
Cytotoxicity of Oleandrin Is Mediated by Calcium Influx and by Increased Manganese Uptake in Saccharomyces cerevisiae Cells


Department of Organic Chemistry, Biochemistry and Catalysis, Faculty of Chemistry, University of Bucharest, Sos. Panduri 90-92, 050663 Bucharest, Romania
*
Author to whom correspondence should be addressed.
Molecules 2020, 25(18), 4259; https://doi.org/10.3390/molecules25184259
Submission received: 17 August 2020
/
Revised: 9 September 2020
/
Accepted: 15 September 2020
/
Published: 17 September 2020
(This article belongs to the Special Issue Cytotoxic Activity of Plant Extracts)
Abstract
:Oleandrin, the main component of Nerium oleander L. extracts, is a cardiotoxic glycoside with multiple pharmacological implications, having potential anti-tumoral and antiviral characteristics. Although it is accepted that the main mechanism of oleandrin action is the inhibition of Na+/K+-ATPases and subsequent increase in cell calcium, many aspects which determine oleandrin cytotoxicity remain elusive. In this study, we used the model Saccharomyces cerevisiae to unravel new elements accounting for oleandrin toxicity. Using cells expressing the Ca2+-sensitive photoprotein aequorin, we found that oleandrin exposure resulted in Ca2+ influx into the cytosol and that failing to pump Ca2+ from the cytosol to the vacuole increased oleandrin toxicity. We also found that oleandrin exposure induced Mn2+ accumulation by yeast cells via the plasma membrane Smf1 and that mutants with defects in Mn2+ homeostasis are oleandrin-hypersensitive. Our data suggest that combining oleandrin with agents which alter Ca2+ or Mn2+ uptake may be a way of controlling oleandrin toxicity.
1. Introduction
Nerium oleander L., commonly known as oleander, is an ornamental shrub with both pharmacological and toxicological properties, whose parts have been used in ethnomedicine since ancient times as natural remedies against cardiac illnesses, cancer, diabetes, asthma, skin diseases, inflammation, etc. [1]. Oleander extracts need to be regarded with caution as they are poisonous in high doses, having important cardiotoxic effects [2,3,4]. Nevertheless, many of the individual components of oleander extracts have been found to have anti-tumor, anti-proliferative, anti-inflammatory and even antiviral properties [5,6,7,8]. One of the emblematic components of oleander extracts is oleandrin (PubChem CID 11541511), a cardiotonic glycoside similar in toxicity and structure to digitoxin from Digitalis purpurea L. [7]. Oleandrin (Figure 1a) is a lipid-soluble glycoside comprised of oleandrigenin (the steroid aglycone) and D-diginosyl (a sugar-like moiety) which slightly increases oleandrin’s water solubility, which otherwise is very low [9]. Oleandrin is mainly responsible for the toxicity of oleander sap and, just as with digitoxin, it is thought to act as a cardiotonic by inhibiting the sodium and potassium ATPases (Na+/K+-ATPase) and subsequently increasing Ca2+ concentration, resulting in activation of various cell survival and death pathways [10,11]. In spite of its toxicity, oleandrin has been increasingly investigated as several studies indicated its potential as an anticancer [12,13,14,15,16] as well as antiviral drug [8,17,18,19]. The therapeutic potential of oleandrin is hampered by its cytotoxicity and by the fact that although several signaling cascades targeted by oleandrin through inhibition of Na+/K+-ATPase have been identified [20], and that it is considered that oleandrin may cause destruction of tumor cells by inducing oxidative stress through generation or reactive oxygen species (ROS) [21], many aspects which mediate oleandrin toxicity still remain obscure.
In this study, we made use of the model microorganism Saccharomyces cerevisiae to investigate oleandrin toxicity on yeast cells. S. cerevisiae is a simplified model of the eukaryotic cell used to elucidate many of the molecular mechanisms conserved in higher eukaryotes due to the ease of manipulation, tractable genetics, exhaustive genome annotation and less restrictive ethical constraints [22,23,24]. So far, no study concerning the effect of oleander extracts or oleandrin has been reported in S. cerevisiae. As oleandrin was shown to alter the fluidity of the human cell membrane [25], we hypothesized that the primary interaction between oleandrin and the yeast cells would occur at the plasma membrane level. We therefore tested the oleandrin toxicity on S. cerevisiae mutants with defects in the cell membrane transport of monovalent ions (Na+ and K+), Ca2+ or essential metal ions.
In S. cerevisiae, the movement of Na+ and K+ across the plasma membrane is ensured by Ena1 P-type Na+/H+ ATPase, Nha1 Na+/H+ antiporter, outward-rectifier K+ channel Tok1 and the Trk1p–Trk2p potassium transport system [26]. Regulation of these transporters has been extensively reviewed [26] and it was shown that salt stress and alkaline stress induce calcium-mediated responses by generating Ca2+ flux into the cytosol [27,28]. Abrupt increases in the cytosolic Ca2+ ([Ca2+]cyt) represent a universal mechanism to trigger signaling cascades involved in cell adaptation, survival or death [29]. In S. cerevisiae, the increase in [Ca2+]cyt occurs through Ca2+ entry into the cytosol via the Cch1/Mid1 channel situated at the plasma membrane [27,30] or via the vacuolar transient receptor potential channel TRPY1 (formerly known as Yvc1) [31,32,33]. Cch1 is similar to the pore-forming subunit (α1) of the plasma membrane, and voltage-gated Ca2+ channels (VGCC) from higher eukaryotes, including humans [34]. Cch1 interacts and partially co-localizes with Mid1p, a stretch-activated cation channel which resembles the VGCC α2/δ regulatory subunits and Na+ leak channel non-selective (NALCN)-associated proteins [35]. Prolonged high [Ca2+]cyt is detrimental to cells, therefore the normal very low [Ca2+]cyt must be restored through the action of Ca2+ pumps and exchangers [33]. In S. cerevisiae, this is done by the concerted actions of the vacuolar Ca2+-ATPase Pmc1 (similar to mammalian PMCA1a) [36] and of the vacuolar Ca2+/H+ exchanger Vcx1 [37,38] (which independently transport [Ca2+]cyt into the vacuole) and by the secretory Ca2+-ATPase Pmr1, which pumps [Ca2+]cyt into the endoplasmic reticulum (ER) and Golgi [39,40]. Pmr1p is the prototype of a family of transporters known as SPCA (Secretory Pathway Ca2+-ATPases) with members found also in mammals [41]. Defects in the human ortholog of PMR1, ATP2C1, are associated with Hailey-Hailey disease [42].
As oleandrin was shown to sensitize human osteosarcoma cells to cisplatin by preventing degradation of the copper transporter CTR1 [15], we also studied the cytotoxicity of oleandrin against S. cerevisiae mutants with defects in the transport of essential metal ions across the plasma membrane. In S. cerevisiae, there is an intricate system of transporters involved in the high- or low-affinity transport of essential metals, with both high and low specificity [43], e.g., Ctr1 (Cu+ transporter [44]), Fet3/Ftr1 (complex involved in the transport of Fe3+ and Cu2+, [45]), Fet4 (low-affinity transporter for Fe2+/3+ and other transition metal ions [46]), Pho84 (phosphate transporter and a low-affinity divalent metal transporter [47]), Smf1 (divalent metal ion transporter with broad specificity and with high affinity for Mn2+ [48,49]), Zrt1 (high-affinity Zn2+ transporter [50]) and Zrt2 (low-affinity Zn2+ transprter [51]). The data obtained in this study indicated that oleandrin exposure induced transient elevations in [Ca2+]cyt, but also an unexpected increase in Smf1-dependent Mn2+ accumulation which was accountable for the increased sensitivity to oleandrin of mutants with defects in Mn2+ homeostasis.
2. Results
2.1. Toxicity of Oleandrin towards Saccharomyces cerevsiae Cells
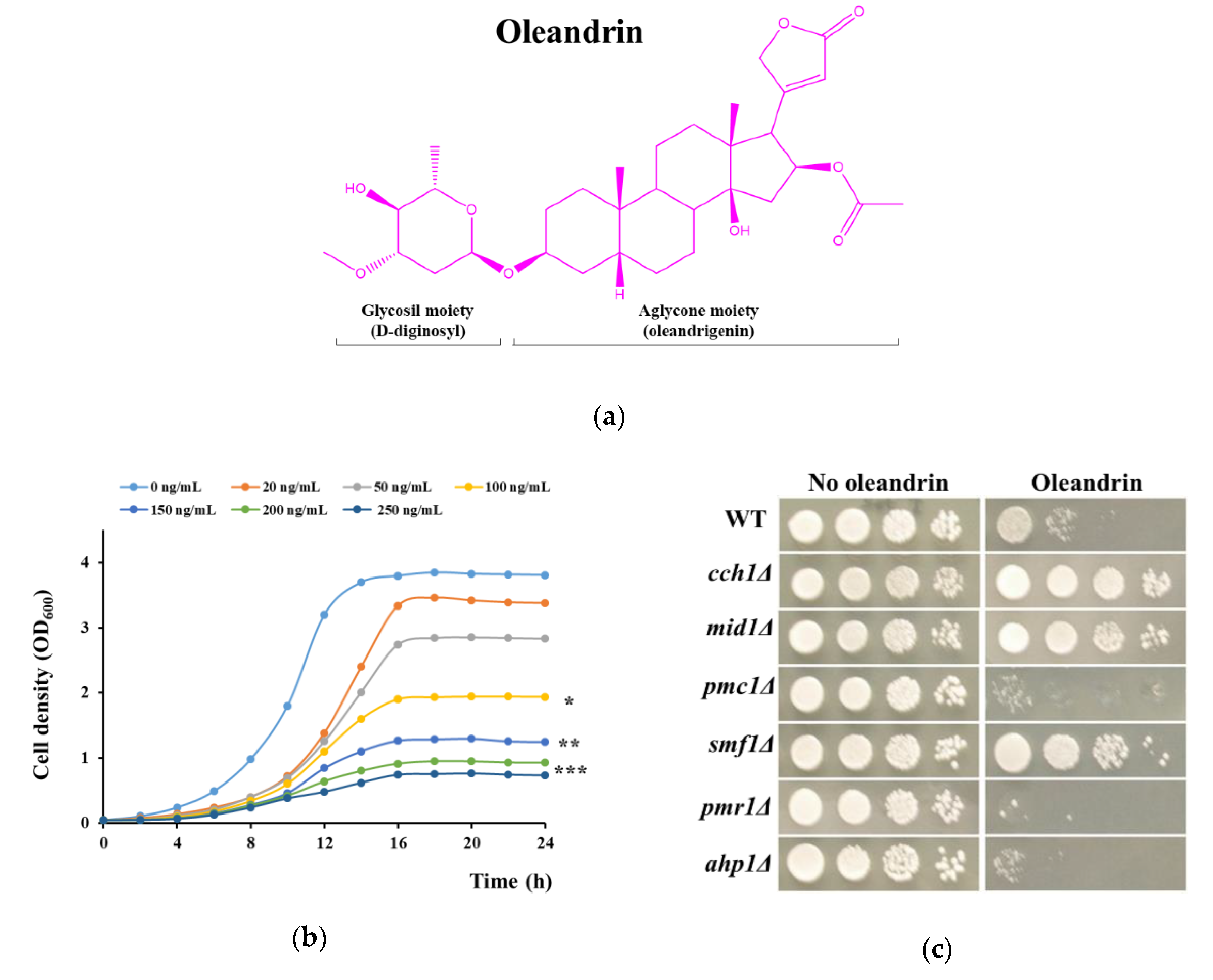
The investigation of the potential action of oleandrin upon S. cerevisiae cells is problematic due to the low solubility of oleandrin in aqueous environments, which does not surpass 1.5 mg/L [9]. We firstly prepared a saturated aqueous solution of oleandrin which was added to yeast culture media at various sub-saturation ratios. It was noted that media containing 500 ng/mL (approximately 1/3 saturation) completely killed the yeast cells, meaning that the toxicity studies would not be hampered by the uneven distribution of oleandrin within the yeast growth media. We further checked the growth of S. cerevisiae cells in liquid media supplemented with various concentrations of oleandrin (Figure 1b). It was noted that the presence of oleandrin in the incubation medium affected cell proliferation; the half minimal inhibition concentration calculated after 16 h of incubation was IC50 = (99.57 ± 1.25) ng/mL which corresponds to a molar concentration of approximately 0.16 µM.
As the presence of oleandrin in the incubation media affected yeast cell growth, we sought to identify molecular targets of oleandrin toxicity. Considering the oleandrin structure (a glycoside containing a steroid aglycone, Figure 1a), it is highly probable that it primarily interacts with components of the plasma membrane.
Starting from the known facts that in mammalian cells oleandrin: (1) interacts with Na+/K+-ATPase; (2) induces cell Ca2+ elevations; (3) inhibits Cu+ transporter Ctr1; and (4) mediates oxidative stress by generation of reactive oxygen species (ROS), we tested the toxicity of oleandrin against yeast mutants hosting individual knockout deletions in genes related to: (1) Na+ or K+ transport across the plasma membrane; (2) Ca2+ transport and homeostasis; (3) heavy metal transporters; and (4) response to oxidative stress. The knockout genes were selected based on a search in the Saccharomyces Genome Database (SGD, [52]) and are presented in Table S1. The screening of oleandrin toxicity against the yeast knockout strains was done by exposing cells to an oleandrin concentration which caused approximately half inhibition of wild type growth (i.e., 100 ng/mL). The strains which significantly grew better or worse than the wild type in the presence of oleandrin were selected for further investigation. As shown in Table S1, no mutant with defects in Na+ or K+ transport showed a different phenotype from the wild type. From the group of mutants with defects in calcium transport, cch1Δ and mid1Δ grew better, while mutants pmc1Δ and pmr1Δ were more sensitive to oleandrin than the wild type cells (Table S1, Figure 1c).
From the group of mutants with defects in essential metal transport, only smf1Δ cells grew considerably better in the presence of oleandrin (Table S1, Figure 1c). In the case of the mutants defective in the response to oxidative stress, solely ahp1Δ grew differently in the presence of oleandrin, being more sensitive than the wild type cells (Table S1, Figure 1c, bottom).
2.2. Oleandrin Induces Calcium Influx via Cch1/Mid1
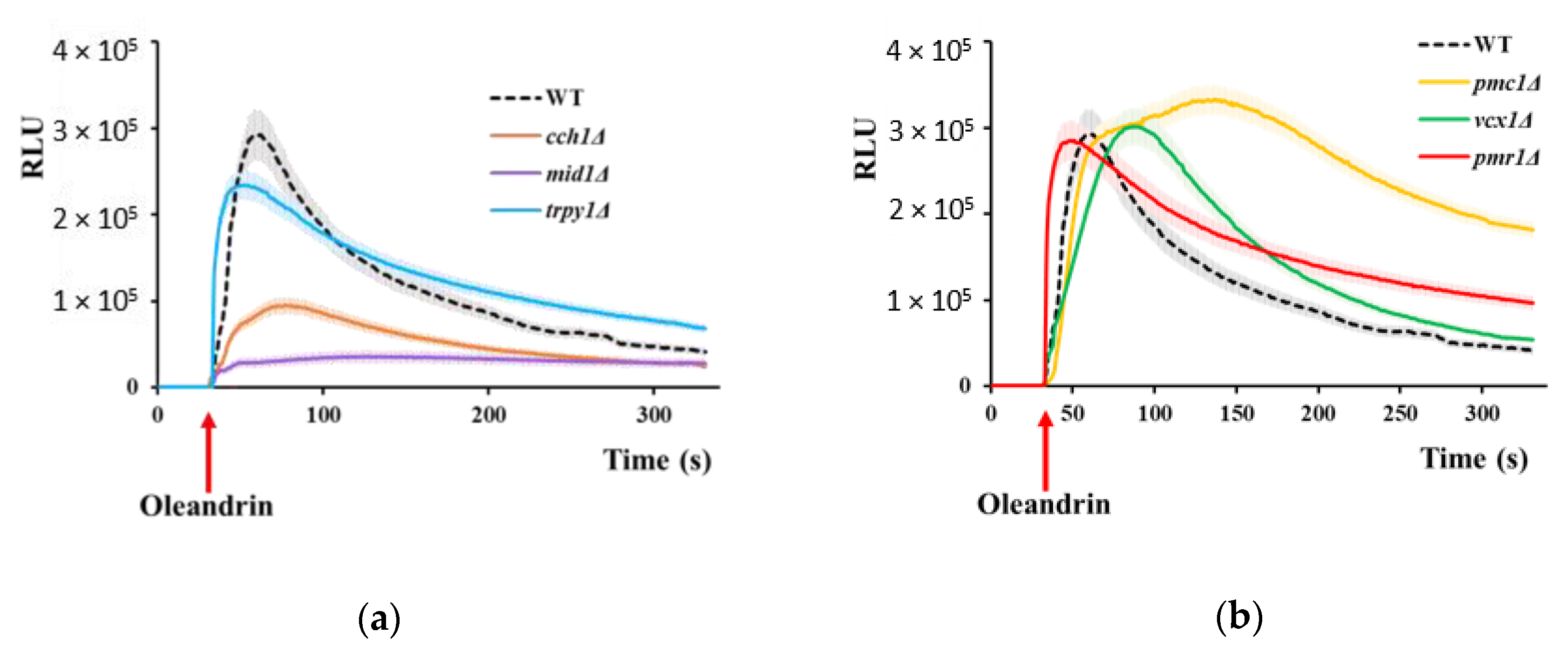
The observation that both cch1Δ and mid1Δ were more tolerant to oleandrin than the wild type cells suggested the idea that oleandrin cytotoxicity is mediated by calcium influx, which in S. cerevisiae occurs primarily via the Cch1/Mid1 channel. To test this possibility, we used transgenic yeast cells expressing aequorin, a photoprotein whose luminescence varies as a function of calcium fluctuations; the aequorin-based system used is suitable for detecting transient modifications in [Ca2+]cyt [53]. For this purpose, wild type cells, as well as cells with defects in calcium transporters (cch1Δ, mid1Δ, pmc1Δ, vcx1Δ, pmr1Δ and trpy1Δ) were transformed with a plasmid harboring the cDNA of apo-aequorin under the control of a constitutive promoter which afforded an abundant expression of aeqorin in the cytosol [54]. Before oleandrin exposure, the cells expressing apo-aequorin were pre-treated with its cofactor coelenterazine to reconstitute the functional aequorin. The cells expressing functional aequorin were stimulated with oleandrin directly in the luminometer tube. It was noted that the luminescence of wild type cells expressing functional aequorin significantly increased when cells were exposed to half-inhibitory concentrations of oleandrin (100 ng/mL), an indication of the [Ca2+]cyt elevation induced by oleandrin shock (Figure 2a).
The onset of [Ca2+]cyt elevation coincided with oleandrin addition and it took 30–40 s before luminescence started to decrease, to reach the basal low level after approximately 300 s (Figure 2a, black dashed line). [Ca2+]cyt elevation occurs when Ca2+ enters the cell from outside via the Cch1/Mid1 channel or is released from the vacuole via the TRPY1 channel (or both). Apparently, the oleandrin-dependent rise in [Ca2+]cyt was predominantly of external origin, since cch1Δ or mid1Δ cells expressing functional aequorin exhibited significantly lower oleandrin-dependent [Ca2+]cyt elevation (Figure 2a, brown and purple line, respectively), while trpy1Δ cells expressing aequorin showed high oleandrin-induced luminescence (Figure 2a, blue line).
Since cch1Δ and mid1Δ were more tolerant to oleandrin than the wild type, it can be speculated that [Ca2+]cyt elevations mediate oleandrin toxicity. In this line of evidence, the luminescence traces shown by the oleandrin-hypersensitive pmc1Δ had a broader pattern compared to wild type, and with no sign of restoring the basal [Ca2+]cyt within the 300 s interval characteristic to the wild type (Figure 2b, orange line). This observation suggested that Pmc1 (and not Vcx1, Figure 2b, green line) is crucial for restoring the low levels of [Ca2+]cyt following an oleandrin-induced calcium wave by transporting Ca2+ to the vacuole and that high Ca2+ lingering in the cytosol of pmc1Δ cells is responsible for their hypersensitivity to oleandrin. Neither vcx1Δ (lacking the vacuolar Ca2+/H+ exchanger which transports [Ca2+]cyt back to the vacuole) nor trpy1Δ (lacking the channel which releases Ca2+ from the vacuole into the cytosol) showed higher sensitivity to oleandrin than the wild type (Table S1, Figure 1c). Although the luminescence traces of aequorin-expressing trpy1Δ and vcx1Δ were slightly different, in both cases, [Ca2+]cyt decreased significantly after 300 s from oleandrin exposure (Figure 2a, blue line and Figure 2b, green line, respectively). Surprisingly, although pmr1Δ expressing functional aequorin showed luminescence traces similar to trpy1Δ (Figure 2b, red line, compared to Figure 2a, blue line), pmr1Δ cells were hypersensitive to oleandrin (Table S1, Figure 1c).
2.3. Oleandrin Exposure Induces Manganese Accumulation
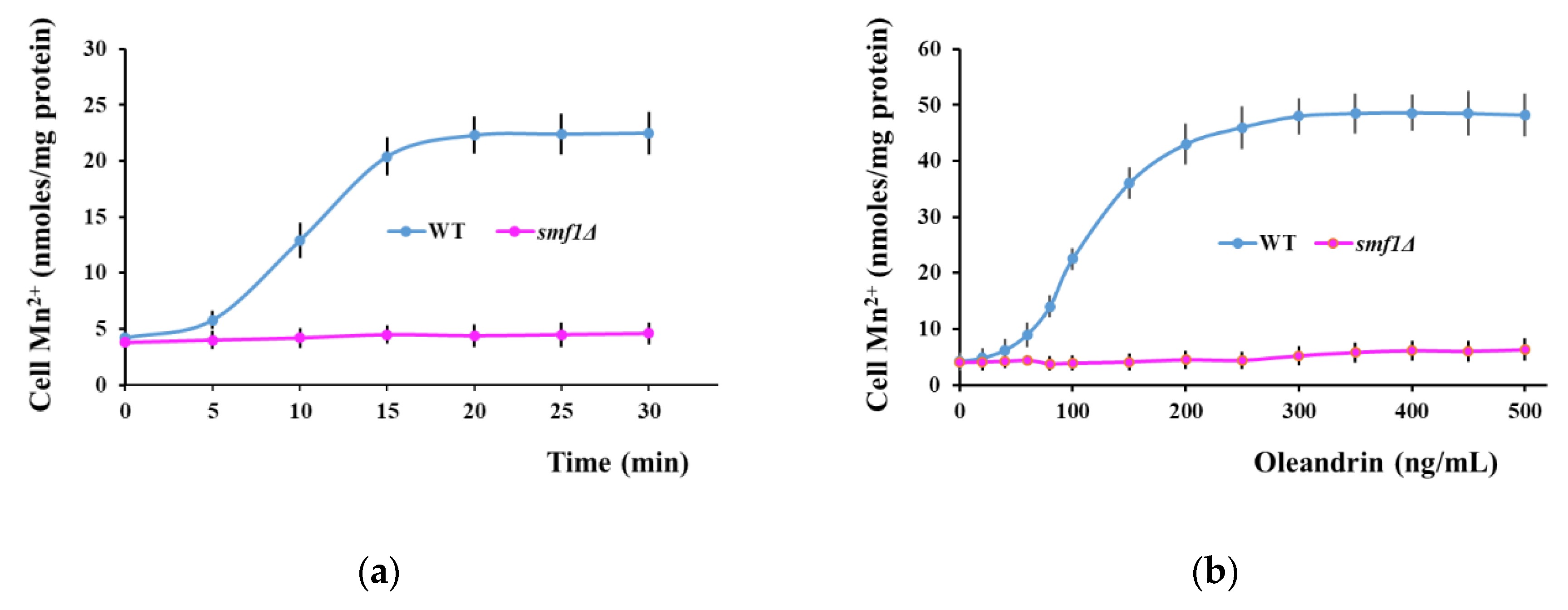
Among the yeast mutants with deletions in the genes encoding essential metal transporters, smf1Δ cells manifested increased tolerance to oleandrin. Smf1 is a divalent metal ion transporter with broad metal specificity for divalent and trivalent metals, and with high affinity for manganese [47], therefore we wondered if oleandrin exposure may be accompanied by accumulation of trace metals, eventually Smf1-dependent. To test this possibility, we performed multi-elemental analysis of yeast cells exposed to oleandrin (Table 1). To avoid inherent variation in trace metal composition of the standard media, we grew the cells in a synthetic medium (MMe) with controlled metal concentrations, containing Co2+, Cu2+, Fe3+, Mn2+, Ni2+ and Zn2+ (1 µM each). In this synthetic medium, Li+ was also added (final concentration 1 µM) as a replacement for Na+. The metal concentrations used were completely non-toxic to cells, even if they were slightly higher than in the standard media (which contain only 0.1 µM Cu2+, ultra-traces of Co2+ and Ni2+ and no Li+). It was noticed that, of all the metal ions present in the growth media, only manganese accumulation seemed to be stimulated by oleandrin (Table 1).
To check Smf1 involvement, we monitored Mn2+ accumulation by wild type and smf1Δ cells exposed to 100 ng/mL oleandrin. It was noted that in wild type cells, Mn2+ accumulation was induced by oleandrin and increased progressively in the first 5–15 min of exposure; after that, Mn2+ accumulation reached a stationary phase (Figure 3a, blue line). No oleandrin-induced Mn2+ accumulation could be detected in smf1Δ cells, indicating that oleandrin stimulated Mn2+ accumulation via the Smf1 transporter (Figure 3a,b purple lines). The Mn2+ accumulation was dose-dependent, as Mn2+ accumulation increased with oleandrin concentration, to reach a plateau when cells were exposed to oleandrin concentrations higher than 200 ng/mL (Figure 3b, blue line).
2.4. Oleandrin Hypersensitivity of Mutants pmr1Δ and ahp1Δ Is Caused by Mn2+ Accumulation
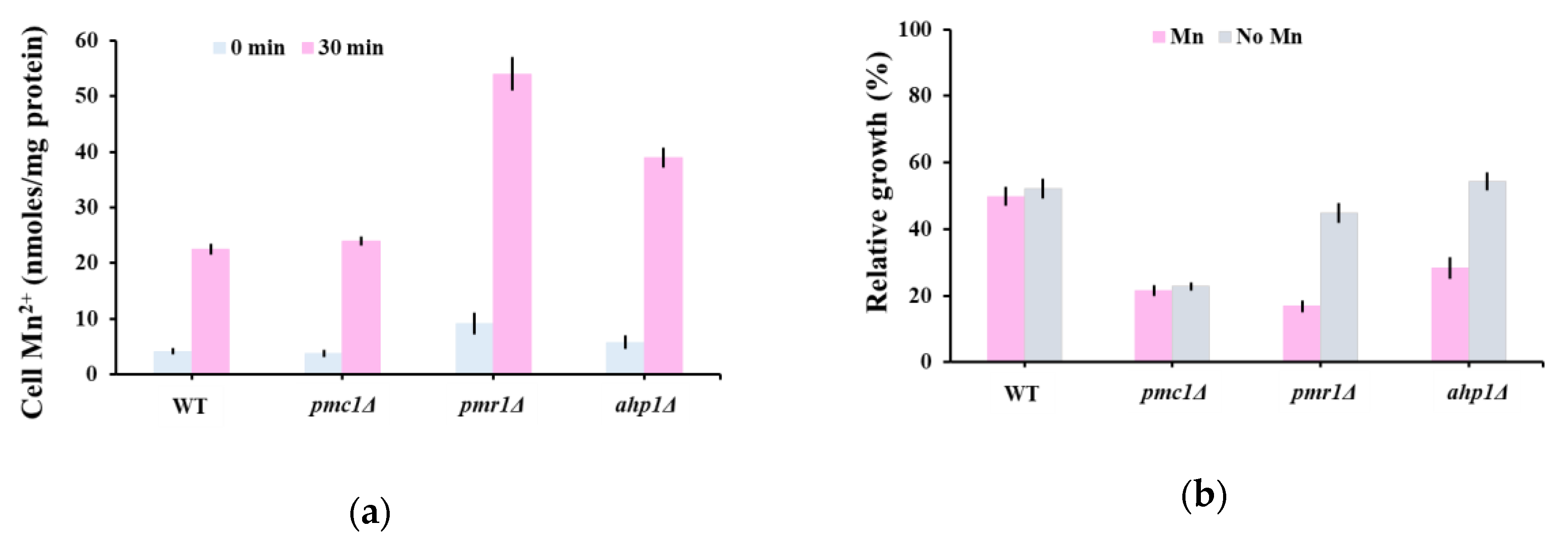
The oleandrin hypersensitivity of pmr1Δ (Figure 1c, Table S1) could not be explained by the calcium cytosolic wave that followed the oleandrin shock on aequorin expressing pmr1Δ (Figure 2b, red line) and had a similar pattern with that of trpy1Δ (Figure 2a, blue line), whose oleandrin sensitivity was similar to that of wild type cells (Figure 1c, Table S1). Pmr1 is a high-affinity Ca2+/Mn2+ P-type ATPase involved in Ca2+ and Mn2+ transport into Golgi; further, via the secretory pathway, excess Mn2+ is extruded from the cell, which is a major route for yeast cells of Mn2+ detoxification [40,55]. We therefore wondered if pmr1Δ hypersensitivity to oleandrin is related to cells’ incapacity to excrete the excess Mn2+ which occurs during oleandrin exposure, rather than to a defect in Ca2+ homeostasis. Indeed, it was noted that pmr1Δ accumulated significantly more Mn2+ than the wild type cells (Figure 4a). Instead, pmc1Δ cells which exhibited oleandrin hypersensitivity similarly to pmr1Δ displayed Mn2+ accumulation which was not significantly different from the wild type (Figure 4a), supporting the idea that pmc1Δ oleandrin hypersensitivity is caused by the incapacity of pmc1Δ cells to reduce the high [Ca2+]cyt levels in due time (Figure 2b).
It was noted that from the group of mutants with defects in the response to antioxidant stress, only ahp1Δ exhibited altered oleandrin sensitivity compared to wild type cells. Ahp1 is a thiol-specific peroxiredoxin that reduces hydroperoxides to protect against oxidative damage [56] and that also has a minor role in Mn2+ intracellular trafficking [57]. We found that ahp1Δ also accumulated more Mn2+ than the wild type in the presence of oleandrin (Figure 4a), thus explaining the ahp1Δ hypersensitivity. To check that the increased toxicity of oleandrin to pmr1Δ and ahp1Δ mutants is caused by the increased influx of external Mn2+, we determined the relative growth of yeast cells in a Mn2+-depleted medium. Indeed, we found that Mn2+ depletion improved the growth of pmr1Δ and ahp1Δ (Figure 4b), indicating that increased Mn2+ influx is responsible for the oleandrin hypersensitivity of these mutants. In this line of evidence, the oleandrin sensitivity of ahp1Δ was not alleviated by antioxidants known to improve ahp1Δ growth, such as ascorbate or tocopherol (data not shown).
3. Discussion
Apart from being a cardiac glycoside, oleandrin has been gathering attention due to its anti-tumoral [12,13,14,15,16] and antiviral potential [8,17,18,19], but its pharmacological use is held back by its variable toxicity. In this study, using the model S. cerevisiae, we detected oleandrin-induced fluctuations in cell Ca2+, which could be related to the Ca2+ entry via the Cch1/Mid1 plasma membrane channel (Figure 5).
At this point, it is hard to determine whether the Ca2+ influx was the result of the oleandrin interaction with the membrane transport of monovalent ions, since no yeast mutant with defective Na+ or K+ transport showed any alteration in oleandrin-mediated toxicity. Although the direct action of oleandrin on the Ena1 Na+/K+-ATPase cannot be ruled out since ENA1 is expressed only under salt stress conditions [26], it is highly possible that calcium influx is also triggered by a non-specific interaction of oleandrin with the yeast plasma membrane. In this line of evidence, cch1Δ cells, which still have a functional Mid1, showed some calcium influx into the cytosol (Figure 2a). Since Mid1 is a stretch-activated Ca2+-permeable cation channel [58], it is possible that oleandrin activates it by mechanical intercalation in the membrane phospholipid layer.
What raised our interest was the unexpected role of oleandrin in stimulating Mn2+ accumulation by yeast cells via the transporter Smf1. Mn2+ is an essential trace metal that serves as a cofactor for several enzymes [59], which becomes toxic when its concentration surpasses the physiological limits. In yeast, it was shown that divalent metal transport and toxicity can be manipulated by addition of natural compounds such as amino acids or polyphenols [60,61], therefore it would be interesting to screen for synergies between oleandrin and other natural compounds.
It has been suggested that the oleandrin toxicity against certain tumoral cell lines may be the result of ROS generation, especially superoxide ion radicals [21]. Nevertheless, we found that yeast mutants sod1Δ (lacking the cytosolic Cu/Zn-superoxide dismutase SOD1) or sod2Δ (lacking the mitochondrial Mn-superoxide dismutase SOD2) showed no increased oleandrin sensitivity compared to the wild type cells (Table S1). It is known that supplementary Mn2+ can suppress the oxidative damage in yeast cells lacking superoxide dismutase due to the intrinsic superoxide scavenger activity [62], therefore it is possible that in the yeast SOD mutants, the oleandrin-generated superoxide toxicity was counterbalanced by the oleandrin-induced Mn2+ influx via Smf1. Either way, it became clear that the effect of oleandrin on the eukaryotic cell may be a multi-facets process, with molecular aspects still waiting to be unraveled.
Oleandrin is of pharmacological interest due to its potential antiviral or anti-tumoral actions; it is tempting to speculate that oleandrin toxicity could be tuned by changing the Mn2+ microenvironment of the virus-infected cells or of the tumoral cells. In S. cerevisiae, oleandrin stimulated the Smf1-dependent Mn2+ influx. Smf1 is a member of the natural resistance-associated macrophage protein (NRAMP) family of transporters which includes the human counterparts DMT1 [63] and NRAMP1 [64] transporters. As NRAMP1 polymorphism has been associated with some types of cancer [65,66], it would be worthwhile studying the oleandrin action on these type of cancer in correlation with Mn2+ homeostasis, considering that Mn2+ alone was found to inhibit the viability of cancer cells [67,68].
4. Materials and Methods
4.1. Yeast Strains and Cultivation Media
The S. cerevisiae diploid strains used in this study were isogenic with the wild type (WT) parental strain BY4741 (MATa his3Δ1; leu2Δ0; ura3Δ0), a S288C-based yeast strain [69]. The knockout strains used harbored individual deletions in YFG (Your Favourite Gene) and had the genotype (BY4741, yfg::kanMX4/ORF), being denoted throughout the manuscript as yfgΔ. The strains are presented in Table S1 and they were obtained from EUROSCARF (European S. cerevisiae Archive for Functional Analysis). Yeast strains were propagated, grown and maintained in YPD medium (1% w/v yeast extract, 2% w/v polypeptone, 2% w/v glucose) or SD (synthetic dextrose, 0.17% w/v yeast nitrogen base without amino acids, 0.5% w/v (NH4)2SO4, 2% w/v glucose, supplemented with the necessary amino acids) [70]. The strains transformed with the plasmids harboring apoaequorin cDNA [53] were selected and maintained on SD lacking uracil (SD-Ura). For luminescence detection, cells were suspended in SD-Ura supplemented with 2 mM CaCl2. Minimal defined media containing known concentrations of metal ions (MMe) were prepared adding individual components as described [70] using ultrapure reagents and contained 1 µM of CoCl2; CuCl2; FeCl3; MnCl2; NiCl2; ZnCl2; and LiCl. The concentrations of metals in MMe were confirmed by inductively coupled plasma mass spectrometry (ICP-MS) (Perkin-Elmer ELAN DRC-e, Concord, ON, Canada). All synthetic media had their pH adjusted to 6.5. For solid media, 2% agar was used. For growth improvement, all the synthetic media were supplemented with an extra 20 mg/L leucine [71]. All chemicals, including media reagents, were from Merck (Darmstadt, Germany). Oleandrin was from Sigma-Aldrich (St. Louis, MO, USA) (Catalog O9640, discontinued) and was of ≥ 98% purity.
4.2. Plasmid and Yeast Transformation
For heterologous expression of apo-aequorin, yeast strains were transformed with the multicopy URA3-based plasmid pYX212-cytAEQ harboring the apoaequorin cDNA under the control of the strong TPI yeast promoter [54]. Plasmid pYX212-cytAEQ was a generous gift from E. Martegani and R. Tisi (University of Milano-Bicocca, Milan, Italy). Yeast transformation [72] was performed using S.c. EasyComp™ Transformation Kit (Invitrogen, Catalog number: K505001) following the manufacturer’s indications.
4.3. Detection of Oleandrin Effect on Yeast Cell Growth
Wild type and yfgΔ cells were inoculated from YPD-exponentially growing cells to SD liquid medium (at OD600 = 0.05) containing various concentrations of oleandrin added from a 10 mg/mL ethanol stock. Strain growth was monitored in time by measuring the turbidity of cell cultures at 600 nm (OD600) recorded in a plate reader equipped with a thermostat and shaker (Varioskan, Thermo Fisher Scientific, Vantaa, Finland). For growth on solid medium, exponentially growing cells (OD600 = 0.5) were decimally serially diluted in a multiwell plate and stamped on SD/agar plates containing oleandrin (added after medium sterilization) using a pin replicator (approximately 4 µL/spot). Plates were incubated at 30 °C for 3 days before being photographed.
4.4. Detection of [Ca2+]cyt by Recording Aequorin Luminescence
Cells transformed with pYX212-cytAEQ [54] were maintained on SD-Ura selective medium and prepared for Ca2+-dependent luminescence detection as described [73], with slight modifications. Exponentially growing yeast cells expressing apo-aequorin were diluted (OD600 = 0.5) in SD-Ura and then incubated to OD600 = 1. Cells were concentrated by centrifugation to OD600 = 10. To reconstitute functional aequorin, native coelenterazine was added to the cell suspension (from a methanol stock, 20 µM final concentration) and the cells were incubated for 2 h at 30 °C in the dark. Cells were washed to remove the excess coelenterazine and re-suspended in SD-Ura supplemented with 2 mM CaCl2. The cells were transferred (approximately 107 cells/determination) to the luminometer tube and a cellular luminescence baseline was determined for each strain by approximately one minute of recordings at 1/s intervals. After ensuring a stable signal, oleandrin was injected (v/v) from a sterile 200 ng/mL solution in SD-Ura medium, to give the final oleandrin concentration 100 ng/mL (approximately corresponding to half minimal inhibitory concentration, IC50). The Ca2+-dependent light emission was monitored in a single-tube luminometer (Turner Biosystems, 20n/20, Sunnyvale, CA, USA). The light emission was measured at 1 s intervals and expressed as relative luminescence units (RLU). To ensure that the total reconstituted aequorin was not limiting in our assay, at the end of each experiment, aequorin activity was checked by lysing cells with 1% Triton X-100 with 5 mM CaCl2 and only the cells with considerable residual luminescence were considered. Relative luminescence emission was normalized to an aequorin content giving a total light emission of 106 RLUs in 10 min after lysing cells with 1% Triton X-100.
4.5. Multielemental Analysis of Yeast Cells
Metal accumulation by cells was done as described [74], with slight modifications. Exponentially YPD-growing cells were washed and suspended in MMe liquid medium to OD600 = 0.5 in the absence or presence of oleandrin (100 ng/mL). The cells were incubated with shaking (200 rpm) for 16 h at 30 °C before they were harvested and washed three times with 10 mM 2-(N-morpholino)ethanesulfonic acid (MES)-Tris buffer, pH 6.0. Cells were finally suspended in deionized water (108 cells/mL) and used for both metal and cell protein assays. Metal detection was done using an instrument with a single collector, quadrupole inductively coupled plasma with mass spectrometry (ICP-MS) with axial field technology for trace elements, rare earth elements and isotopic analyses. Metal analyses were performed after digestion of cells with 65% ultrapure HNO3 (Merck). Standard solutions were prepared by diluting a 10 µg/mL multielement solution (Multielement ICP Calibration Standard 3, matrix 5%HNO3, Perkin Elmer Pure Plus, Shelton, CT, USA). The metal cellular content was normalized to total cellular proteins, which were assayed spectrophotometrically [75].
4.6. Statistics
All experiments were repeated, independently, in three biological replicates at least. For each individual experiment, values were expressed as the mean ± standard error of the mean (SEM). For aequorin luminescence determinations, traces represent the mean (±SEM) from three independent transformants. The numerical data were examined by Student t test or by analysis of variance with multiple comparisons (ANOVA) using the statistical software Prism version 6.05 for Windows (GraphPad Software, La Jolla, CA, USA). The differences were considered to be significant when p < 0.05. One sample t test was used for the statistical analysis of each strain/condition compared with a strain/condition considered as reference. Asterisks indicate the level of significance: * p < 0.05, ** p < 0.01 and *** p < 0.001.
5. Conclusions
Oleandrin toxicity against eukaryotic cells was investigated using the model microorganism S. cerevisiae. We found that exposing yeast cells to oleandrin resulted in Ca2+ influx into the cytosol and that defects in restoring the normal level of cytosolic Ca2+ (e.g., by pumping excess cytosolic Ca2+ to the vacuole) augmented the oleandrin toxicity. We also found that oleandrin exposure induced Mn2+ accumulation by the yeast cells via the plasma membrane Smf1 and that mutants with defects in Mn2+ homeostasis may become oleandrin-hypersensitive. Our data suggest that combining oleandrin with agents which alter Ca2+ or Mn2+ homeostasis may be a way of scope-tuning oleandrin toxicity.
Supplementary Materials
The following are available online, Table S1: Effect of oleandrin on the growth of various single-gene knockout mutants.
Author Contributions
Conceptualization, I.C.F.; Methodology, L.L.R., C.V.P. and I.C.F; Validation, L.L.R., C.V.P. and I.C.F; Formal analysis, I.C.F.; L.L.R., C.V.P. and I.C.F; Data curation, L.L.R. and C.V.P; Writing—original draft preparation, I.C.F.; Writing—review and editing, I.C.F.; Supervision, I.C.F.; Project administration, L.L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Enzo Martegani and Renata Tisi (from University of Milano-Bicocca, Milan, Italy) for providing the plasmid pYX212-cytAEQ, Andrei F. Danet for technical support and Aurora D. Neagoe for ICP-MS analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dey, P. The pharmaco-toxicological conundrum of oleander: Potential role of gutmicrobiome. Biomed. Pharmacother. 2020, 129, 110422. [Google Scholar] [CrossRef] [PubMed]
- Langford, S.D.; Boor, P.J. Oleander toxicity: An examination of human and animal toxic exposures. Toxicology 1996, 109, 1–13. [Google Scholar] [CrossRef]
- Karthik, G.; Iyadurai, R.; Ralph, R.; Prakash, V.; Abhilash, K.P.P.; Sathyendra, S.; Abraham, O.C.; Truman, C.; Reginald, A. Acute oleander poisoning: A study of clinical profile from a tertiary care center in South India. J. Fam. Med. Prim. Care 2020, 9, 136–140. [Google Scholar] [CrossRef]
- Botelho, A.F.M.; Santos-Miranda, A.; Joca, H.C.; Mattoso, C.R.S.; de Oliveira, M.S.; Pierezan, F.; Cruz, J.S.; Soto-Blanco, B.; Melo, M.M. Hydroalcoholic extract from Nerium oleander L. (Apocynaceae) elicits arrhythmogenic activity. J. Ethnopharmacol. 2017, 206, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Rashan, L.J.; Franke, K.; Khine, M.M.; Kelter, G.; Fiebig, H.H.; Neumann, J.; Wessjohann, L.A. Characterization of the anticancer properties of monoglycosidic cardenolides isolated from Nerium oleander and Streptocaulon tomentosum. J. Ethnopharmacol. 2011, 134, 781–788. [Google Scholar] [CrossRef]
- Cao, Y.L.; Zhang, M.H.; Lu, Y.F.; Li, C.Y.; Tang, J.S.; Jiang, M.M. Cardenolides from the leaves of Nerium oleander. Fitoterapia 2018, 127, 293–300. [Google Scholar] [CrossRef]
- Botelho, A.F.M.; Pierezan, F.; Soto-Blanco, B.; Melo, M.M. A review of cardiac glycosides: Structure, toxicokinetics, clinical signs, diagnosis and antineoplastic potential. Toxicon 2019, 158, 63–68. [Google Scholar] [CrossRef]
- Hutchison, T.; Yapindi, L.; Malu, A.; Newman, R.A.; Sastry, K.J.; Harrod, R. The botanical glycoside oleandrin inhibits human T-cell leukemia virus type-1 infectivity and Env-dependent virological synapse formation. J. Antivir. Antiretrovir. 2019, 11, 184. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11541511 (accessed on 10 August 2020).
- Lin, Y.; Ho, D.H.; Newman, R.A. Human tumor cell sensitivity to oleandrin is dependent on relative expression of Na+,K+-ATPase subunits. J. Exp. Ther. Oncol. 2010, 8, 271–286. [Google Scholar]
- Botelho, A.F.M.; Miranda, A.L.S.; Freitas, T.G.; Milani, P.F.; Barreto, T.; Cruz, J.S.; Melo, M.M. Comparative cardiotoxicity of low doses of digoxin, ouabain, and oleandrin. Cardiovasc. Toxicol. 2020. [Google Scholar] [CrossRef]
- Bao, Z.; Tian, B.; Wang, X.; Feng, H.; Liang, Y.; Chen, Z.; Li, W.; Shen, H.; Ying, S. Oleandrin induces DNA damage responses in cancer cells by suppressing the expression of Rad51. Oncotarget 2016, 7, 59572–59579. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, Y.; Zhao, W.; Zhou, X.; Wang, C.; Deng, F. The cardiac glycoside oleandrin induces apoptosis in human colon cancer cells via the mitochondrial pathway. Cancer Chemother. Pharmacol. 2017, 80, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.S.; Rugira, T.; Jin, H.; Park, S.W.; Kim, H.J. Oleandrin and its derivative odoroside a, both cardiac glycosides, exhibit anticancer effects by inhibiting invasion via suppressing the STAT-3 signaling pathway. Int. J. Mol. Sci. 2018, 19, 3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, L.; Ma, Y.; Liang, C.; He, G.; Zhao, Z.; Yang, C.; Hai, B.; Pan, X.; Liu, Z.; Liu, X. Oleandrin sensitizes human osteosarcoma cells to cisplatin by preventing degradation of the copper transporter 1. Phytother. Res. 2019, 33, 1837–1850. [Google Scholar] [CrossRef]
- Li, X.X.; Wang, D.Q.; Sui, C.G.; Meng, F.D.; Sun, S.L.; Zheng, J.; Jiang, Y.H. Oleandrin induces apoptosis via activating endoplasmic reticulum stress in breast cancer cells. Biomed. Pharmacother. 2020, 124, 109852. [Google Scholar] [CrossRef]
- Singh, S.; Shenoy, S.; Nehete, P.N.; Yang, P.; Nehete, B.; Fontenot, D.; Yang, G.; Newman, R.A.; Sastry, K.J. Nerium oleander derived cardiac glycoside oleandrin is a novel inhibitor of HIV infectivity. Fitoterapia 2013, 84, 32–39. [Google Scholar] [CrossRef]
- Yang, C.W.; Chang, H.Y.; Hsu, H.Y.; Lee, Y.Z.; Chang, H.S.; Chen, I.S.; Lee, S.J. Identification of anti-viral activity of the cardenolides, Na(+)/K(+)-ATPase inhibitors, against porcine transmissible gastroenteritis virus. Toxicol. Appl. Pharmacol. 2017, 332, 129–137. [Google Scholar] [CrossRef]
- Plante, K.S.; Plante, J.A.; Fernandez, D.; Mirchandani, D.; Bopp, N.; Aguilar, P.V.; Sastry, K.J.; Newman, R.A.; Weaver, S.C. Prophylactic and therapeutic inhibition of in vitro SARS-CoV-2 replication by Oleandrin. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kanwal, N.; Rasul, A.; Hussain, G.; Anwar, H.; Shah, M.A.; Sarfraz, I.; Riaz, A.; Batool, R.; Shahba, M.; Hussain, A.; et al. Oleandrin: A bioactive phytochemical and potential cancer killer via multiple cellular signaling pathway. Food Chem. Toxicol. 2020, 143, 111570. [Google Scholar] [CrossRef]
- Newman, R.A.; Yang, P.; Hittelman, W.N.; Lu, T.; Ho, D.H.; Ni, D.; Chan, D.; Vijjeswarapu, M.; Cartwright, C.; Dixon, S.; et al. Oleandrin-mediated oxidative stress in human melanoma cells. J. Exp. Ther. Oncol. 2006, 5, 167–181. [Google Scholar]
- Castrillo, J.I.; Oliver, S. Yeast as a touchstone in post-genomic research: Strategies for integrative analysis in functional genomics. J. Biochem. Mol. Biol. 2004, 37, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuo, R.; Sousa, F.G.; Soares, D.G.; Bonatto, D.; Saffi, J.; Escargueil, A.E.; Larsen, A.K.; Henriques, J.A. Saccharomyces cerevisiae as a model system to study the response to anticancer agents. Cancer Chemother. Pharmacol. 2012, 70, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.C.; Sá-Correia, I. Yeast toxicogenomics: Lessons from a eukaryotic cell model and cell factory. Curr. Opin. Biotechnol. 2015, 33, 183–191. [Google Scholar] [CrossRef]
- Manna, S.K.; Sah, N.K.; Newman, A.; Cisneros, A.; Aggarwal, B.B. Oleandrin suppresses activation of nuclear transcription factor-kB, activator protein-1, and c-Jun N-terminal kinase. Cancer Res. 2000, 60, 3838–3847. [Google Scholar] [PubMed]
- Ariño, J.; Ramos, J.; Sychrova, H. Monovalent cation transporters at the plasma membrane in yeasts. Yeast 2019, 36, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.K.; Ellsmore, A.J.; Cessna, S.G.; Low, P.S.; Pardo, J.M.; Bressan, R.A.; Hasegawa, P.M. An osmotically induced cytosolic Ca2+ transient activates calcineurin signaling to mediate ion homeostasis and salt tolerance of Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 33075–33080. [Google Scholar] [CrossRef] [Green Version]
- Viladevall, L.; Serrano, R.; Ruiz, A.; Domenech, G.; Giraldo, J.; Barceló, A.; Ariño, J. Characterization of the calcium-mediated response to alkaline stress in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 43614–43624. [Google Scholar] [CrossRef] [Green Version]
- Bootman, M.D.; Berridge, M.J.; Putney, J.W.; Roderick, H.L. Calcium Signaling; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2011; 449p, ISBN 978-0-87969-903-1. [Google Scholar]
- Batiza, A.F.; Schulz, T.; Masson, P.H. Yeast respond to hypotonic shock with a calcium pulse. J. Biol. Chem. 1996, 271, 23357–23362. [Google Scholar] [CrossRef] [Green Version]
- Denis, V.; Cyert, M.S. Internal Ca(2+) release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J. Cell Biol. 2002, 156, 29–34. [Google Scholar] [CrossRef]
- Palmer, C.P.; Zhou, X.; Lin, J.; Loukin, S.H.; Kung, C.; Saimi, Y. A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca(2+)-permeable channel in the yeast vacuolar membrane. Proc. Natl. Acad. Sci. USA 2001, 98, 7801–7805. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, K.W. Acidic calcium stores of Saccharomyces cerevisiae. Cell Calcium 2011, 50, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paidhungat, M.; Garrett, S. A homolog of mammalian, voltage-gated calcium channels mediates yeast pheromone-stimulated Ca2+ uptake and exacerbates the cdc1(Ts) growth defect. Mol. Cell. Biol. 1997, 17, 6339–6347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, K.; Teng, J.; Cho, T.; Yoshikawa-Kimura, S.; Iida, H. Post-translational processing and membrane translocation of the yeast regulatory Mid1 subunit of the Cch1/VGCC/NALCN cation channel family. J. Biol. Chem. 2017, 292, 20570–20582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, K.W.; Fink, G.R. Calcineurin-dependent growth control in Saccharomyces cerevisiae mutants lacking PMC1, a homolog of plasma membrane Ca2+ ATPases. J. Cell Biol. 1994, 124, 351–363. [Google Scholar] [CrossRef]
- Cunningham, K.W.; Fink, G.R. Calcineurin inhibits VCX1-dependent H+/Ca2+ exchange and induces Ca2+ ATPases in Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 2226–2237. [Google Scholar] [CrossRef] [Green Version]
- Miseta, A.; Kellermayer, R.; Aiello, D.P.; Fu, L.; Bedwell, D.M. The vacuolar Ca2+/H+ exchanger Vcx1p/Hum1p tightly controls cytosolic Ca2+ levels in S. cerevisiae. FEBS Lett. 1999, 451, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Sorin, A.; Rosas, G.; Rao, R. PMR1, a Ca2+-ATPase in yeast Golgi, has properties distinct from sarco/endoplasmic reticulum and plasma membrane calcium pumps. J. Biol. Chem. 1997, 272, 9895–9901. [Google Scholar] [CrossRef] [Green Version]
- Dürr, G.; Strayle, J.; Plemper, R.; Elbs, S.; Klee, S.K.; Catty, P.; Wolf, D.H.; Rudolph, H.K. The medial-Golgi ion pump Pmr1 supplies the yeast secretory pathway with Ca2+ and Mn2+ required for glycosylation, sorting, and endoplasmic reticulum-associated protein degradation. Mol. Biol. Cell 1998, 9, 1149–1162. [Google Scholar] [CrossRef] [Green Version]
- Culotta, V.C.; Yang, M.; Hall, M.D. Manganese transport and trafficking: Lessons learned from Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Kellermayer, R. Hailey-Hailey disease as an orthodisease of PMR1 deficiency in Saccharomyces cerevisiae. FEBS Lett. 2005, 579, 2021–2025. [Google Scholar] [CrossRef] [Green Version]
- Van Ho, A.; Ward, D.M.; Kaplan, J. Transition metal transport in yeast. Annu. Rev. Microbiol. 2002, 56, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Dancis, A.; Haile, D.; Yuan, D.S.; Klausner, R.D. The Saccharomyces cerevisiae copper transport protein (Ctr1p). Biochemical characterization, regulation by copper, and physiologic role in copper uptake. J. Biol. Chem. 1994, 269, 25660–25667. [Google Scholar] [PubMed]
- Eide, D.J. The molecular biology of metal ion transport in Saccharomyces cerevisiae. Annu. Rev. Nutr. 1998, 18, 441–469. [Google Scholar] [CrossRef] [PubMed]
- Hassett, R.; Dix, D.R.; Eide, D.J.; Kosman, D.J. The Fe(II) permease Fet4p functions as a low affinity copper transporter and supports normal copper trafficking in Saccharomyces cerevisiae. Biochem. J. 2000, 351 Pt 2, 477–484. [Google Scholar] [CrossRef]
- Supek, F.; Supekova, L.; Nelson, H.; Nelson, N. A yeast manganese transporter related to the macrophage protein involved in conferring resistance to mycobacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 5105–5110. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.T.; Ajua-Alemanji, M.; Culotta, V.C. The Saccharomyces cerevisiae high affinity phosphate transporter encoded by PHO84 also functions in manganese homeostasis. J. Biol. Chem. 2003, 278, 42036–42040. [Google Scholar] [CrossRef] [Green Version]
- Ofiteru, A.M.; Ruta, L.L.; Rotaru, C.; Dumitru, I.; Ene, C.D.; Neagoe, A.; Farcasanu, I.C. Overexpression of the PHO84 gene causes heavy metal accumulation and induces Ire1p-dependent unfolded protein response in Saccharomyces cerevisiae cells. Appl. Microbiol. Biotechnol. 2012, 94, 425–435. [Google Scholar] [CrossRef]
- Zhao, H.; Eide, D. The yeast ZRT1 gene encodes the zinc transporter protein of a high-affinity uptake system induced by zinc limitation. Proc. Natl. Acad. Sci. USA 1996, 93, 2454–2458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Eide, D. The ZRT2 gene encodes the low affinity zinc transporter in Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 23203–23210. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.yeastgenome.org/ (accessed on 10 August 2020).
- Nakajima-Shimada, J.; Iida, H.; Tsuji, F.I.; Anraku, Y. Monitoring of intracellular calcium in Saccharomyces cerevisiae with an apoaequorine cDNA expression system. Proc. Natl. Acad. Sci. USA 1991, 88, 6878–6882. [Google Scholar] [CrossRef] [Green Version]
- Tisi, R.; Baldassa, S.; Belotti, F.; Martegani, E. Phospholipase C is required for glucose-induced calcium influx in budding yeast. FEBS Lett. 2002, 520, 133–138. [Google Scholar] [CrossRef]
- Mandal, D.; Woolf, T.B.; Rao, R. Manganese selectivity of Pmr1, the yeast secretory pathway ion pump, is defined by residue gln783 in transmembrane segment 6. Residue Asp778 is essential for cation transport. J. Biol. Chem. 2000, 275, 23933–23938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Spector, D.; Godon, C.; Labarre, J.; Toledano, M.B. A new antioxidant with alkyl hydroperoxide defense properties in yeast. J. Biol. Chem. 1999, 274, 4537–4544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farcasanu, I.C.; Hirata, D.; Tsuchiya, E.; Mizuta, K.; Miyakawa, T. Involvement of thioredoxin peroxidase type II (Ahp1p) of Saccharomyces cerevisiae in Mn2+ homeostasis. Biosci. Biotechnol. Biochem. 1999, 63, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, M.; Nagasawa, M.; Kojima, I.; Sato, C.; Naruse, K.; Sokabe, M.; Iida, H. Molecular identification of a eukaryotic, stretch-activated nonselective cation channel. Science 1999, 5429, 882–886. [Google Scholar] [CrossRef]
- Erikson, K.M.; Aschner, M. Manganese: Its role in disease and health. In Essential Metals in Medicine: Therapeutic Use and Toxicity of Metal Ions in the Clinic; Sigel, A., Freisinger, E., Sigel, R.K.O., Carver, P.L., Eds.; De Gruyter GmbH: Berlin, Germany, 2019; pp. 253–266. [Google Scholar] [CrossRef]
- Farcasanu, I.C.; Mizunuma, M.; Nishiyama, F.; Miyakawa, T. Role of L-histidine in conferring tolerance to Ni2+ in Sacchromyces cerevisiae cells. Biosci. Biotechnol. Biochem. 2005, 69, 2343–2348. [Google Scholar] [CrossRef]
- Oprea, E.; Ruta, L.L.; Nicolau, I.; Popa, C.V.; Neagoe, A.D.; Farcasanu, I.C. Vaccinium corymbosum L. (blueberry) extracts exhibit protective action against cadmium toxicity in Saccharomyces cerevisiae cells. Food Chem. 2014, 152, 516–521. [Google Scholar] [CrossRef]
- Lapinskas, P.J.; Cunningham, K.W.; Liu, X.F.; Fink, G.R.; Culotta, V.C. Mutations in PMR1 suppress oxidative damage in yeast cells lacking superoxide dismutase. Mol. Cell. Biol. 1995, 15, 1382–1388. [Google Scholar] [CrossRef] [Green Version]
- Au, C.; Benedetto, A.; Aschner, M. Manganese transport in eukaryotes: The role of DMT1. Neurotoxicology 2008, 29, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Courville, P.; Chaloupka, R.; Cellier, M.F. Recent progress in structure-function analyses of Nramp proton-dependent metal-ion transporters. Biochem. Cell Biol. 2006, 84, 960–978. [Google Scholar] [CrossRef]
- Calhoun, E.S.; McGovern, R.M.; Janney, C.A.; Cerhan, J.R.; Iturria, S.J.; Smith, D.I.; Gostout, B.S.; Persing, D.H. Host genetic polymorphism analysis in cervical cancer. Clin. Chem. 2002, 48, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Lenormand, C.; Couteau, J.; Nouhaud, F.X.; Maillet, G.; Bou, J.; Gobet, F.; Pfister, C. Predictive value of NRAMP1 and HGPX1 gene polymorphism for maintenance BCG response in non-muscle-invasive bladder cancer. Anticancer Res. 2016, 36, 1737–1743. [Google Scholar] [PubMed]
- Hernroth, B.; Holm, I.; Gondikas, A.; Tassidis, H. Manganese inhibits viability of prostate cancer cells. Anticancer Res. 2018, 38, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Doble, P.A.; Miklos, G.L.G. Distributions of manganese in diverse human cancers provide insights into tumour radioresistance. Metallomics 2018, 10, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer deletion strains derived from Saccharomyces cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- Sherman, F. Getting started with yeast. Methods Enzymol. 2002, 350, 3–41. [Google Scholar] [CrossRef]
- Cohen, R.; Engelberg, D. Commonly used Saccharomyces cerevisiae strains (e.g., BY4741, W303) are growth sensitive on synthetic complete medium due to poor leucine uptake. FEMS Microbiol. Lett. 2007, 273, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Dohmen, R.J.; Strasser, A.W.M.; Honer, C.B.; Hollenberg, C.P. An efficient transformation procedure enabling long-term storage of competent cells of various yeast genera. Yeast 1991, 7, 691–692. [Google Scholar] [CrossRef]
- Tisi, R.; Martegani, E.; Brandão, R.L. Monitoring yeast intracellular Ca2+ levels using an in vivo bioluminescence assay. Cold Spring Harb. Protoc. 2015, 2015, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Ruta, L.L.; Kissen, R.; Nicolau, I.; Neagoe, A.D.; Petrescu, A.J.; Bones, A.M.; Farcasanu, I.C. Heavy metal accumulation by Saccharomyces cerevisiae cells armed with metal binding hexapeptides targeted to the inner face of the plasma membrane. Appl. Microbiol. Biotechnol. 2017, 101, 5749–5763. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Sample Availability: Samples of the oleandrin stock solution used are available from the authors. |
Figure 1.
Effect of oleandrin on yeast growth. (a) Oleandrin structure. (b) Effect of oleandrin concentration on growth of wild type cells. BY4741 cells were inoculated (OD600 = 0.05) and grown in SD (synthetic dextrose) liquid medium in the presence of various concentrations of oleandrin. Cell density was determined spectrophotometrically at 600 nm (OD600) as described in the Materials and Methods section. One-way ANOVA, * p < 0.05; ** p < 0.01; *** p < 0.005. (c) Growth on oleandrin-supplemented solid medium. Wild type (WT) or knockout mutant cells cch1Δ, mid1Δ, pmc1Δ, smf1Δ, pmr1Δ and ahp1Δ with oleandrin sensitivity different from WT (see Table S1) were serially diluted and stamped on SD/agar containing or not 100 ng/mL oleandrin. Plates were photographed after 3 days’ incubation at 30 °C.
Figure 1.
Effect of oleandrin on yeast growth. (a) Oleandrin structure. (b) Effect of oleandrin concentration on growth of wild type cells. BY4741 cells were inoculated (OD600 = 0.05) and grown in SD (synthetic dextrose) liquid medium in the presence of various concentrations of oleandrin. Cell density was determined spectrophotometrically at 600 nm (OD600) as described in the Materials and Methods section. One-way ANOVA, * p < 0.05; ** p < 0.01; *** p < 0.005. (c) Growth on oleandrin-supplemented solid medium. Wild type (WT) or knockout mutant cells cch1Δ, mid1Δ, pmc1Δ, smf1Δ, pmr1Δ and ahp1Δ with oleandrin sensitivity different from WT (see Table S1) were serially diluted and stamped on SD/agar containing or not 100 ng/mL oleandrin. Plates were photographed after 3 days’ incubation at 30 °C.
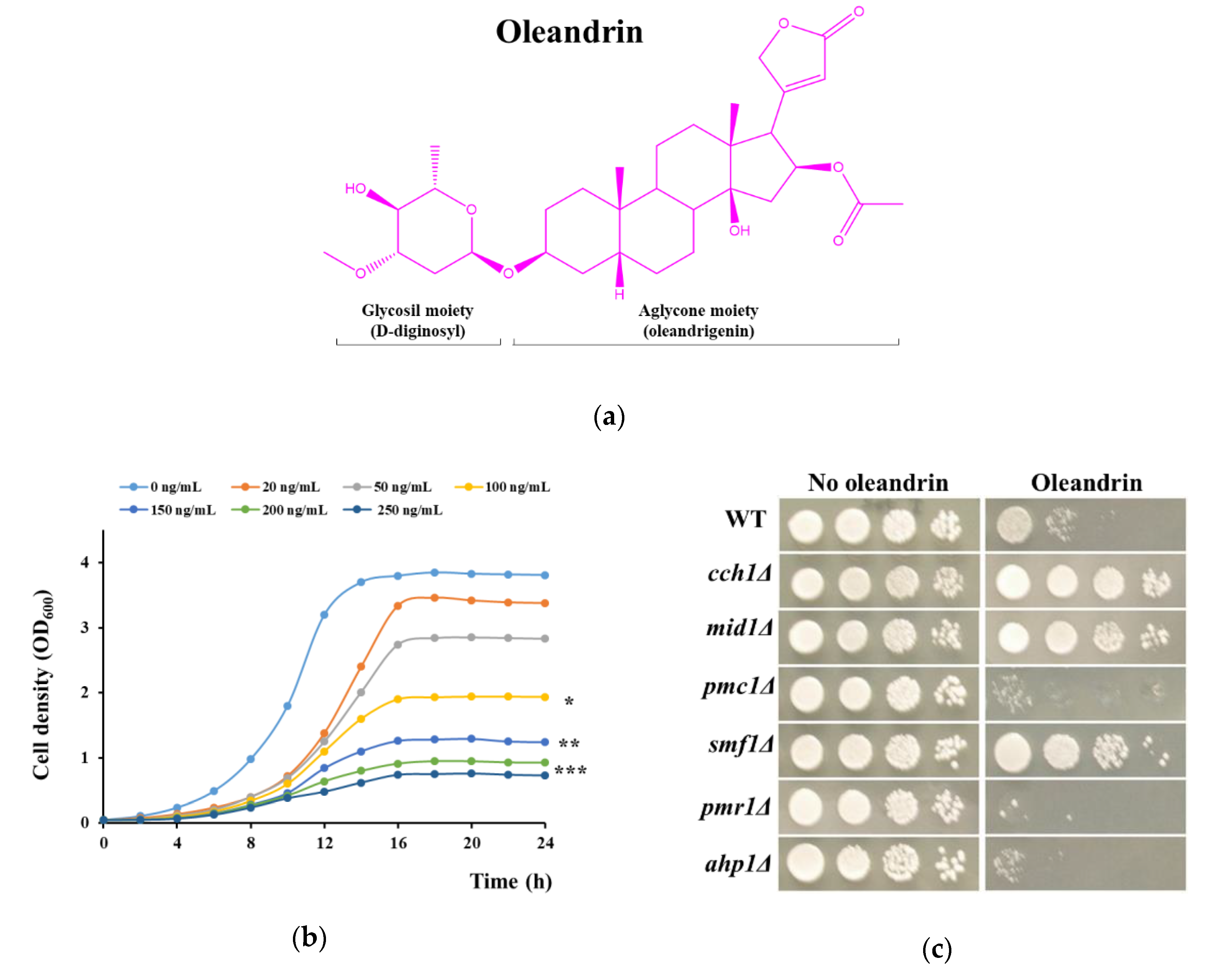
Figure 2.
Increase in [Ca2+]cyt under oleandrin exposure. Wild type cells or cells defective in calcium transport expressing reconstituted aequorin were pre-grown in SD-Ura and subjected to oleandrin stress (100 ng/mL) as described in Materials and Methods. [Ca2+]cyt-dependent aequorin luminescence was recorded on samples of approximately 107 cells (OD600 = 1). The arrow indicates the time when the oleandrin was added. (a) Calcium-dependent luminescence of wild type, and of mutants with defects in Ca2+ release to the cytosol cch1Δ, mid1Δ and trpy1Δ. (b) Calcium-dependent luminescence of type, and of mutants with defects in Ca2+ removal from cytosol pmc1Δ, vcx1Δ and pmr1Δ. The luminescence traces represent the mean ± SEM from 3 independent transformants. SEM are illustrated as bars of a lighter nuance. RLU, relative luminescence units.
Figure 2.
Increase in [Ca2+]cyt under oleandrin exposure. Wild type cells or cells defective in calcium transport expressing reconstituted aequorin were pre-grown in SD-Ura and subjected to oleandrin stress (100 ng/mL) as described in Materials and Methods. [Ca2+]cyt-dependent aequorin luminescence was recorded on samples of approximately 107 cells (OD600 = 1). The arrow indicates the time when the oleandrin was added. (a) Calcium-dependent luminescence of wild type, and of mutants with defects in Ca2+ release to the cytosol cch1Δ, mid1Δ and trpy1Δ. (b) Calcium-dependent luminescence of type, and of mutants with defects in Ca2+ removal from cytosol pmc1Δ, vcx1Δ and pmr1Δ. The luminescence traces represent the mean ± SEM from 3 independent transformants. SEM are illustrated as bars of a lighter nuance. RLU, relative luminescence units.
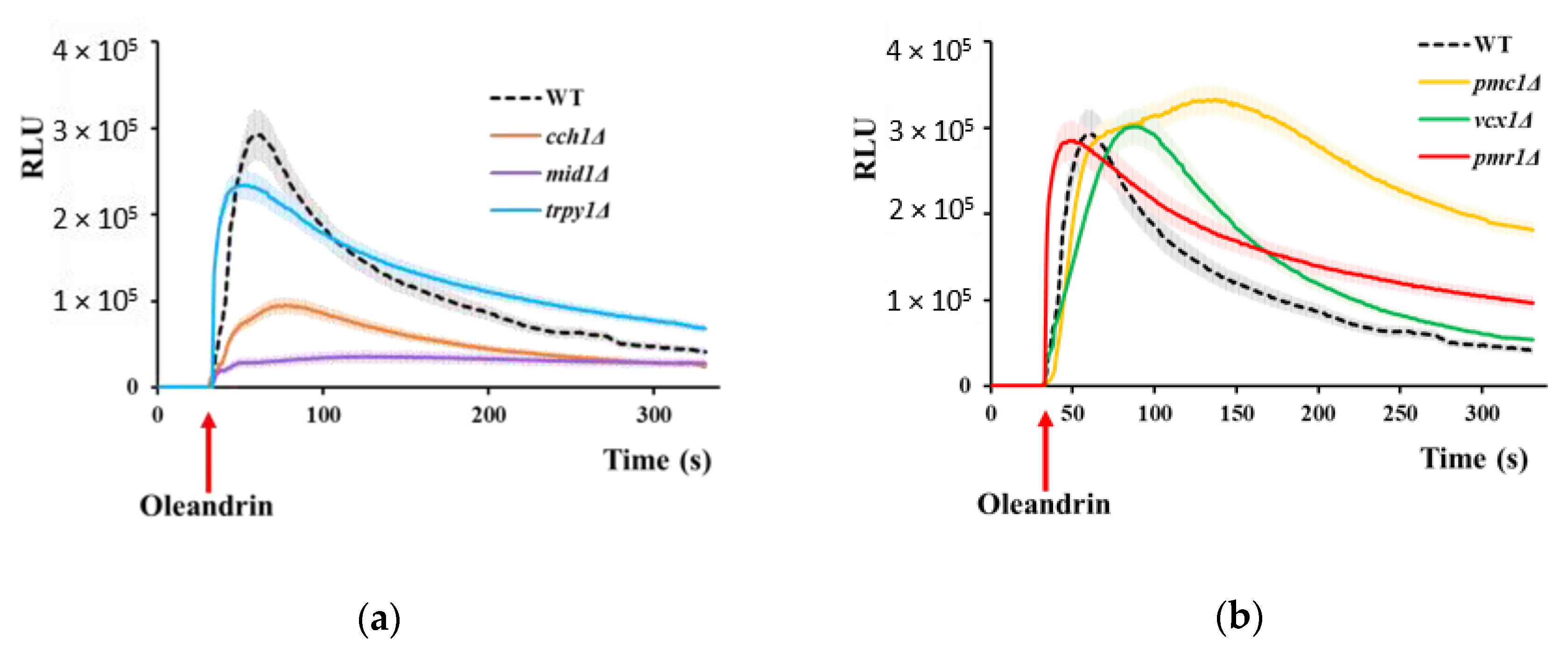
Figure 3.
Oleandrin induces Smf1-dependent Mn2+ accumulation in yeast cells. (a) Time course of Mn2+ accumulation following oleandrin exposure. Exponentially growing wild type and smf1Δ cells were treated with oleandrin (100 ng/mL) and samples were collected every 5 min for Mn2+ assay. (b) Effect of oleandrin concentrations on Mn2+ accumulation. Exponentially growing wild type and smf1Δ cells were treated with increasing oleandrin concentrations and samples were collected after 20 min for Mn2+ assay. Cells were incubated (30 °C, 200 rpm) in liquid MMe containing 1 μM MnCl2. *** p < 0.005, Student’s t test.
Figure 3.
Oleandrin induces Smf1-dependent Mn2+ accumulation in yeast cells. (a) Time course of Mn2+ accumulation following oleandrin exposure. Exponentially growing wild type and smf1Δ cells were treated with oleandrin (100 ng/mL) and samples were collected every 5 min for Mn2+ assay. (b) Effect of oleandrin concentrations on Mn2+ accumulation. Exponentially growing wild type and smf1Δ cells were treated with increasing oleandrin concentrations and samples were collected after 20 min for Mn2+ assay. Cells were incubated (30 °C, 200 rpm) in liquid MMe containing 1 μM MnCl2. *** p < 0.005, Student’s t test.
Figure 4.
(a) Mn2+ accumulation by mutants hypersensitive to oleandrin. Exponentially growing yeast cells were treated with oleandrin (100 ng/mL) in MMe containing 1 μM MnCl2 and samples were collected after 20 min for Mn2+ assay. (b) Effect of Mn2+ depletion on oleandrin toxicity. Cells were incubated (30 °C, 200 rpm) in liquid MMe containing 1 μM MnCl2 (Mn) or Mn2+-depleted (No Mn) in the presence of 100 ng/mL oleandrin. Cell growth was measured spectrophotometrically (OD600) and expressed relatively to growth under the same conditions, but in the absence of oleandrin.
Figure 4.
(a) Mn2+ accumulation by mutants hypersensitive to oleandrin. Exponentially growing yeast cells were treated with oleandrin (100 ng/mL) in MMe containing 1 μM MnCl2 and samples were collected after 20 min for Mn2+ assay. (b) Effect of Mn2+ depletion on oleandrin toxicity. Cells were incubated (30 °C, 200 rpm) in liquid MMe containing 1 μM MnCl2 (Mn) or Mn2+-depleted (No Mn) in the presence of 100 ng/mL oleandrin. Cell growth was measured spectrophotometrically (OD600) and expressed relatively to growth under the same conditions, but in the absence of oleandrin.

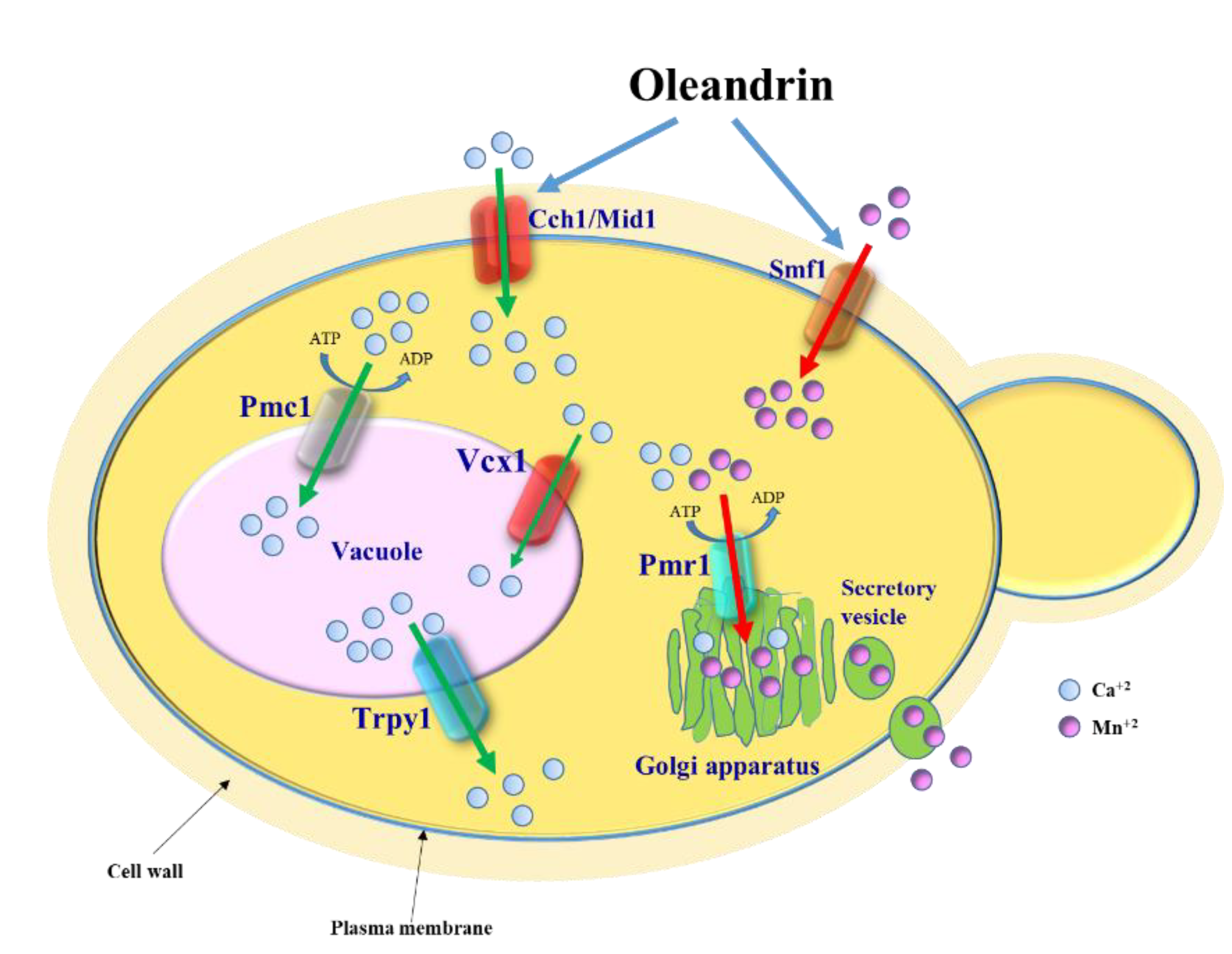
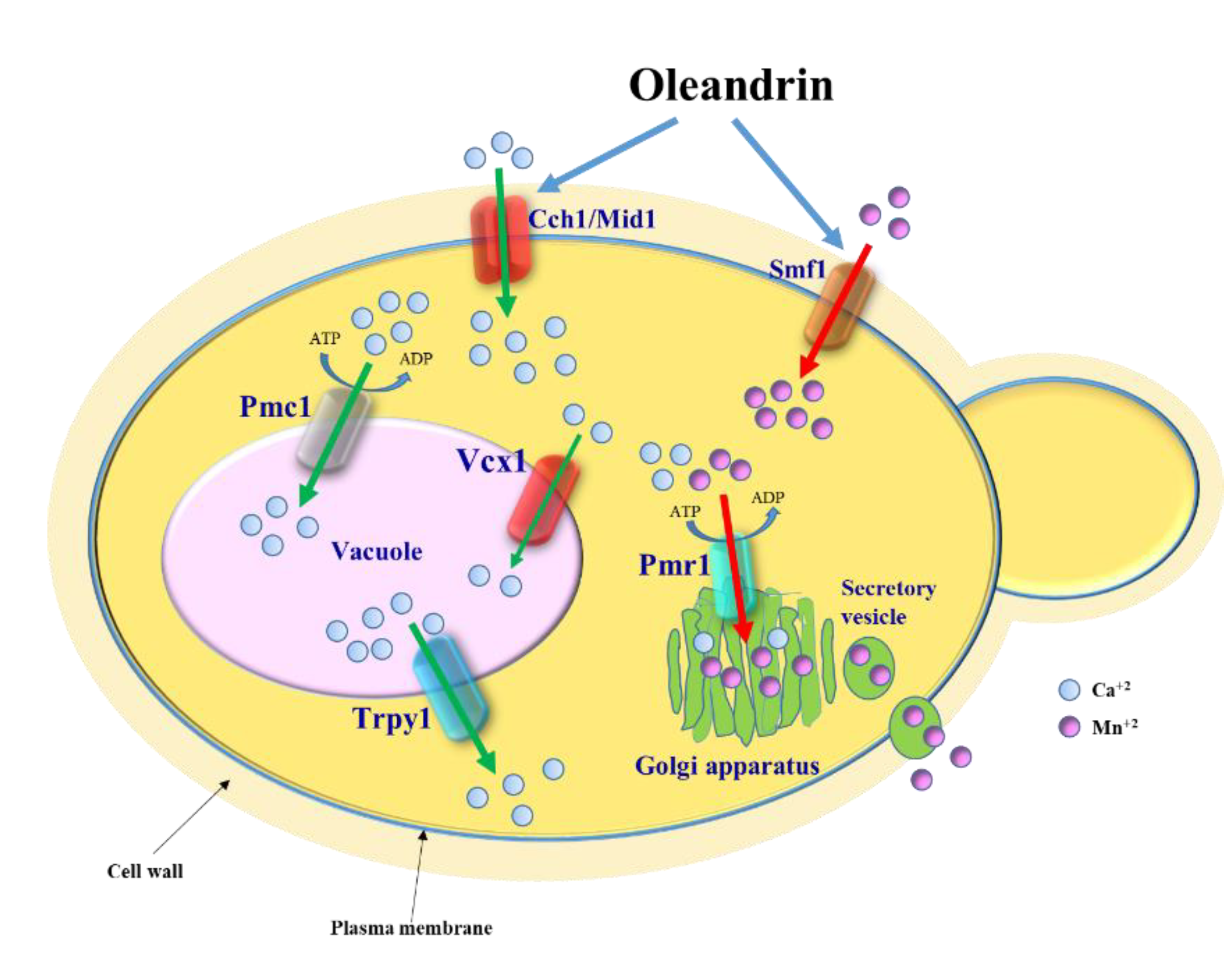
Figure 5.
Model depicting the effect of oleandrin on S. cerevisiae cells. Oleandrin stimulates Ca2+ influx via the Cch1/Mid1 channel and Mn2+ uptake via the Smf11 transporter. Excess of [Ca2+]cyt is transported into the vacuole via the Pmc1 ATPase pump and Vcx1 transporter. Cells lacking Pmc1 become oleandrin-hypersensitive due to a delay in restoring the low level of [Ca2+]cyt (Figure 3b, orange line). The excess of Mn2+ taken up via Smf1 in response to oleandrin exposure is removed by the Ca2+/Mn2+-ATPase pump Pmr1 which transports the excess Mn2+ to the Golgi apparatus to be further secreted from the cell via secretory vesicles.
Figure 5.
Model depicting the effect of oleandrin on S. cerevisiae cells. Oleandrin stimulates Ca2+ influx via the Cch1/Mid1 channel and Mn2+ uptake via the Smf11 transporter. Excess of [Ca2+]cyt is transported into the vacuole via the Pmc1 ATPase pump and Vcx1 transporter. Cells lacking Pmc1 become oleandrin-hypersensitive due to a delay in restoring the low level of [Ca2+]cyt (Figure 3b, orange line). The excess of Mn2+ taken up via Smf1 in response to oleandrin exposure is removed by the Ca2+/Mn2+-ATPase pump Pmr1 which transports the excess Mn2+ to the Golgi apparatus to be further secreted from the cell via secretory vesicles.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of oleandrin on metal content of yeast cells. Exponentially growing wild type cells were shifted to MMe (OD600 = 0.5) in the presence of oleandrin (100 ng/mL). Cells were grown with agitation for 16 h before being harvested for multi-elemental analysis, as described in the Materials and Methods section. Each determination was done in triplicate on approximately 108 cells from three biological replicates. Results are given as mean ± standard deviation.
Table 1.
Effect of oleandrin on metal content of yeast cells. Exponentially growing wild type cells were shifted to MMe (OD600 = 0.5) in the presence of oleandrin (100 ng/mL). Cells were grown with agitation for 16 h before being harvested for multi-elemental analysis, as described in the Materials and Methods section. Each determination was done in triplicate on approximately 108 cells from three biological replicates. Results are given as mean ± standard deviation.
| Metal Detected | Cellular Metal Content (nmoles/mg Total Cell Protein) | |
|---|---|---|
| No Oleandrin | Oleandrin | |
| Co2+ | 1.32 ± 0.24 | 1.44 ± 0.31 |
| Cu2+ | 5.84 ± 0.42 | 5.52 ± 0.82 |
| Fe3+ | 52.82 ± 3.24 | 54.33 ± 3.84 |
| Mn2+ | 4.25 ± 0.62 | 9.92 ± 1.82 * |
| Ni2+ | 0.24 ± 0.12 | 0.22 ± 0.21 |
| Zn2+ | 12.42 ± 1.14 | 11.88 ± 1.43 |
| Li+ | 1.14 ± 0.32 | 1.21 ± 0.22 |
* Significantly different from control (no oleandrin) as of the one-sample t test, p < 0.05.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ruta, L.L.; Popa, C.V.; Farcasanu, I.C. Cytotoxicity of Oleandrin Is Mediated by Calcium Influx and by Increased Manganese Uptake in Saccharomyces cerevisiae Cells. Molecules 2020, 25, 4259. https://doi.org/10.3390/molecules25184259
AMA Style
Ruta LL, Popa CV, Farcasanu IC. Cytotoxicity of Oleandrin Is Mediated by Calcium Influx and by Increased Manganese Uptake in Saccharomyces cerevisiae Cells. Molecules. 2020; 25(18):4259. https://doi.org/10.3390/molecules25184259
Chicago/Turabian StyleRuta, Lavinia L., Claudia V. Popa, and Ileana C. Farcasanu. 2020. "Cytotoxicity of Oleandrin Is Mediated by Calcium Influx and by Increased Manganese Uptake in Saccharomyces cerevisiae Cells" Molecules 25, no. 18: 4259. https://doi.org/10.3390/molecules25184259