
Optimal Production of Protein Hydrolysates from Monkfish By-Products: Chemical Features and Associated Biological Activities

 ,
, 

Abstract
1. Introduction
2. Results
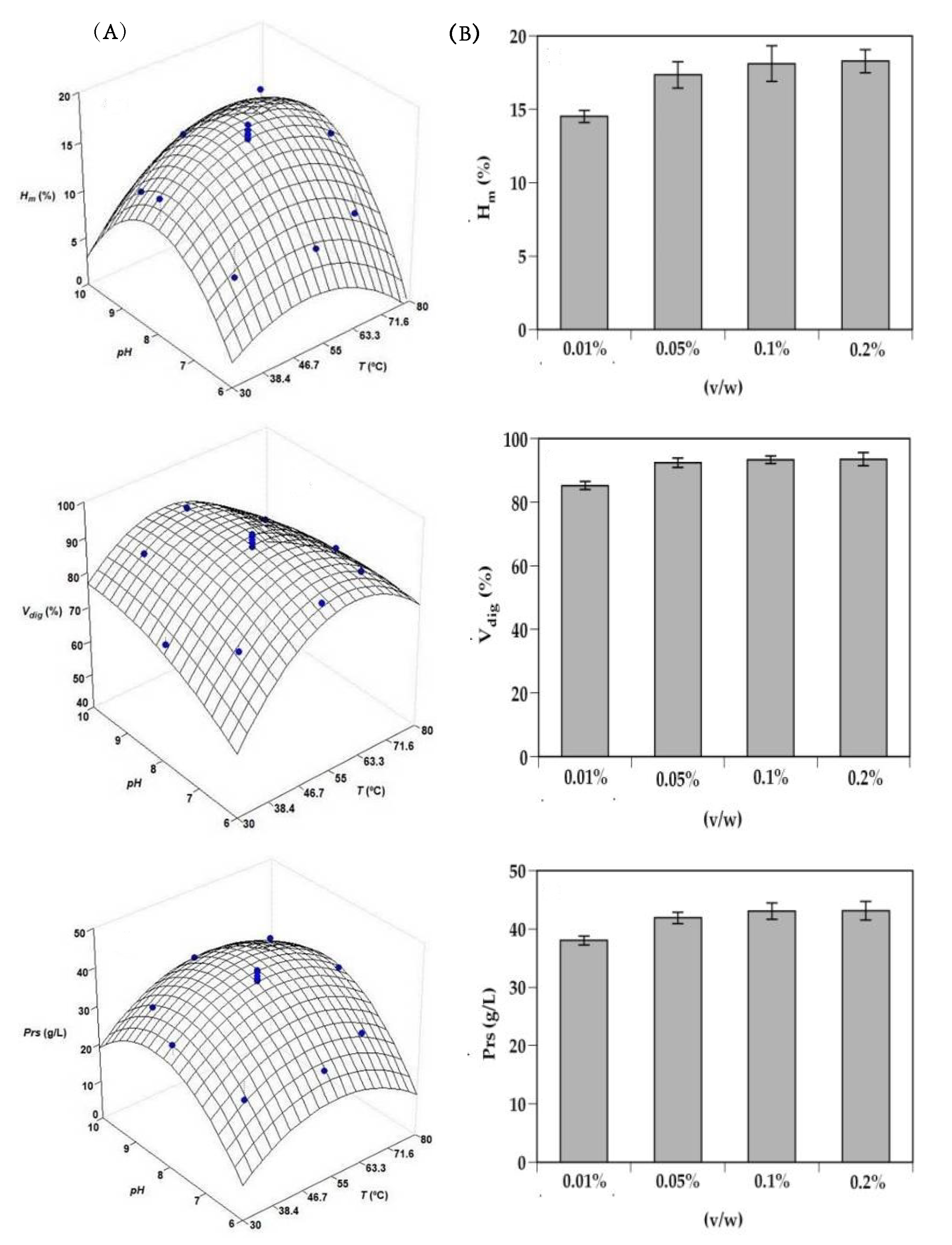
2.1. Optimization of Alcalase Hydrolysis of Head Monkfish
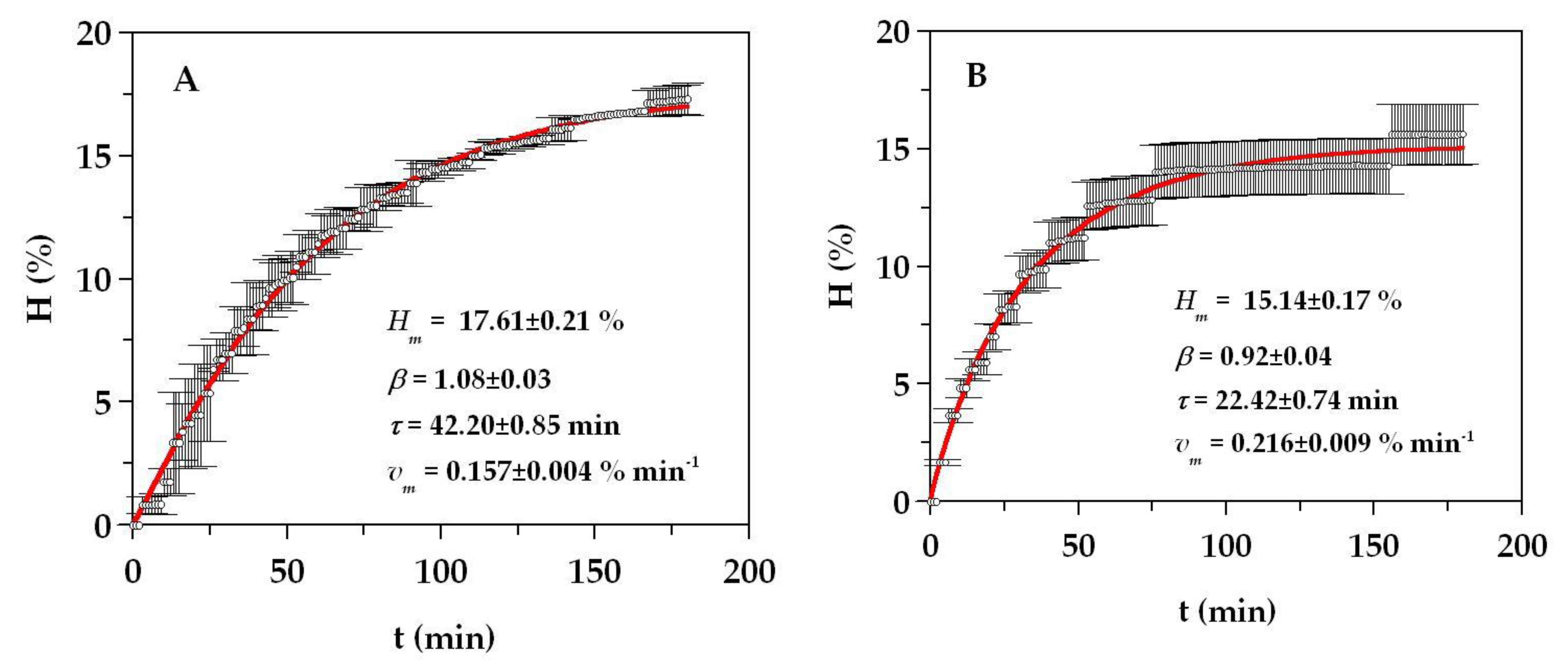
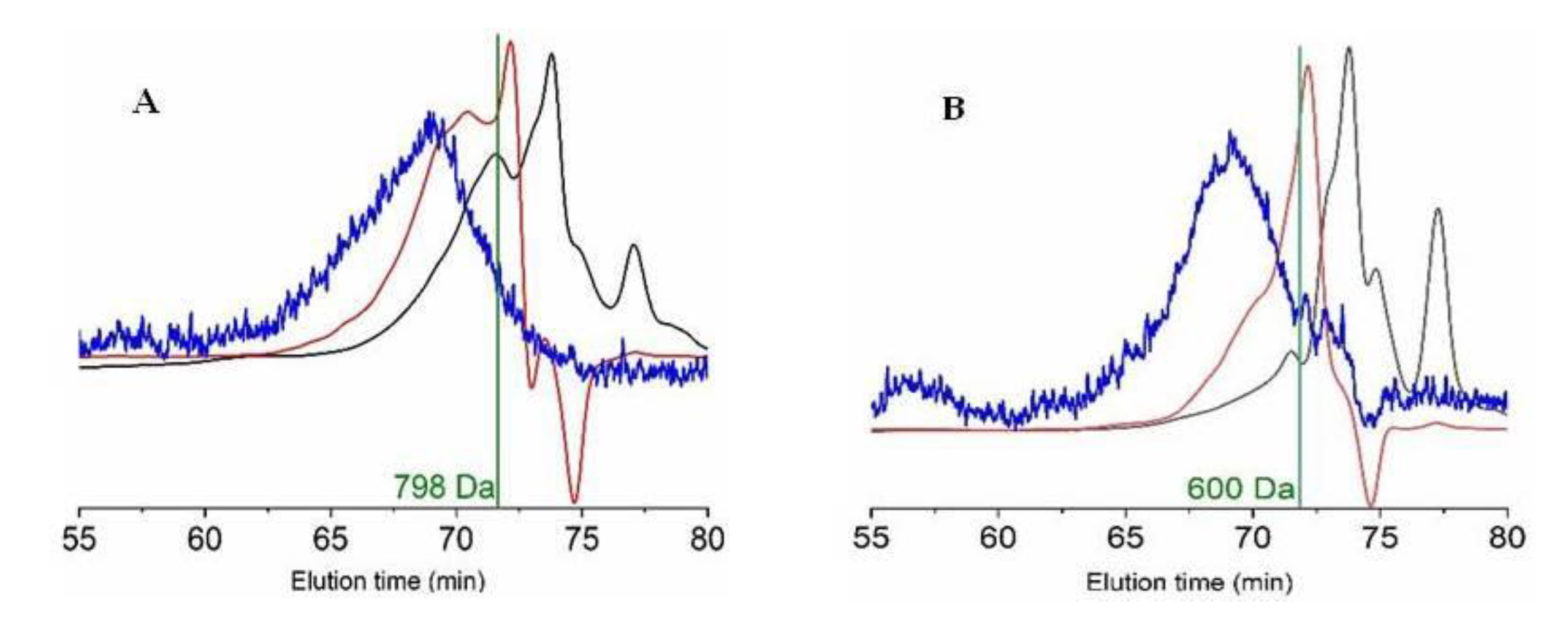
2.2. Production and Chemical Characteristics of Monkfish FPHs
2.3. Bioactivities of Monkfish FPH
3. Discussion
4. Materials and Methods
4.1. Monkfish Wastes and Proteolysis Optimization
4.2. Production of Monkfish Hydrolysates
4.3. Analytical and Biological Determinations
4.4. Numerical and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bren d’Amour, C.; Reitsma, F.; Baiocchi, G.; Barthel, S.; Güneralp, B.; Erb, K.-H.; Haberl, F.; Creutzig, F.; Seto, K.C. Future urban land expansion and implications for global croplands. Proc. Nat. Acad. Sci. USA 2017, 114, 8939–8944. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Jennings, S.; Stentiford, G.D.; Leocadio, A.M.; Jeffery, K.R.; Metcalfe, J.D.; Katsiadaki, I.; Auchterloine, N.A.; Mangi, S.C.; Pinnegar, J.K.; Ellis, T.; et al. Aquatic food security: Insights into challenges and solutions from an analysis of interactions between fisheries, aquaculture, food safety, human health, fish and human welfare, economy and environment. Fish Fish. 2016, 17, 893–938. [Google Scholar] [CrossRef]
- Arvanitoyannis, I.S.; Kassaveti, A. Fish industry waste: Treatments, environmental impacts, current and potential uses. Int. J. Food Sci. Tech. 2008, 43, 726–745. [Google Scholar] [CrossRef]
- Pérez-Martín, R.I.; Antelo, L.T.; Vázquez, J.A.; Mirón, J. An on-land management and valorisation approach for biomass associated with landing obligation compliance. Mar. Policy 2020, 116, 103506. [Google Scholar] [CrossRef]
- Blanco, M.; Vázquez, J.A.; Pérez-Martín, R.I.; Sotelo, C. Collagen extraction optimization from the skin of the small-spotted catshark (S. canicula) by response surface methodology. Mar. Drugs 2019, 17, 40. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.C.; Vázquez, J.A.; Pérez-Martín, R.I.; Carvalho, A.P.; Gomes, A.M. Valorization of by-products from commercial fish species: Extraction and chemical properties of skin gelatins. Molecules 2017, 22, 1545. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Fraguas, J.; Novoa-Carvallal, R.; Reis, R.; Antelo, L.; Pérez-Martín, R.; Valcarcel, J. Isolation and chemical characterization of chondroitin sulfate from cartilage by-products of blackmouth catshark (Galeus melastomus). Mar. Drugs 2018, 16, 344. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Ramos, P.; Valcarcel, J.; Antelo, L.T.; Novoa-Carballal, R.; Reis, R.L.; Pérez-Martín, R.I. An integral and sustainable valorisation strategy of squid pen by-products. J. Cleaner Prod. 2018, 201, 207–218. [Google Scholar] [CrossRef]
- Mackie, I.M. General review of fish protein hydrolysates. Animal Feed Sci. Tech. 1982, 7, 113–124. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Fraguas, J.; Mirón, J.; Valcárcel, J.; Pérez-Martín, R.I.; Antelo, L.T. Valorisation of fish discards assisted by enzymatic hydrolysis and microbial bioconversion: Lab and pilot plant studies and preliminary sustainability evaluation. J. Clean. Prod. 2020, 246, 119027. [Google Scholar]
- Da Rocha, M.; Alemán, A.; Baccan, G.C.; López-Caballero, M.E.; Gómez-Guillén, C.; Montero, P.; Prentice, C. Anti-Inflammatory, antioxidant, and antimicrobial effects of underutilized fish protein hydrolysate. J. Aquatic Food Prod. Tech. 2018, 27, 592–608. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.; FitzGerald, R.J. Blue whiting (Micromesistius poutassou) muscle protein hydrolysate with in vitro and in vivo antidiabetic properties. J. Funct. Foods 2018, 40, 137–145. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Rodríguez-Amado, I.; Sotelo, C.G.; Sanz, N.; Pérez-Martín, R.I.; Valcárcel, J. Production, characterization, and bioactivity of fish protein hydrolysates from aquaculture turbot (Scophthalmus maximus) wastes. Biomolecules 2020, 10, 310. [Google Scholar] [CrossRef]
- Ovissipour, M.; Abedian Kenari, A.; Motamedzadegan, A.; Nazari, R.M. Optimization of enzymatic hydrolysis of visceral waste proteins of Yellowfin Tuna (Thunnus albacares). Food Bioproc. Tech. 2012, 5, 696–705. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Sotelo, C.G.; Sanz, N.; Pérez-Martín, R.I.; Rodríguez-Amado, I.; Valcarcel, J. Valorization of aquaculture by-products of salmonids to produce enzymatic hydrolysates: Process optimization, chemical characterization and evaluation of bioactives. Mar. Drugs 2019, 17, 676. [Google Scholar] [CrossRef]
- Wu, H.-C.; Chen, H.-M.; Shiau, C.-Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Fisher, R.A.; DuPaul, B. A Fisherman’s Guide: Getting the Most Out of Monkfish; Marine Resource Advisory No. 37; Virginia Institute of Marine Science: Gloucester Point, VA, USA, 1990. [Google Scholar] [CrossRef]
- Chi, C.-F.; Wang, B.; Deng, Y.-Y.; Wang, Y.-M.; Deng, S.-G.; Ma, J.-Y. Isolation and characterization of three antioxidant pentapeptides from protein hydrolysate of monkfish (Lophius litulon) muscle. Food Res. Int. 2014, 55, 222–228. [Google Scholar] [CrossRef]
- Hu, X.-M.; Wang, Y.-M.; Zhao, Y.-Q.; Chi, C.-F.; Wang, B. Antioxidant peptides from the protein hydrolysate of monkfish (Lophius litulon) muscle: Purification, identification, and cytoprotective function on HepG2 Cells damage by H2O2. Mar. Drugs 2020, 18, 153. [Google Scholar] [CrossRef]
- Xu, J.; Li, Y.; Regenstein, J.; Su, X. In vitro and in vivo anti-oxidation and anti-fatigue effect of monkfish liver hydrolysate. Food Biosci. 2017, 18, 9–14. [Google Scholar] [CrossRef]
- Batista, I. Recovery of proteins from fish waste products by alkaline extraction. Eur. Food Res. Tech. 1999, 210, 84–89. [Google Scholar] [CrossRef]
- Greyling, N.; Bordoloi, A.; Goosen, N.J. Optimising enzymatic conditions of monkfish (Lophius vomerinus) heads hydrolysis towards potential waste biomass valorisation. Biomass Conver. Bioref. 2020, in press. [Google Scholar] [CrossRef]
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Fernández-Compás, A.; Blanco, M.; Rodríguez-Amado, I.; Moreno, H.; Borderías, J.; Pérez-Martín, R.I. Development of bioprocesses for the integral valorisation of fish discards. Biochem. Eng. J. 2019, 144, 198–208. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Menduíña, A.; Durán, A.I.; Nogueira, M.; Fernández-Compás, A.; Pérez-Martín, R.I.; Rodríguez-Amado, I. Production of valuable compounds and bioactive metabolites from by-products of fish discards using chemical processing, enzymatic hydrolysis, and bacterial fermentation. Mar. Drugs 2019, 17, 139. [Google Scholar] [CrossRef]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproducts hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- Aspmo, S.I.; Horn, S.J.; Eijsink, V.G.H. Enzymatic hydrolysis of Atlantic cod (Gadus morhua L.) viscera. Proc. Biochem. 2005, 40, 1957–1966. [Google Scholar] [CrossRef]
- Lapeña, D.; Vuoristo, K.S.; Kosa, G.; Horn, S.J.; Eijsink, V.G.H. Comparative assessment of enzymatic hydrolysis for valorization of different protein-rich industrial byproducts. J. Agric. Food Chem. 2018, 66, 9738–9749. [Google Scholar] [CrossRef]
- Safari, R.; Motamedzadegan, A.; Ovissipour, M.; Regenstein, J.M.; Gildberg, A.; Rasco, B. Use of hydrolysates from yellowfin tuna (Thunnus albacares) heads as a complex nitrogen source for lactic acid bacteria. Food Bioproc. Tech. 2012, 5, 73–79. [Google Scholar] [CrossRef]
- Sanchez, A.; Blanco, M.; Correa, B.; Perez-Martin, R.I.; Sotelo, C.G. Effect of fish collagen hydrolysates on type I collagen mRNA levels of human dermal fibroblast culture. Mar. Drugs 2018, 16, 144. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Ramos, P.; Mirón, J.; Valcarcel, J.; Sotelo, C.G.; Pérez-Martín, R.I. Production of chitin from Penaeus vannamei by-products to pilot plant scale using a combination of enzymatic and chemical processes and subsequent optimization of the chemical production of chitosan by response surface methodology. Mar. Drugs 2017, 15, 180. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Noriega, D.; Ramos, P.; Valcarcel, J.; Novoa-Carballal, R.; Pastrana, L.; Reis, R.L.; Pérez-Martín, R.I. Optimization of high purity chitin and chitosan production from Illex argentinus pens by a combination of enzymatic and chemical processes. Carbohyd. Polym. 2017, 174, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, J.C.; Goosen, N.J. Evaluation of fish protein hydrolysates in juvenile African catfish (Clarias gariepinus) diets. Aquacult. 2018, 496, 262–269. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; WHO Technical Report Series Nº935; WHO Library Cataloguing-in-Publication Data: Geneva, Switzerland; United Nations University: Tokyo, Japan, 2017; ISBN 924 1209356. [Google Scholar]
- FAO/WHO. Food and Agriculture Organization/World Health Organization Protein quality evaluation. In Proceedings of the Report of a Joint FAO/WHO Expert Consultation, Bethesda, MD, USA, 4–8 December 1989; FAO/WHO: Rome, Italy, 1990. [Google Scholar]
- Ospina-Salazar, G.H.; Ríos-Durán, M.G.; Toledo-Cuevas, E.M.; Martínez-Palacios, C.A. The effects of fish hydrolysate and soy protein isolate on the growth performance, body composition and digestibility of juvenile pike silverside, Chirostoma estor. Anim. Feed Sci. Technol. 2016, 220, 168–179. [Google Scholar] [CrossRef]
- Rajabzadeh, M.; Pourashouri, P.; Shabanpour, B.; Alishahi, A. Amino acid composition, antioxidant and functional properties of protein hydrolysates from the roe of rainbow trout (Oncorhynchus mykiss). Int. J. Food Sci. Tech. 2018, 53, 313–319. [Google Scholar] [CrossRef]
- Taheri, A.; Farvin, K.H.S.; Jacobsen, C.; Baron, C.P. Antioxidant activities and functional properties of protein and peptide fractions isolated from salted herring brine. Food Chem. 2014, 142, 318–326. [Google Scholar] [CrossRef]
- Kim, S.; Je, J.; Kim, S. Purification and characterization of antioxidant peptide from hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Cheung, L.K.Y.; Tan, N.Y.; Li-Chan, E.C.Y. The role of molecular size in antioxidant activity of peptide fractions from Pacific hake (Merluccius productus) hydrolysates. Food Chem. 2012, 134, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Aissaoui, N.; Abidi, F.; Marzouki, M.N. ACE inhibitory and antioxidant activities of red scorpionfish (Scorpaena notata) protein hydrolysates. J. Food Sci. Technol. 2015, 52, 7092–7102. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stantone, C.; Ross, R.P. Characterization of protein hydrolysates from blue whiting (Micromesistius poutassou) and their application in beverage fortification. Food Chem. 2018, 245, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.B.; Kim, J.G.; Je, J.Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H. Antioxidant and anti-freezing peptides from salmon collagen hydrolysate prepared by bacterial extracellular protease. Food Chem. 2018, 248, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Aissaoui, N.; Abidi, F.; Hardouin, J.; Abdelkafi, Z.; Marrakchi, N.; Jouenne, T.; Marzouki, M.N. Two novel peptides with angiotensin I converting enzyme inhibitory and antioxidative activities from Scorpaena notata muscle protein hydrolysate. Biotech. Appl. Biochem. 2017, 64, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Cinq-Mars, C.D.; Li-Chan, E.C. Optimizing angiotensin I-converting enzyme inhibitory activity of Pacific hake (Merluccius productus) fillet hydrolysate using response surface methodology and ultrafiltration. J. Agric. Food Chem. 2007, 55, 9380–9388. [Google Scholar] [CrossRef]
- Himaya, S.W.A.; Ngo, D.H.; Ryu, B.; Kim, S.K. An active peptide purified from gastrointestinal enzyme hydrolysate of Pacific cod skin gelatin attenuates angiotensin-1 converting enzyme (ACE) activity and cellular oxidative stress. Food Chem. 2012, 132, 1872–1882. [Google Scholar] [CrossRef]
- Je, J.Y.; Qian, Z.; Byun, H.; Kim, S. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Proc. Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Ngo, D.; Qian, Z.; Ryu, B.; Park, J.W.; Kim, S. In vitro antioxidant activity of a peptide isolated from Nile Tilapia (Oreochromis niloticus) scale gelatin in free radical-mediated oxidative systems. J. Funct. Foods 2010, 2, 107–117. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: London, UK, 1986. [Google Scholar]
- AOAC. Association of Official Analytical Chemistry. In Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1997; Volume 1–2, pp. 69–79, 864–890. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Moore, S.; Spackman, D.H.; Stein, W.H. Chromatography of amino acids on sulfonated polystyrene resins. An improved system. Anal. Chem. 1958, 30, 1185–1190. [Google Scholar] [CrossRef]
- Miller, E.L.; Bimbo, A.P.; Walters, D.E.; Barlow, S.M.; Sheridan, B. Determination of nitrogen solubility in dilute pepsin hydrochloric acid solution of fishmeal: Interlaboratory study. J. AOAC Int. 2002, 85, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Griboff, J.; Wunderlin, D.A.; Monferran, M.V. Metals, As and Se determination by inductively coupled plasma-mass spectrometry (ICP-MS) in edible fish collected from three eutrophic reservoirs. Their consumption represents a risk for human health? Microchem. J. 2017, 130, 236–244. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; González, M.P.; Murado, M.A. Production of antihypertensive and antioxidant activities by enzymatic hydrolysis of protein concentrates recovered by ultrafiltration from cuttlefish processing wastewaters. Biochem. Eng. J. 2013, 76, 43–54. [Google Scholar] [CrossRef]
- Prieto, M.A.; Curran, T.; Gowen, A.; Vázquez, J.A. An efficient methodology for quantification of synergy and antagonism in single electron transfer antioxidant assays. Food Res. Int. 2015, 67, 284–298. [Google Scholar] [CrossRef]
- Prieto, M.A.; Vázquez, J.A.; Murado, M.A. Crocin bleaching antioxidant assay revisited. Application to microplate to analyse antioxidant and prooxidant activities. Food Chem. 2015, 167, 299–310. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the monkfish hydrolysates are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WASTE | Mo (%) | OM (%) | Ash (%) | Lip (%) | Pr-tN (%) | Pr-tN (%) |
|---|---|---|---|---|---|---|
| Head | 84.6 ± 0.9 a | 11.5 ± 0.9 a | 3.9 ± 1.5 a | 3.0 ± 0.9 a | 88.3 ± 1.1 a | 89.5 ± 11.7 a |
| Viscera | 84.6 ± 0.4 a | 13.8 ± 0.4 b | 1.6 ± 0.1 b | 8.7 ± 0.4 b | 83.0 ± 4.1 b | 85.0 ± 4.0 a |
| FPH | mb (%) | Vdig (%) | Prs (g/L) | Pr-tN (g/L) | TS (g/L) | Dig (%) |
|---|---|---|---|---|---|---|
| Head | 6.8 ± 4.1 | 91.3 ± 1.5 a | 41.4 ± 1.0 a | 43.1 ± 1.1 a | 0.89 ± 0.04 a | 90.9 ± 0.5 a |
| Viscera | - | 90.0 ± 1.9 a | 38.0 ± 1.2 b | 39.5 ± 1.3 b | 1.27 ± 0.12 b | 91.8 ± 0.9 a |
| Amino Acids | Head | Viscera | Amino Acids | Head | Viscera |
|---|---|---|---|---|---|
| Asp | 9.43 ± 0.27 a | 9.97 ± 0.01 b | Leu | 6.15 ± 0.05 a | 5.55 ± 0.13 b |
| Thr | 4.03 ± 0.07 a | 3.73 ± 0.28 a | Tyr | 3.56 ± 0.29 a | 3.44 ± 0.89 a |
| Ser | 5.69 ± 0.09 a | 5.56 ± 0.08 a | Phe | 4.59 ± 0.16 a | 4.24 ± 0.10 b |
| Glu | 13.59 ± 0.52 a | 14.48 ± 1.00 a | His | 1.92 ± 0.32 a | 1.86 ± 0.11 a |
| Gly | 11.66 ± 0.60 a | 12.19 ± 0.34 a | Lys | 6.28 ± 0.10 a | 5.82 ± 0.40 a |
| Ala | 7.28 ± 0.25 a | 7.61 ± 0.00 b | Arg | 6.06 ± 0.09 a | 6.13 ± 0.63 a |
| Cys | 1.13 ± 0.28 a | 1.17 ± 0.03 a | OHPro | 3.02 ± 0.17 a | 2.87 ± 0.15 a |
| Val | 3.65 ± 0.09 a | 3.53 ± 0.00 b | Pro | 6.23 ± 0.10 a | 5.91 ± 0.57 a |
| Met | 3.20 ± 0.22 a | 3.12 ± 0.26 a | TEAA/TAA (%) | 42.06 ± 1.19 a | 40.32 ± 1.27 a |
| Ile | 2.53 ± 0.21 a | 2.80 ± 0.03 b |
| FPH | Mn (Da) | Mw (Da) | PDI | 0–0.2 kDa | 0.2–0.5 kDa | 0.5–1 kDa | 1–3 kDa | >3 kDa |
|---|---|---|---|---|---|---|---|---|
| Head | 798 ± 91 a | 1576 ± 72 a | 1.975 | 11.2 ± 4.0 a | 15.8 ± 6.7 a | 27.6 ± 3.7 a | 35.1 ± 4.4 a | 10.3 ± 2.0 a |
| Viscera | 600 ± 66 b | 947 ± 44 b | 1.578 | 35.1 ± 12.3 b | 16.5 ± 5.8 a | 22.1 ± 2.9 a | 22.8 ± 2.9 b | 3.5 ± 0.7 b |
| FPH | Mo (%) | OM (%) | Ash (%) | Lip (%) | Pr-tN (%) |
|---|---|---|---|---|---|
| Head | 9.25 ± 0.35 a | 72.28 ± 0.34 a | 18.47 ± 0.07 a | 2.39 ± 0.21 a | 69.80 ± 0.64 a |
| Viscera | 5.19 ± 0.32 b | 75.07 ± 3.91 a | 19.74 ± 3.62 a | 4.82 ± 0.15 b | 67.41 ± 0.13 b |
| Heading | ANTIOXIDANT | ANTIHYPERTENSIVE | |||
|---|---|---|---|---|---|
| FPH | DPPH (%) | ABTS (μg BHT/mL) | Crocin (μg Trolox/mL) | IACE (%) | IC50 (μg protein/mL) |
| Head | 45.05 ± 3.25 a | 13.52 ± 0.39 a | 8.52 ± 0.65 a | 63.5 ± 4.2 a | 931.3 ± 85.2 a |
| Viscera | 49.66 ± 2.65 a | 14.47 ± 0.44 b | 9.49 ± 0.28 b | 56.6 ± 2.3 b | 1142.5 ± 97.1 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, J.A.; Menduíña, A.; Nogueira, M.; Durán, A.I.; Sanz, N.; Valcarcel, J. Optimal Production of Protein Hydrolysates from Monkfish By-Products: Chemical Features and Associated Biological Activities. Molecules 2020, 25, 4068. https://doi.org/10.3390/molecules25184068
Vázquez JA, Menduíña A, Nogueira M, Durán AI, Sanz N, Valcarcel J. Optimal Production of Protein Hydrolysates from Monkfish By-Products: Chemical Features and Associated Biological Activities. Molecules. 2020; 25(18):4068. https://doi.org/10.3390/molecules25184068
Chicago/Turabian StyleVázquez, José Antonio, Araceli Menduíña, Margarita Nogueira, Ana I. Durán, Noelia Sanz, and Jesus Valcarcel. 2020. "Optimal Production of Protein Hydrolysates from Monkfish By-Products: Chemical Features and Associated Biological Activities" Molecules 25, no. 18: 4068. https://doi.org/10.3390/molecules25184068
APA StyleVázquez, J. A., Menduíña, A., Nogueira, M., Durán, A. I., Sanz, N., & Valcarcel, J. (2020). Optimal Production of Protein Hydrolysates from Monkfish By-Products: Chemical Features and Associated Biological Activities. Molecules, 25(18), 4068. https://doi.org/10.3390/molecules25184068