Control of Growth and Persistence of Listeria monocytogenes and β-Lactam-Resistant Escherichia coli by Thymol in Food Processing Settings

 ,
,  ,
, 
Abstract
1. Introduction
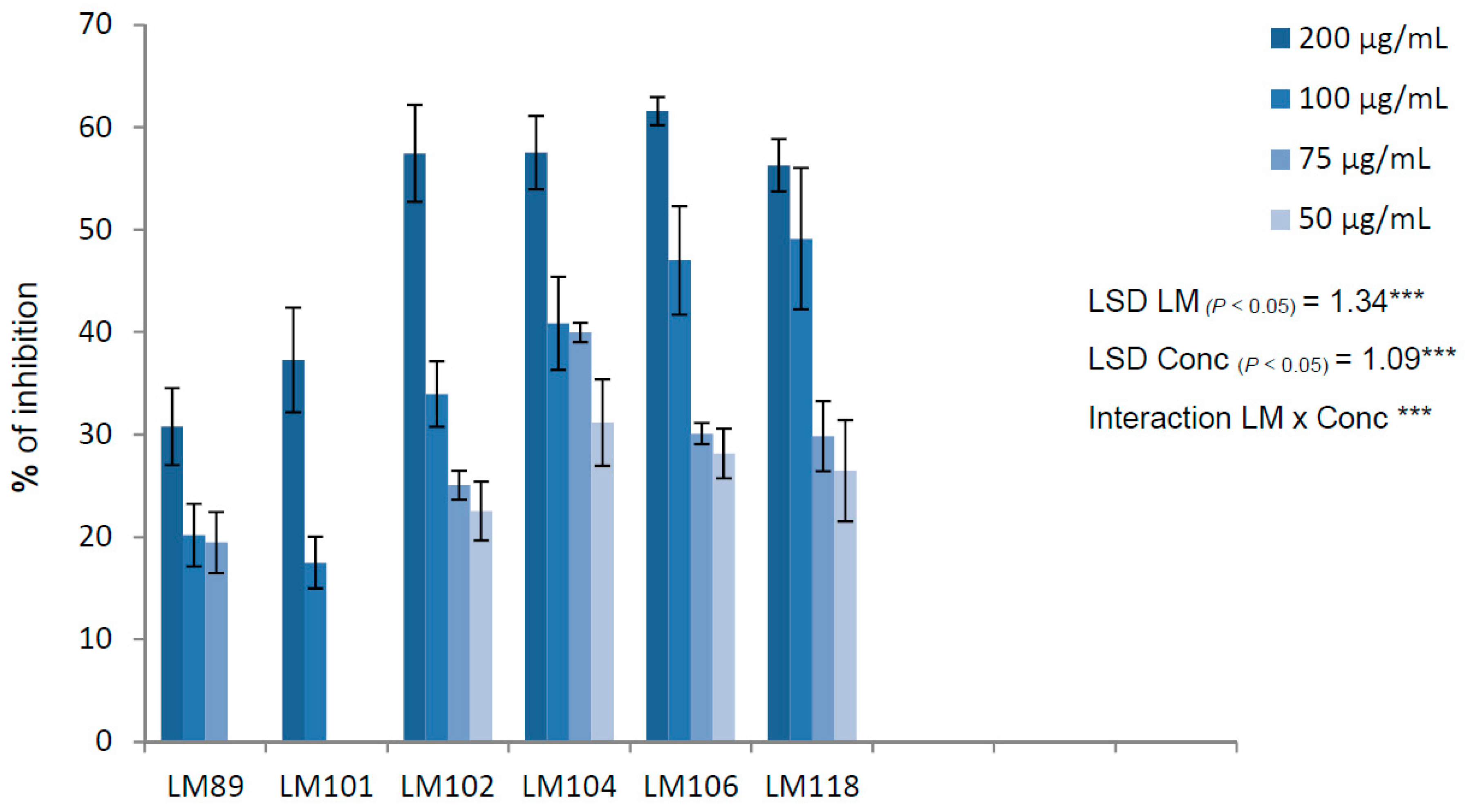
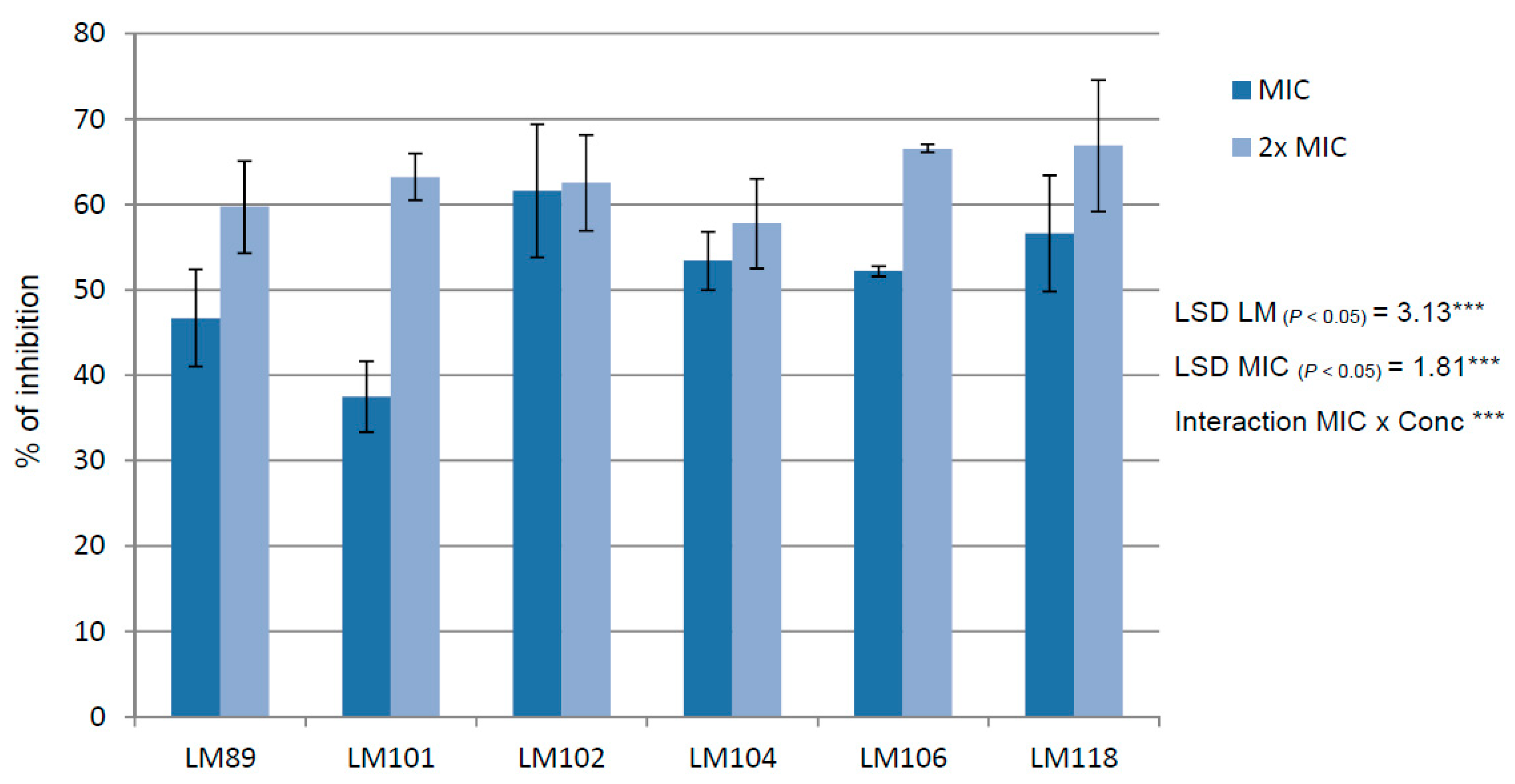
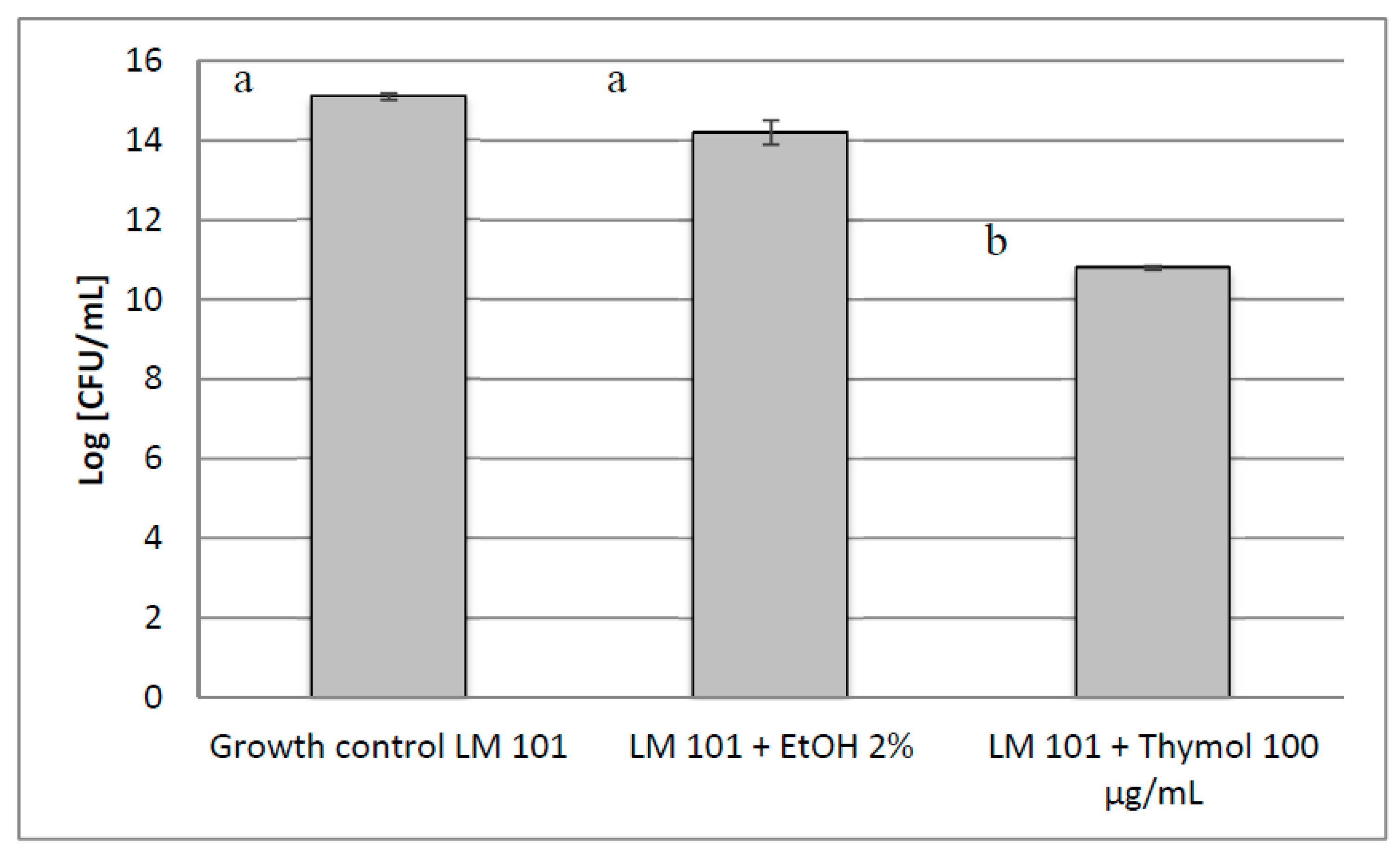
2. Results and Discussion
3. Materials and Methods
3.1. Bacterial Strains
3.2. Genetic Analysis on E. coli Isolates
3.3. Determination of Minimal Inhibitory Concentrations (MICs)
3.4. Evaluation of Biofilm Formation and Inhibition
3.5. Inhibition of Biofilm Formation
3.6. Anti-Biofilm Activity
3.7. Inhibition of Biofilm Formation on Circular Stainless-Steel Coupons Using the Viable Plate Count Method
3.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jasovský, D.; Littmann, J.; Zorzet, A.; Cars, O. Antimicrobial resistance—A threat to the world’s sustainable development. Ups. J. Med. Sci. 2016, 121, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial resistance in humans, livestock and the wider environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. 2017, 70, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.-N.; Tang, G.-Y.; Li, H.-B. Antibacterial and Antifungal Activities of Spices. Int. J. Mol. Sci. 2017, 18, 1283. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Schillaci, D.; Spanò, V.; Parrino, B.; Carbone, A.; Montalbano, A.; Barraja, P.; Diana, P.; Cirrincione, G.; Cascioferro, S. Pharmaceutical Approaches to Target Antibiotic Resistance Mechanisms. J. Med. Chem. 2017, 60, 8268–8297. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef]
- Kurpas, M.; Wieczorek, K.; Osek, J. Ready-to-eat Meat Products as a Source of Listeria monocytogenes. J. Vet. Res. 2018, 62, 49–55. [Google Scholar] [CrossRef]
- Sandasi, M.; Leonard, C.; Viljoen, A. The effect of five common essential oil components on Listeria monocytogenes biofilms. Food Control 2008, 19, 1070–1075. [Google Scholar] [CrossRef]
- Geraci, A.; Di Stefano, V.; Di Martino, E.; Schillaci, D.; Schicchi, R. Essential oil components of orange peels and antimicrobial activity. Nat. Prod. Res. 2017, 31, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Nyila, M.A.; Leonard, C.M.; Hussein, A.A.; Lall, N. Activity of South African medicinal plants against Listeria monocytogenes biofilms, and isolation of active compounds from Acacia karroo. S. Afr. J. Bot. 2012, 78, 220–227. [Google Scholar] [CrossRef]
- Ferreira, S.; Domingues, F. The antimicrobial action of resveratrol against Listeria monocytogenes in food-based models and its antibiofilm properties. J. Sci. Food Agric. 2016, 96, 4531–4535. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Sudano Roccaro, A.; Bisignano, G.; Marino, A.; Cannatelli, M.A.; Pizzimenti, F.C.; Cioni, P.L.; Procopio, F.; Blanco, A.R. Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 2007, 56, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Braga, P.C.; Culici, M.; Alfieri, M.; Dal Sasso, M. Thymol inhibits Candida albicans biofilm formation and mature biofilm. Int. J. Antimicrob. Agents 2008, 31, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Yair, Y.; Gophna, U. Pandemic Bacteremic Escherichia coli Strains: Evolution and Emergence of Drug-Resistant Pathogens. Curr. Top. Microbiol. Immunol. 2018, 416, 163–180. [Google Scholar]
- Kerekes, E.-B.; Vidács, A.; Török, J.J.; Gömöri, C.; Petkovits, T.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Krisch, J. Anti-listerial effect of selected essential oils and thymol. Acta Biol. Hung. 2016, 67, 333–343. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, H.; Zhang, W.; Mu, H.; Sun, C.; Duan, J. Synergy among thymol, eugenol, berberine, cinnamaldehyde and streptomycin against planktonic and biofilm-associated food-borne pathogens. Lett. Appl. Microbiol. 2015, 60, 421–430. [Google Scholar] [CrossRef]
- Orhan-Yanıkan, E.; da Silva-Janeiro, S.; Ruiz-Rico, M.; Jiménez-Belenguer, A.I.; Ayhan, K.; Barat, J.M. Essential oils compounds as antimicrobial and antibiofilm agents against strains present in the meat industry. Food Control 2019, 101, 29–38. [Google Scholar] [CrossRef]
- Hebbelstrup Jensen, B.; Poulsen, A.; Hebbelstrup Rye Rasmussen, S.; Struve, C.; Engberg, J.H.; Friis-Møller, A.; Boisen, N.; Jønsson, R.; Petersen, R.F.; Petersen, A.M.; et al. Genetic Virulence Profile of Enteroaggregative Escherichia coli Strains Isolated from Danish Children with Either Acute or Persistent Diarrhea. Front. Cell. Infect. Microbiol. 2017, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Saralaya, V.; Adhikari, P.; Shenoy, S.; Baliga, S.; Hegde, A. Characterization of Escherichia coli Phylogenetic Groups Associated with Extraintestinal Infections in South Indian Population. Ann. Med. Health Sci. Res. 2015, 5, 241–246. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Mahizan, N.A.; Yang, S.-K.; Moo, C.-L.; Song, A.A.-L.; Chong, C.-M.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.E.; Lai, K.-S. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR) Pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jeon, S.; Rhie, H.; Lee, B.; Park, M.; Lee, H.; Lee, J.; Kim, S. Rapid Detection of Extended Spectrum β-Lactamase (ESBL) for Enterobacteriaceae by use of a Multiplex PCR-based Method. Infect. Chemother. 2009, 41, 181–184. [Google Scholar] [CrossRef]
- Bai, J.; Paddock, Z.D.; Shi, X.; Li, S.; An, B.; Nagaraja, T.G. Applicability of a multiplex PCR to detect the seven major Shiga toxin-producing Escherichia coli based on genes that code for serogroup-specific O-antigens and major virulence factors in cattle feces. Foodborne Pathog. Dis. 2012, 9, 541–548. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Schillaci, D.; Spinello, A.; Cusimano, M.G.; Cascioferro, S.; Barone, G.; Vitale, M.; Arizza, V. A peptide from human β thymosin as a platform for the development of new anti-biofilm agents for Staphylococcus spp. and Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2016, 32, 124. [Google Scholar] [CrossRef]
- Schillaci, D.; Petruso, S.; Raimondi, M.V.; Cusimano, M.G.; Cascioferro, S.; Scalisi, M.; La Giglia, M.A.; Vitale, M. Pyrrolomycins as potential anti-staphylococcal biofilms agents. Biofouling 2010, 26, 433–438. [Google Scholar] [CrossRef]
Sample Availability: Sample of the compound thymol is available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| aatA | shf | Irp2 | wzxO103 | ehxA | Thymol | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli Isolate | Phylogenetic Group | TEM | CTX MIV | OXA | SHV | CTX MI | CMYII | CTX MII | DHA | Dispersin Transporter | Cryptic Open Reading | Yersinia Bactin | O-polysaccharide Export | Virulence Gene | Biofilm Formation | MICs (µg/mL) |
| 334 | A | + | − | − | − | − | − | − | − | − | − | − | + | + | − | 400 |
| 335 | B1 | + | − | − | − | − | − | − | − | + | − | − | + | + | − | 300 |
| 336 | B1 | + | − | − | − | − | − | − | − | − | − | − | + | + | + | 400 |
| 337 | C | + | − | − | − | − | − | − | − | + | − | − | + | + | − | 400 |
| 339 | C | + | − | − | − | − | − | − | − | + | − | − | + | + | − | 400 |
| Percentage Inhibition Biofilm Formation | ||||
|---|---|---|---|---|
| Thymol | 200 μg/mL | 100 μg/mL | 75 μg/mL | 50 μg/mL |
| Inhibition % | 94% ± 4.0% | 59.4% ± 1.6% | 55% ± 1.0% | 54.5% ± 3.5% |
| L. monocytogenes | Origin Product | Thymol MIC (µg/mL) | OD |
|---|---|---|---|
| 45 | Meat | 250 | 0.424 ± 0.008 efg |
| 46 | Meat | 250 | 0.447 ± 0.04 ef |
| 47 | Vegetable | 250 | 0.228 ± 0.03 m |
| 48 | Vegetable | 400 | 0.148 ± 0.02 n |
| 49 | Vegetable | 400 | 0.593 ± 0.08 c |
| 50 | Vegetable | 400 | 0.376 ± 0.05 ghij |
| 51 | Vegetable | 400 | 0.520 ± 0.06 d |
| 52 | Vegetable | 400 | 0.290 ± 0.04 l |
| 53 | Dairy | 400 | 0.388 ± 0.03 ghi |
| 79 | Dairy | 400 | 0.388 ± 0.04 ghi |
| 85 | Meat | 400 | 0.386 ± 0.01 ghi |
| 89 | Meat | 400 | 0.663 ± 0.07 b |
| 96 | Dairy | 400 | 0.225 ± 0.03 m |
| 98 | Dairy | 400 | 0.348 ± 0.003 ijk |
| 99 | Fish | 400 | 0.461 ± 0.04 e |
| 100 | Fish | 400 | 0.323 ± 0.006 k |
| 101 | Fish | 400 | 0.888 ± 0.12 a |
| 102 | Fish | 400 | 0.536 ± 0.03 d |
| 103 | Fish | 400 | 0.598±0.03 c |
| 104 | Fish | 400 | 0.553 ± 0.05 d |
| 105 | Dairy | 400 | 0.370 ± 0.05 hij |
| 106 | Dairy | 250 | 0.537 ± 0.06 d |
| 107 | Fish | 250 | 0.414 ± 0.05 fgh |
| 112 | Meat | 250 | 0.335 ± 0.03 jk |
| 118 | Dairy | 250 | 0.542 ± 0.02 d |
| ATCC 7644 | 800 | 0.424 ± 0.06 efg | |
| ATCC 19114 | 800 | 0.543 ± 0.07 d | |
| ATCC 19115 | 800 | 0.252 ± 0.02 lm | |
| NCTC 10887 | 800 | 0.405 ± 0.04 fgh | |
| NCTC 18890 | 800 | 0.251 ± 0.02 lm | |
| LSD (p < 0.05) | 0.033 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cusimano, M.G.; Di Stefano, V.; La Giglia, M.; Di Marco Lo Presti, V.; Schillaci, D.; Pomilio, F.; Vitale, M. Control of Growth and Persistence of Listeria monocytogenes and β-Lactam-Resistant Escherichia coli by Thymol in Food Processing Settings. Molecules 2020, 25, 383. https://doi.org/10.3390/molecules25020383
Cusimano MG, Di Stefano V, La Giglia M, Di Marco Lo Presti V, Schillaci D, Pomilio F, Vitale M. Control of Growth and Persistence of Listeria monocytogenes and β-Lactam-Resistant Escherichia coli by Thymol in Food Processing Settings. Molecules. 2020; 25(2):383. https://doi.org/10.3390/molecules25020383
Chicago/Turabian StyleCusimano, Maria Grazia, Vita Di Stefano, Maria La Giglia, Vincenzo Di Marco Lo Presti, Domenico Schillaci, Francesco Pomilio, and Maria Vitale. 2020. "Control of Growth and Persistence of Listeria monocytogenes and β-Lactam-Resistant Escherichia coli by Thymol in Food Processing Settings" Molecules 25, no. 2: 383. https://doi.org/10.3390/molecules25020383
APA StyleCusimano, M. G., Di Stefano, V., La Giglia, M., Di Marco Lo Presti, V., Schillaci, D., Pomilio, F., & Vitale, M. (2020). Control of Growth and Persistence of Listeria monocytogenes and β-Lactam-Resistant Escherichia coli by Thymol in Food Processing Settings. Molecules, 25(2), 383. https://doi.org/10.3390/molecules25020383