | 80 | 150 | |||
| 3 | AMBER-99SB-STAR-ILDN-2014 | 80 | 150 | |||
| 4 | Ark2C E3 Ligase (RNF165) | X-RAY DIFFRACTION 2.65 Å | 5D0I | AMBER-99SB-STAR-ILDN-2014 | 80 | 150 |
| 5 | E3-E2-Ub non-catalytic complex | X-RAY DIFFRACTION 1.91 Å | 5D0M | AMBER-99SB-STAR-ILDN-2014 | 92 | 150 |
| 6 | Ub-E3-E2-Ub catalytic complex | X-RAY DIFFRACTION 2.65 Å | 5D0K | AMBER-99SB-STAR-ILDN-2014 | 100 | 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mintis, D.G.; Chasapi, A.; Poulas, K.; Lagoumintzis, G.; Chasapis, C.T. Assessing the Direct Binding of Ark-Like E3 RING Ligases to Ubiquitin and Its Implication on Their Protein Interaction Network. Molecules 2020, 25, 4787. https://doi.org/10.3390/molecules25204787
Mintis DG, Chasapi A, Poulas K, Lagoumintzis G, Chasapis CT. Assessing the Direct Binding of Ark-Like E3 RING Ligases to Ubiquitin and Its Implication on Their Protein Interaction Network. Molecules. 2020; 25(20):4787. https://doi.org/10.3390/molecules25204787
Chicago/Turabian StyleMintis, Dimitris G., Anastasia Chasapi, Konstantinos Poulas, George Lagoumintzis, and Christos T. Chasapis. 2020. "Assessing the Direct Binding of Ark-Like E3 RING Ligases to Ubiquitin and Its Implication on Their Protein Interaction Network" Molecules 25, no. 20: 4787. https://doi.org/10.3390/molecules25204787
APA StyleMintis, D. G., Chasapi, A., Poulas, K., Lagoumintzis, G., & Chasapis, C. T. (2020). Assessing the Direct Binding of Ark-Like E3 RING Ligases to Ubiquitin and Its Implication on Their Protein Interaction Network. Molecules, 25(20), 4787. https://doi.org/10.3390/molecules25204787