
Extra Virgin Olive Oil Phenols Dilate the Rat Mesenteric Artery by Activation of BKCa2+ Channels in Smooth Muscle Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
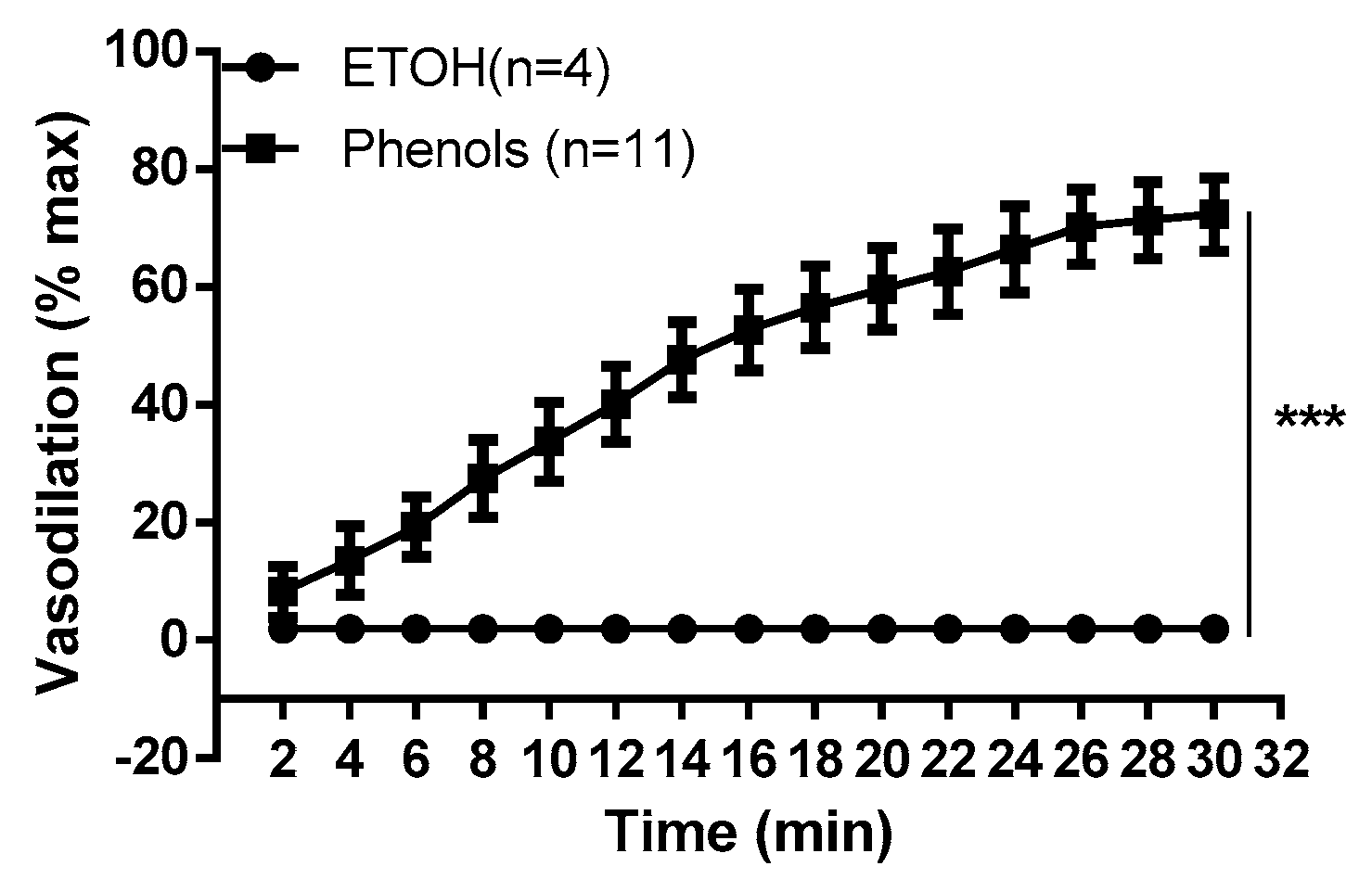
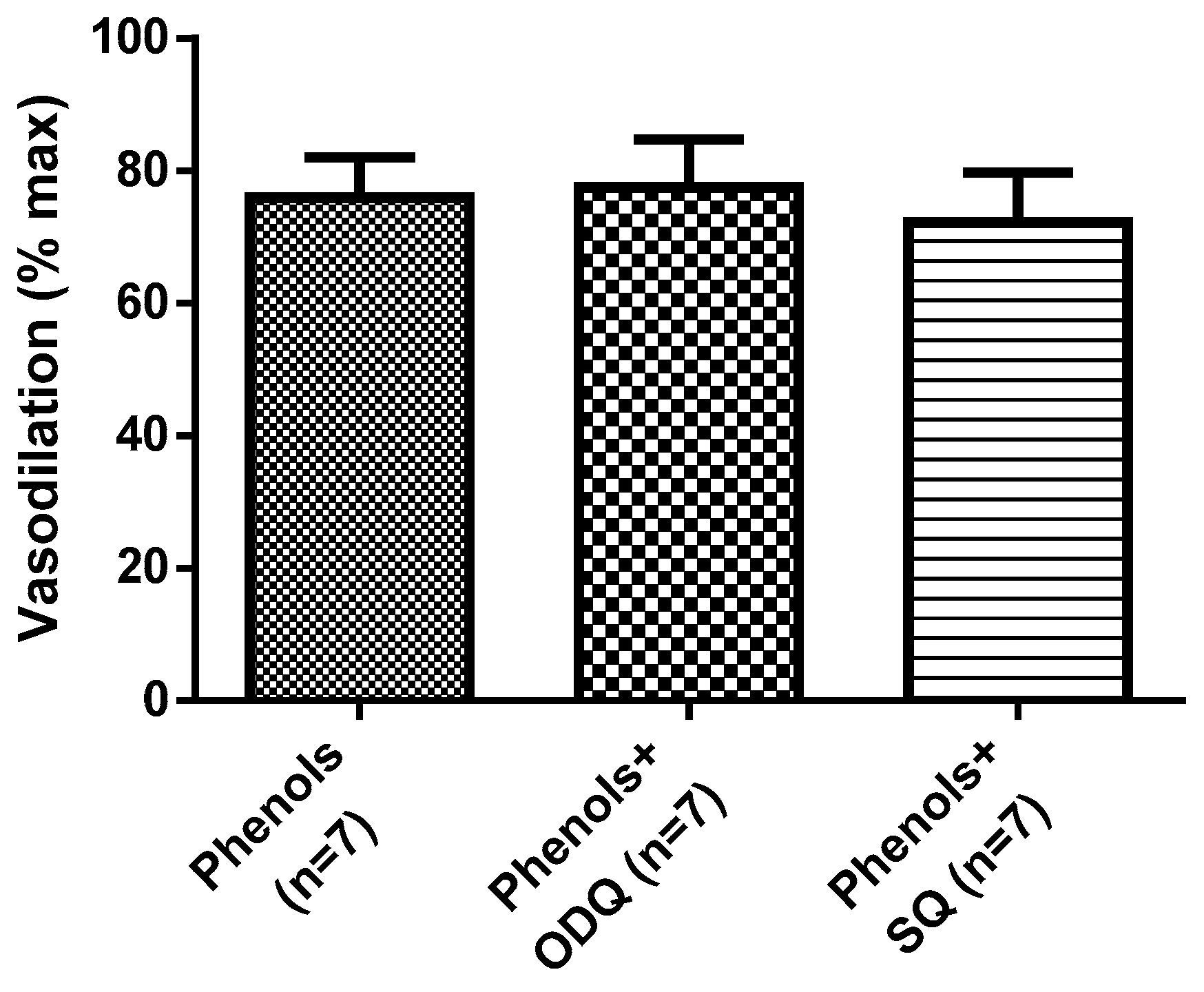
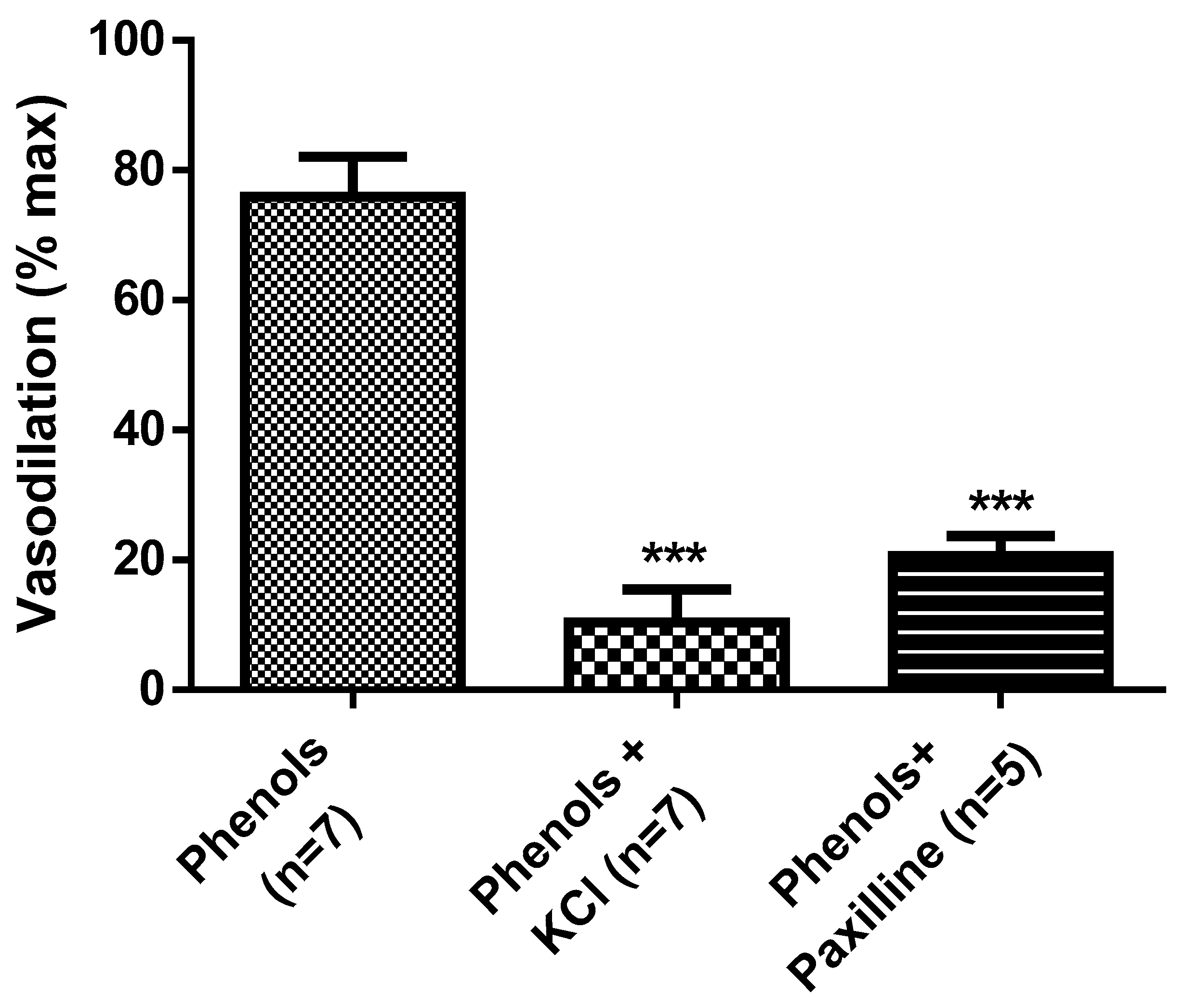
2. Results
3. Discussion
4. Materials and Methods
4.1. Extra Virgin Olive Oil Phenols
4.1.1. Extraction
4.1.2. Colorimetric Determination
4.1.3. Animals
4.1.4. Isolated Vessel Preparation
4.1.5. Reactivity Study
4.2. Drugs and Solutions
4.3. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Perona, J.S.; Canizares, J.; Montero, E.; Sanchez-Domıngueza, J.M.; Catala, A.; Ruiz-Gutierrez, V. Virgin olive oil reduces blood pressure in hypertensive elderly subjects. Clin. Nutr. 2004, 23, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Bondia-Pons, I.; Schröder, H.; Covas, M.I.; Castellote, A.I.; Kaikkonen, J.; Poulsen, H.E.; Gaddi, A.V.; Machowetz, A.; Kiesewetter, H.; López-Sabater, M.C. Moderate consumption of olive oil by healthy European men reduces systolic blood pressure in non-Mediterranean participants. J. Nutr. 2007, 137, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Safar, M.E.; Levy, B.I.; Struijker-Boudier, H. Current Perspectives on Arterial Stiffness and Pulse Pressure in Hypertension and Cardiovascular Diseases. Circulation 2003, 107, 2864–2869. [Google Scholar] [CrossRef] [PubMed]
- Drozdz, D.; Kawecka-Jaszcz, K. Cardiovascular changes during chronic hypertensive states. Pediatr. Nephrol. 2014, 29, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Psaltopoulou, T.; Naska, A.; Orfanos, P.; Trichopoulos, D.; Mountokalakis, T.; Trichopoulou, A. Olive oil, the Mediterranean diet, and arterial blood pressure: The Greek European Prospective Investigation into Cancer and Nutrition (EPIC) study. Am. J. Clin. Nutr. 2004, 80, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.A. The SUN cohort study (Seguimiento University of Navarra). Public Health Nutr. 2006, 9, 127–131. [Google Scholar] [CrossRef]
- Medina-Remón, A.; Tresserra-Rimbau, A.; Pons, A.; Tur, J.A.; Martorell, M.; Ros, E.; Buil-Cosiales, P.; Sacanella, E.; Covas, M.I.; Corella, D.; et al. Effects of total dietary polyphenols on plasma nitric oxide and blood pressure in a high cardiovascular risk cohort. The PREDIMED randomized trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 60–67. [Google Scholar] [CrossRef]
- Ruíz-Gutiérrez, V.; Muriana, F.J.; Guerrero, A.; Cert, A.M.; Villar, J. Plasma lipids, erythrocyte membrane lipids and blood pressure of hypertensive women after ingestion of dietary oleic acid from two different sources. J. Hypertens. 1996, 14, 1483–1490. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Castañer, O.; Konstantinidou, V.; Subirana, I.; Muñoz-Aguayo, D.; Blanchart, G.; Gaixas, S.; De La Torre, R.; Farré, M.; Saez, G.T.; et al. Effect of olive oil phenolic compounds on the expression of blood pressure-related genes in healthy individuals. Eur. J. Nutr. 2015, 56, 663–670. [Google Scholar] [CrossRef]
- Moreno-Luna, R.; Muñoz-Hernandez, R.; Miranda, M.L.; Costa, A.F.; Jimenez-Jimenez, L.; Vallejo-Vaz, A.J.; Muriana, F.J.; Villar, J.; Stiefel, P. Olive Oil Polyphenols Decrease Blood Pressure and Improve Endothelial Function in Young Women with Mild Hypertension. Am. J. Hypertens. 2012, 25, 1299–1304. [Google Scholar] [CrossRef]
- Vissers, M.N.; Zock, P.; Roodenburg, A.J.C.; Leenen, R.; Katan, M.B. Olive Oil Phenols Are Absorbed in Humans. J. Nutr. 2002, 132, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Edgecombe, S.C.; Stretch, G.L.; Hayball, P.J. Oleuropein, an antioxidant polyphenol from olive oil, is poorly absorbed from isolated perfused rat intestine. J. Nutr. 2000, 130, 2996–3002. [Google Scholar] [CrossRef] [PubMed]
- Tuck, K.L.; Freeman, M.P.; Hayball, P.J.; Stretch, G.L.; Stupans, I. The in vivo fate of hydroxytyrosol and tyrosol, antioxidant phenolic constituents of olive oil, after intravenous and oral dosing of labeled compounds to rats. J. Nutr. 2001, 131, 1993–1996. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, A.; Sanchez-Rodriguez, E.; Vargas, F.; Montoro-Molina, S.; Romero, M.; Calvo, J.A.E.; Vilchez, P.; Jaramillo-Carmona, S.; Olmo-García, L.; Pancorbo, A.C.; et al. Cardioprotective Effect of a Virgin Olive Oil Enriched with Bioactive Compounds in Spontaneously Hypertensive Rats. Nutrients 2019, 11, 1728. [Google Scholar] [CrossRef]
- Villarejo, A.; Ramírez-Sánchez, M.; Segarra, A.; Cañamero, M.M.M.; Prieto, I. Influence of Extra Virgin Olive Oil on Blood Pressure and Kidney Angiotensinase Activities in Spontaneously Hypertensive Rats. Planta Med. 2014, 81, 664–669. [Google Scholar] [CrossRef]
- Estruch, R.; Martínez-González, M.A.; Corella, D.; Salas-Salvadó, J.; Ruiz-Gutierrez, V.; Covas, M.I.; Fiol, M.; Gomez-Gracia, E.; López-Sabater, M.C.; Vinyoles, E.; et al. Effects of a Mediterranean-Style Diet on Cardiovascular Risk Factors. Ann. Intern. Med. 2006, 145, 1–11. [Google Scholar] [CrossRef]
- Camargo, A.; Ruano, J.; Fernandez, J.M.; Parnell, L.D.; Jimenez, A.; Santos-Gonzalez, M.; Marin, C.; Pérez-Martínez, P.; Uceda, M.; Lopez-Miranda, J.; et al. Gene expression changes in mononuclear cells in patients with metabolic syndrome after acute intake of phenol-rich virgin olive oil. BMC Genom. 2010, 11, 253. [Google Scholar] [CrossRef]
- Dell’Agli, M.; Fagnani, R.; Mitro, N.; Scurati, S.; Masciadri, M.; Mussoni, L.; Galli, G.V.; Bosisio, E.; Crestani, M.; De Fabiani, E.; et al. Minor Components of Olive Oil Modulate Proatherogenic Adhesion Molecules Involved in Endothelial Activation. J. Agric. Food Chem. 2006, 54, 3259–3264. [Google Scholar] [CrossRef]
- Covas, M.I.; Nyyssonen, K.; Poulsen, H.; Kaikkonen, J.; Zunft, H.-J.F.; Kiesewetter, H.; Gaddi, A.; De La Torre, R.; Mursu, J.; Bäumler, H.; et al. The Effect of Polyphenols in Olive Oil on Heart Disease Risk Factors. Ann. Intern. Med. 2006, 145, 333–341. [Google Scholar] [CrossRef]
- Gonzalez-Correa, J.A.; López-Villodres, J.A.; Asensi, R.; Espartero, J.L.; Rodríguez-Gutiérrez, G.; De La Cruz, J. Virgin olive oil polyphenol hydroxytyrosol acetate inhibits in vitro platelet aggregation in human whole blood: Comparison with hydroxytyrosol and acetylsalicylic acid. Br. J. Nutr. 2008, 101, 1157–1164. [Google Scholar] [CrossRef]
- Herrera, M.D.; Péerez-Guerrero, C.; Marhuenda, E.; Ruiz-Gutiéerrez, V. Effects of dietary oleic-rich oils (virgin olive and high-oleic-acid sunflower) on vascular reactivity in Wistar-Kyoto and spontaneously hypertensive rats. Br. J. Nutr. 2001, 86, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Benkhalti, F.; Legssyer, A.; Gómez, P.; Paz, E.; Lopez-Miranda, J.; Jiménez, F.P.; El Boustani, E.S. Effects of virgin olive oil phenolic compounds on LDL oxidation and vasorelaxation activity. Therapies 2003, 58, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Segade, M.; Bermejo, R.; Monteiro-Silva, F.; Paiva-Martins, F.; Gil-Longo, J.; Campos-Toimil, M. Involvement of endothelium in the vasorelaxant effects of 3,4-DHPEA-EA and 3,4-DHPEA-EDA, two major functional bioactives in olive oil. J. Funct. Foods 2016, 23, 637–646. [Google Scholar] [CrossRef]
- Wei, A.D.; Gutman, G.A.; Aldrich, R.; Chandy, K.G.; Grissmer, S.; Wulff, H. International Union of Pharmacology. LII. Nomenclature and Molecular Relationships of Calcium-Activated Potassium Channels. Pharmacol. Rev. 2005, 57, 463–472. [Google Scholar] [CrossRef]
- Gebremedhin, D.; Kaldunski, M.L.; Jacobs, E.R.; Harder, D.R.; Roman, R.J. Coexistence of two types of Ca(2+)-activated K+ channels in rat renal arterioles. Am. J. Physiol. Physiol. 1996, 270, F69–F81. [Google Scholar] [CrossRef] [PubMed]
- Jackson, W.F.; Blair, K.L. Characterization and function of Ca(2+)-activated K+ channels in arteriolar muscle cells. Am. J. Physiol. Content 1998, 274, H27–H34. [Google Scholar] [CrossRef]
- Davis, M.J.; Hill, M.A.; Kuo, L. Local regulation of microvascular perfusion. In Comprehensive Physiology; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 161–284. [Google Scholar] [CrossRef]
- Pizarro, M.L.; Becerra, M.; Sayago, A.; Beltrán, M.; Beltrán, R. Comparison of different extraction methods to determine phenolic compounds in virgin olive oil. Food Anal. Methods 2013, 6, 123–132. [Google Scholar] [CrossRef]
- Fuentes, E.; Báez, M.E.; Bravo, M.; Cid, C.; Labra, F. Determination of total phenolic content in olive oil Samples by UV–visible spectrometry and multivariate calibration. Food Anal. Methods 2012, 5, 1311–1319. [Google Scholar] [CrossRef]
- Maille, N.; Gokina, N.; Mandalà, M.; Colton, I.; Osol, G. Mechanism of hydralazine-induced relaxation in resistance arteries during pregnancy. Vasc. Pharmacol. 2016, 78, 36–42. [Google Scholar] [CrossRef]
- Colton, I.; Mandalà, M.; Morton, J.; Davidge, S.T.; Osol, G. Influence of Constriction, Wall Tension, Smooth Muscle Activation and Cellular Deformation on Rat Resistance Artery Vasodilator Reactivity. Cell. Physiol. Biochem. 2012, 29, 883–892. [Google Scholar] [CrossRef]
- Mohanty, I.; Parija, S.C.; Suklabaidya, S.; Rattan, S. Acidosis potentiates endothelium-dependent vasorelaxation and gap junction communication in the superior mesenteric artery. Eur. J. Pharmacol. 2018, 827, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Leo, C.-H.; Hart, J.L.; Woodman, O.L. 3′,4′-Dihydroxyflavonol Reduces Superoxide and Improves Nitric Oxide Function in Diabetic Rat Mesenteric Arteries. PLoS ONE 2011, 6, e20813. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Hull, A.D.; Pearce, W. Maturational modulation of endothelium-dependent vasodilatation in ovine cerebral arteries. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R149–R157. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.N.; Silva, J.F.; Silva, G.C.; Lemos, V.S.; Cortes, S. Activation of eNOS by D-pinitol Induces an Endothelium-Dependent Vasodilatation in Mouse Mesenteric Artery. Front. Pharmacol. 2018, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Legros, E.; Tirapelli, C.R.; Carrier, E.; Brochu, I.; Fournier, A.; D’Orléans-Juste, P. Characterization of the non-adrenergic/non-cholinergic response to perivascular nerve stimulation in the double-perfused mesenteric bed of the mouse. Br. J. Pharmacol. 2007, 152, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds not available. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Agostino, R.; Barberio, L.; Gatto, M.; Muzzalupo, I.; Mandalà, M. Extra Virgin Olive Oil Phenols Dilate the Rat Mesenteric Artery by Activation of BKCa2+ Channels in Smooth Muscle Cells. Molecules 2020, 25, 2601. https://doi.org/10.3390/molecules25112601
D’Agostino R, Barberio L, Gatto M, Muzzalupo I, Mandalà M. Extra Virgin Olive Oil Phenols Dilate the Rat Mesenteric Artery by Activation of BKCa2+ Channels in Smooth Muscle Cells. Molecules. 2020; 25(11):2601. https://doi.org/10.3390/molecules25112601
Chicago/Turabian StyleD’Agostino, Rossana, Laura Barberio, MariaCarmela Gatto, Innocenzo Muzzalupo, and Maurizio Mandalà. 2020. "Extra Virgin Olive Oil Phenols Dilate the Rat Mesenteric Artery by Activation of BKCa2+ Channels in Smooth Muscle Cells" Molecules 25, no. 11: 2601. https://doi.org/10.3390/molecules25112601
APA StyleD’Agostino, R., Barberio, L., Gatto, M., Muzzalupo, I., & Mandalà, M. (2020). Extra Virgin Olive Oil Phenols Dilate the Rat Mesenteric Artery by Activation of BKCa2+ Channels in Smooth Muscle Cells. Molecules, 25(11), 2601. https://doi.org/10.3390/molecules25112601