The Effect of Furanocoumarin Derivatives on Induction of Apoptosis and Multidrug Resistance in Human Leukemic Cells

 ,
,  ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Analysis of Cytotoxicity
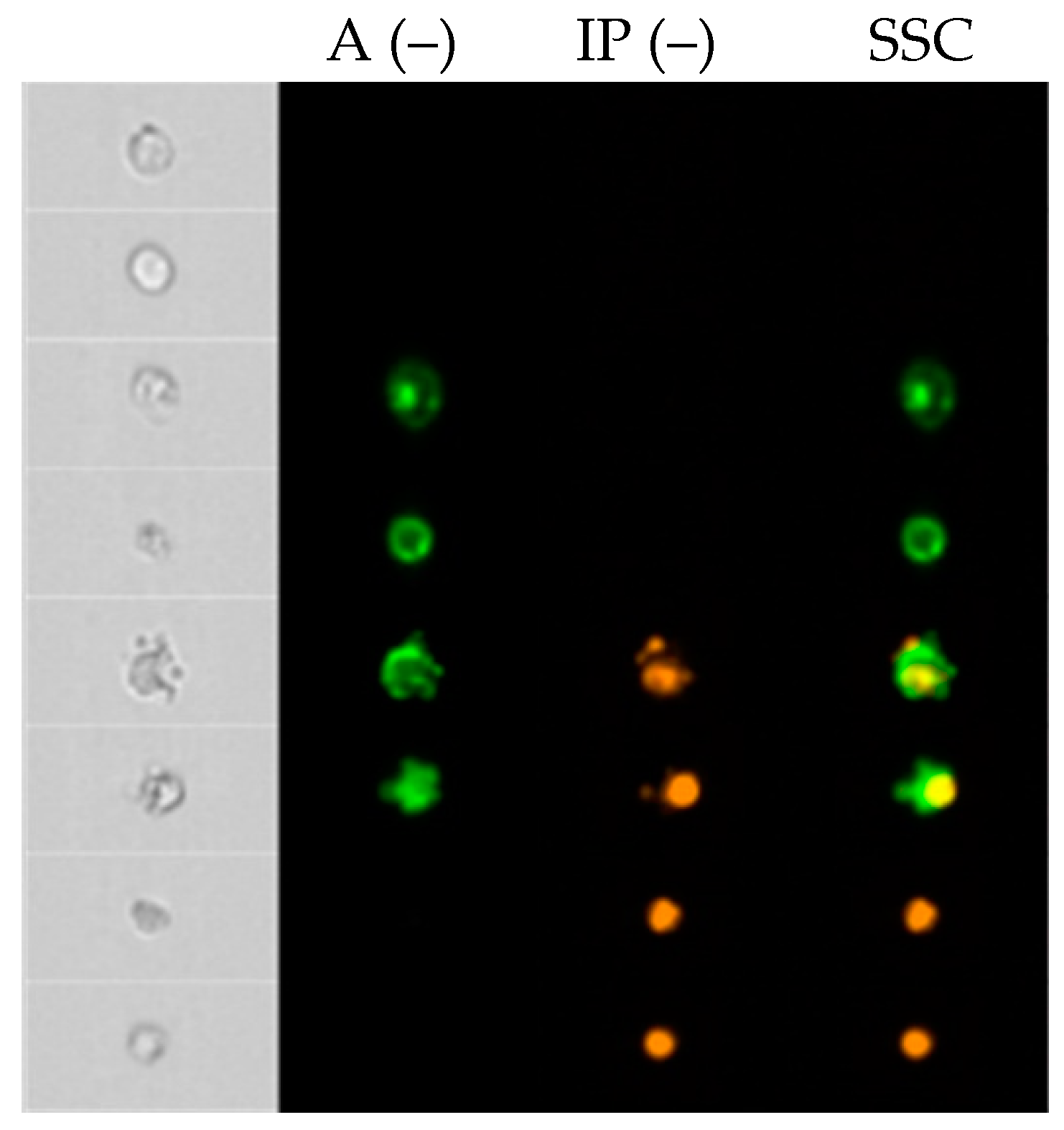
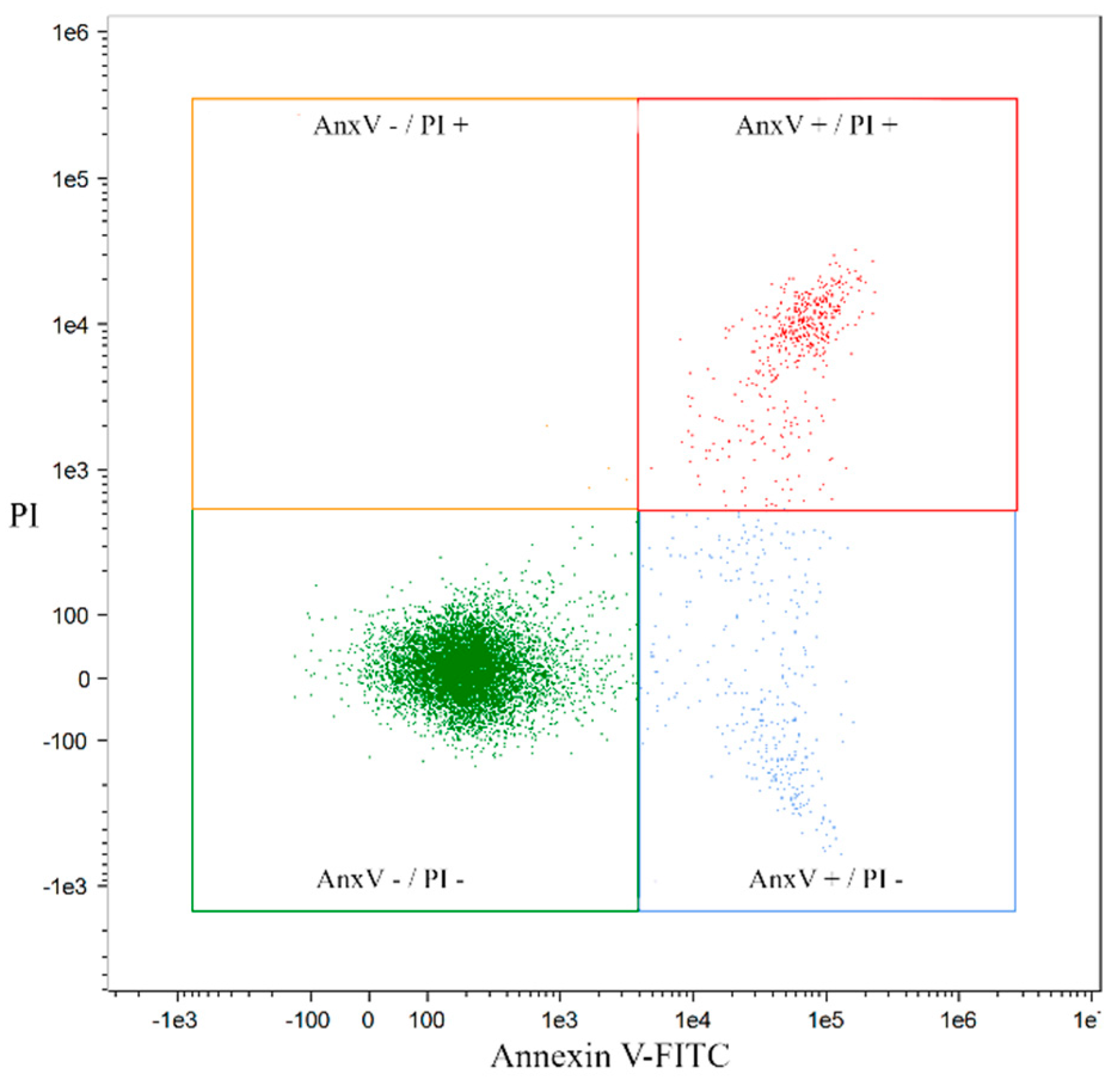
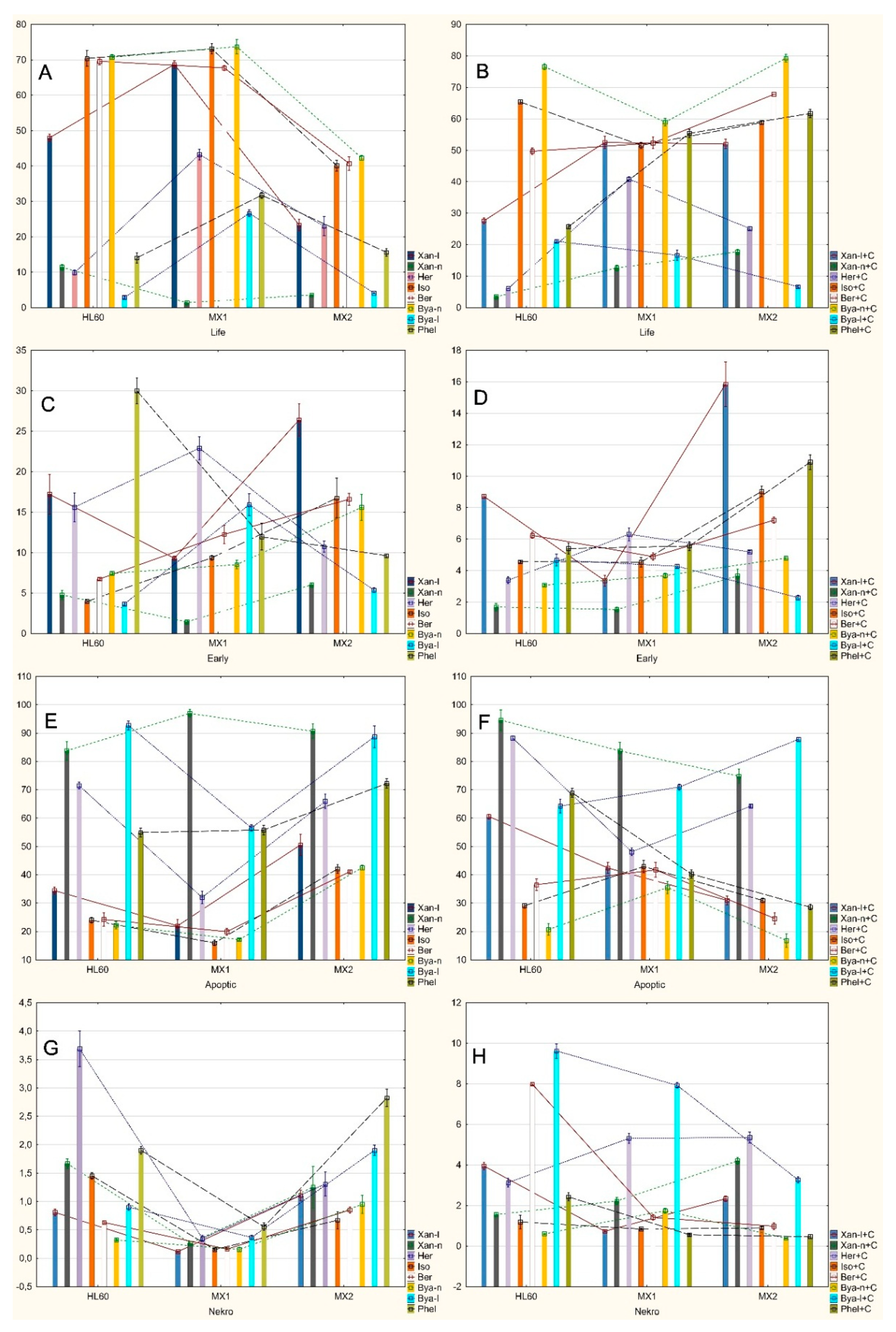
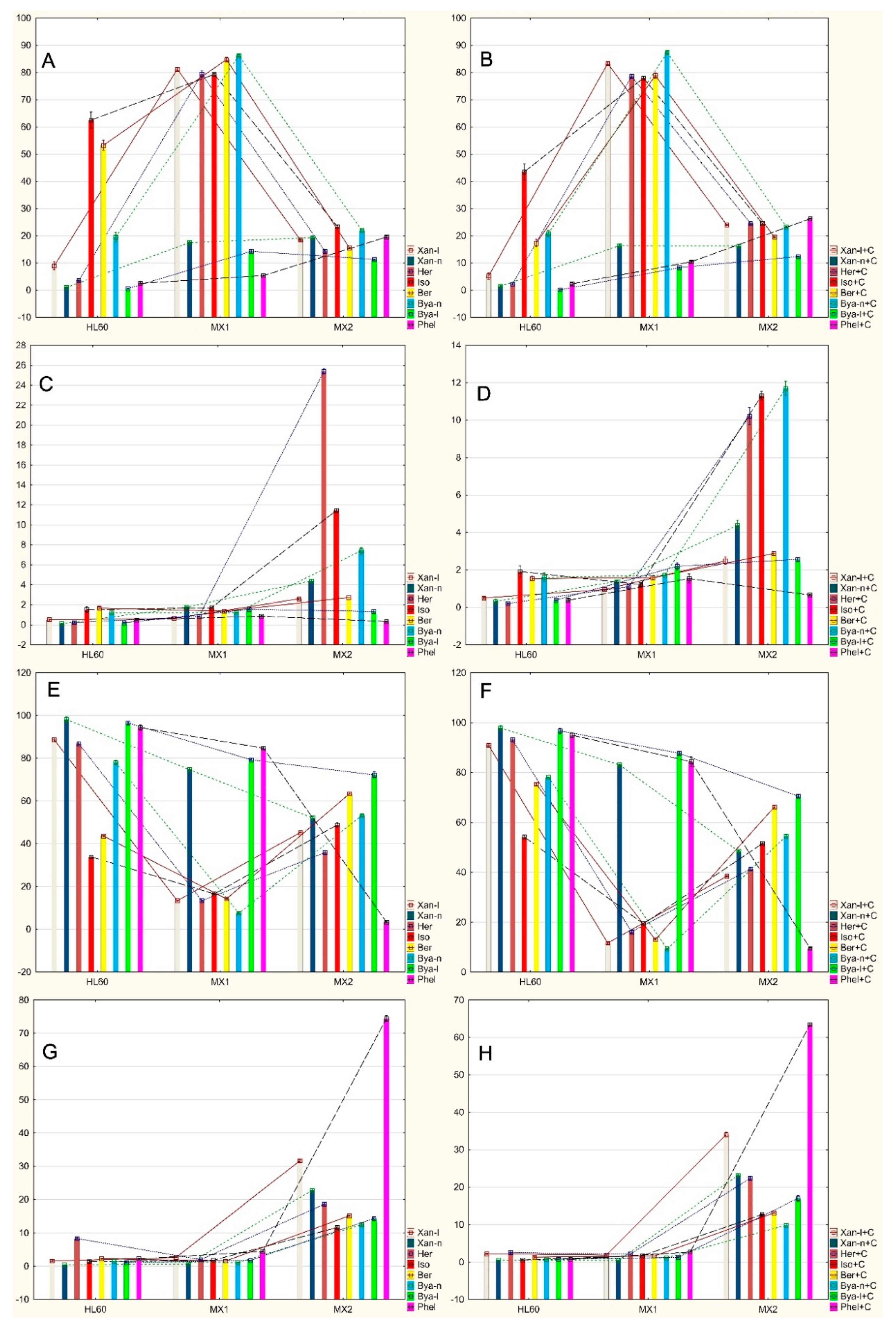
2.2. Analysis of Apoptosis Induced by Coumarin Compounds at Doses of IC50 in the Presence of Mitoxantrone (+C) and without Mitoxantrone Using AnnexinV/IP
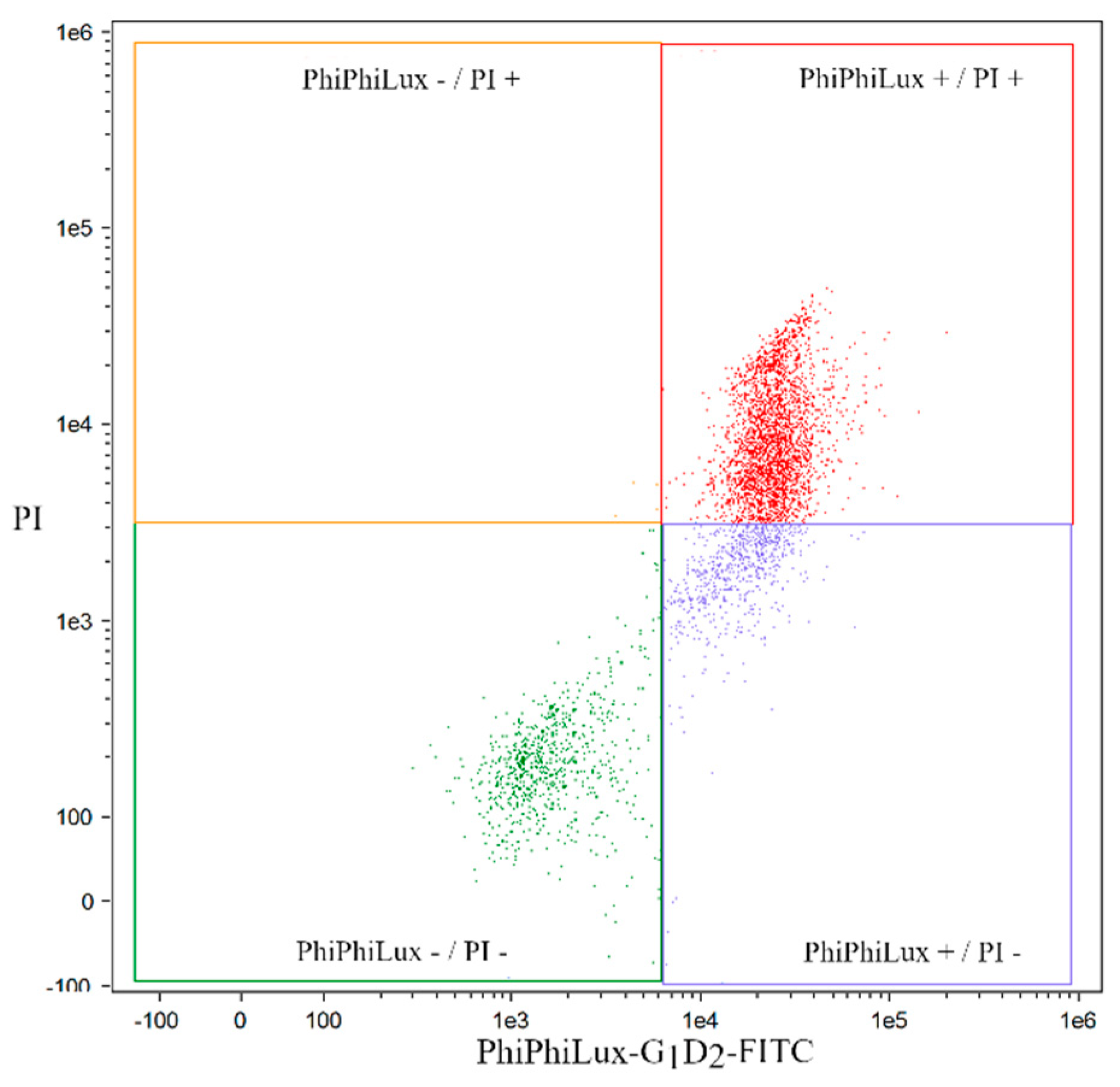
2.3. Analysis of Apoptosis Induced by Coumarin Compounds in an IC50 Dose in the Presence of Mitoxantrone (+C) and without Mitoxantrone Using Caspase 3
3. Discussion
4. Material and Methods
4.1. Cell Lines and Cell Culture
4.2. Analysis of Cell Viability
4.3. Chemicals
4.4. Cell Preparation
4.5. Quantification of Apoptosis by Annexin V and PI Double Staining
4.6. Quantification of Apoptosis by Caspase 3 (PhiPhiLux-G1D2)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jain, P.K.; Joshi, H. Coumarin: Chemical and Pharmacological Profile. J. App. Pharm. Sci. 2012, 2, 236–240. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on Natural Coumarin Lead Compounds for Their Pharmacological Activity. Biomed Res Int. 2013, 963248, 1–14. [Google Scholar] [CrossRef]
- Hassan, M.Z.; Osman, H.; Ali, M.A.; Ahsan, M.J. Therapeutic potential of coumarins as antiviral agents. Eur. J. Med. Chem. 2016, 123, 236–255. [Google Scholar] [CrossRef]
- Hu, X.L.; Xu, Z.; Liu, M.L.; Feng, L.S.; Zhang, G.D. Recent Developments of Coumarin Hybrids as Anti-fungal Agents. Curr. Top. Med. Chem. 2017, 17, 3219–3231. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Hou, Z.; Yang, X.; Mou, Y.; Guo, C. Design, Synthesis, and Mechanism of Dihydroartemisinin–Coumarin Hybrids as Potential Anti-Neuroinflammatory Agents. Molecules 2019, 24, 1672. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.D.H.; Méndez, J.; Brown, S.A. The natural Coumarins: Occurrence, Chemistry, and Biochemistry; John Willey and Sons LTD: Chichester, England; New York, USA; Brisbane, Australia; Toronto, Canada; Singapore, Asia, 1982; pp. 154–196. [Google Scholar]
- Hung, W.-L.; Suh, J.H.; Wang, Y. Chemistry and health effects of furanocoumarins in grapefruit. J. Food Drug Anal. 2017, 25, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Portt, L.; Norman, G.; Clapp, C.; Greenwood, M.; Greenwood, M.T. Anti-apoptosis and cell survival: A review. Biochim. Biophys. Acta 2011, 1813, 238–259. [Google Scholar] [CrossRef]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.-T.; Zhou, T.-T.; Liu, B.; Bao, J.-K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Reutelingsperger, C. Flow cytometry of apoptotic cell death. J. Immunol. Methods 2000, 243, 167–190. [Google Scholar] [CrossRef]
- Zhu, T.; Wang, Y.; Ding, W.; Xu, J.; Chen, R.; Xie, J.; Zhu, W.; Jia, L.; Ma, T. Anticancer activity and DNA-binding investigations of the Cu(II) and Ni(II) complexes with coumarin derivative. Chem. Biol. Drug Des. 2015, 85, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Bacso, Z.; Everson, R.B.; Eliason, J.F. The DNA of Annexin V-binding apoptotic cells is highly fragmented. Cancer Res. 2000, 60, 4623–4628. [Google Scholar]
- Darzynkiewicz, Z.; Bruno, S.; Del Bino, G.; Gorczyca, W.; Hotz, M.A.; Lassota, P.; Traganos, F. Features of apoptotic cells measured by flow cytometry. Cytometry 1992, 13, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Denecker, G.; Vercammen, D.; Declercq, W.; Vandenabeele, P. Apoptotic and necrotic cell death induced by death domain receptors. Cell Mol. Life Sci. 2001, 58, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, L.; Lazebnik, Y. Caspases disrupt the nuclear-cytoplasmic barrier. J. Cell Biol. 2000, 151, 951–959. [Google Scholar] [CrossRef]
- Kubrak, T.; Bogucka-Kocka, A.; Komsta, Ł.; Załuski, D.; Bogucki, J.; Galkowski, D.; Kaczmarczyk, R.; Feldo, M.; Cioch, M.; Kocki, J. Modulation of multidrug resistance gene expression by coumarin derivatives in human leukemic cells. Oxid. Med. Cell Longev. 2017, 5647281. [Google Scholar] [CrossRef]
- Kasaian, J.; Mosaffa, F.; Behravan, J.; Masullo, M.; Piacente, S.; Ghandadi, M.; Iranshahi, M. Reversal of P-glycoprotein-mediated multidrug resistance in MCF-7/Adr cancer cells by sesquiterpene coumarins. Fitoterapia 2015, 103, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Dua, T.K.; Bhattacharjee, N.; Das, A.; Gangopadhyay, M.; Khanra, R.; Joardar, S.; Riaz, M.; Feo, V.; Zia-Ul-Haq, M. Natural Products as Alternative Choices for P-Glycoprotein (P-gp) Inhibition. Molecules 2017, 22, 871. [Google Scholar] [CrossRef]
- Yang, L.L.; Min-Chieh, W.; Lih-Geeng, C.; Ching-Chiung, W. Cytotoxic activity of coumarins from the fruits of Cnidium monnieri on leukemia cell lines. Planta Med. 2003, 69, 1091–1095. [Google Scholar] [CrossRef]
- Kostova, I. Synthetic and natural coumarins as cytotoxic agents. Curr. Med. Chem. Anticancer Agents 2005, 5, 29–46. [Google Scholar] [CrossRef]
- Musa, M.A.; Cooperwood, J.S.; Khan, M.O. A review of coumarin derivatives in pharmacotherapy of breast cancer. Curr. Med. Chem. 2008, 15, 2664–2679. [Google Scholar] [CrossRef]
- Musa, M.A.; Badisa, V.L.; Latinwo, L.M.; Cooperwood, J.; Sinclair, A.; Abdullah, A. Cytotoxic activity of new acetoxycoumarin derivatives in cancer cell lines. Anticancer Res. 2011, 31, 2017–2022. [Google Scholar]
- Klenkar, J.; Molnar, M. Natural and synthetic coumarins as potential anticancer agents. J. Chem. Pharm. Res. 2015, 7, 1223–1238. [Google Scholar]
- Patil, J.R.; Jayaprakasha, G.K.; Kim, J.; Chidambara Murthy, K.N.; Chetti, M.B.; Nam, S.J.; Patil, B.S. 5-Geranyloxy-7-Methoxycoumarin inhibits colon cancer (SW480) cells growth by inducing apoptosis. Planta Med. 2013, 79, 219–226. [Google Scholar] [CrossRef]
- Panno, M.L.; Giordano, F.; Palma, M.G.; Bartella, V.; Rago, V.; Maggiolini, M.; Sisci, D.; Lanzino, M.; De Amicis, F.; Andò, S. Evidence that bergapten, independently of its photoactivation, enhances p53 gene expression and induces apoptosis in human breast cancer cells. Curr. Cancer Drug Targets 2009, 9, 469–481. [Google Scholar] [CrossRef]
- Panno, M.L.; Giordano, F.; Mastroianni, F.; Palma, M.G.; Bartella, V.; Carpino, A.; Aquila, S.; Andò, S. Breast cancer cell survival signal is affected by bergapten combined with an ultraviolet irradiation. FEBS Lett. 2010, 584, 2321–2326. [Google Scholar] [CrossRef]
- Santoro, M.; Guido, C.; De Amicis, F.; Sisci, D.; Cione, E.; Vincenza, D.; Donà, A.; Panno, M.L.; Aquila, S. Bergapten induces metabolic reprogramming in breast cancer cells. Oncol. Rep. 2016, 35, 568–576. [Google Scholar] [CrossRef]
- Rasul, A.; Khan, M.; Yu, B.; Ma, T.; Yang, H. Xanthoxyletin, a coumarin induces S phase arrest and apoptosis in human gastric adenocarcinoma SGC-7901 cells. Asian Pac. J. Cancer Prev. 2011, 12, 1219–1223. [Google Scholar]
- Bogucka-Kocka, A.; Smolarz, H.; Cioch, M.; Dmoszyńska, A.; Kocki, J. Xantotoxin-induced apoptosis in Chronic Myeleogenous Leukemia. Pol. J. Environ. Stud. 2005, 14, 453–454. [Google Scholar]
- Zhang, Y.-Y.; Zhang, Q.-Q.; Song, J.-L.; Zhang, L.; Jiang, C.-S.; Zhang, H. Design, synthesis, and antiproliferative evaluation of novel coumarin/2-cyanoacryloyl hybrids as apoptosis inducing agents by activation of caspase-dependent pathway. Molecules 2018, 23, 1972. [Google Scholar] [CrossRef] [PubMed]
- Teiten, M.H.; Dicato, M.; Diederich, M. Hybrid curcumin compounds: A new strategy for cancer treatment. Molecules. 2014, 19, 20839–20863. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.J.; Jiang, J.G. Pharmacological and Nutritional Effects of Natural Coumarins and Their Structure-Activity Relationships. Mol. Nutr. Food Res. 2018, 62, 1701073. [Google Scholar] [CrossRef] [PubMed]
- Kerru, N.; Singh, P.; Koorbanally, N.; Raj, R.; Kumar, V. Recent advances (2015-2016) in anticancer hybrids. Eur. J. Med. Chem. 2017, 142, 179–212. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LIFE | EARLY APOPTOTIC | LATE APOPTOTIC | NECROTIC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | ||
| Xan-l | Z | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | |
| Xan-n | Z | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 2.683282 | 1.341641 | 1.341641 | 2.385139 | 0.745356 | 1.639783 |
| P | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.021871 | 0.539137 | 0.539137 | 0.051218 | 1.000000 | 0.303151 | |
| Her | Z | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 |
| p | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | |
| Iso | Z | 1.043498 | 1.490712 | 2.534210 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 2.683282 | 1.341641 | 1.341641 |
| p | 0.890153 | 0.408111 | 0.033810 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.021871 | 0.539137 | 0.539137 | |
| Ber | Z | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | |
| Bya-n | Z | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | |
| Bya-l | Z | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.385139 | 0.745356 | 1.639783 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.051218 | 1.000000 | 0.303151 | 0.539137 | 0.539137 | 0.021871 | |
| Phel | Z | 2.385139 | 0.745356 | 1.639783 | 1.341641 | 2.683282 | 1.341641 | 0.447214 | 2.236068 | 1.788854 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.051218 | 1.000000 | 0.303151 | 0.539137 | 0.021871 | 0.539137 | 1.000000 | 0.076042 | 0.220915 | 0.539137 | 0.539137 | 0.021871 | |
| LIFE | EARLY APOPTOTIC | LATE APOPTOTIC | NECROTIC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | ||
| Xan-l+C | Z | 2.236068 | 1.788854 | 0.447214 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 |
| p | 0.076042 | 0.220915 | 1.000000 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | |
| Xan-n+C | Z | 1.341641 | 2.683282 | 1.341641 | 0.447214 | 1.788854 | 2.236068 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| P | 0.539137 | 0.021871 | 0.539137 | 1.000000 | 0.220915 | 0.076042 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Her+C | Z | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.788854 | 2.236068 | 0.447214 |
| p | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.220915 | 0.076042 | 1.000000 | |
| Iso+C | Z | 2.683282 | 1.341641 | 1.341641 | 0.149071 | 1.937926 | 2.086997 | 2.683282 | 1.341641 | 1.341641 | 1.490712 | 0.745356 | 0.745356 |
| p | 0.021871 | 0.539137 | 0.539137 | 1.000000 | 0.157897 | 0.110665 | 0.021871 | 0.539137 | 0.539137 | 0.408111 | 1.000000 | 1.000000 | |
| Ber+C | Z | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | |
| Bya-n+C | Z | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.490712 | 1.043498 | 2.534210 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.408111 | 0.890153 | 0.033810 | 0.539137 | 0.539137 | 0.021871 | |
| Bya-l+C | Z | 1.341641 | 2.683282 | 1.341641 | 0.745356 | 2.385139 | 1.639783 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.539137 | 0.021871 | 0.539137 | 1.000000 | 0.051218 | 0.303151 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Phel+C | Z | 1.341641 | 2.683282 | 1.341641 | 0.447214 | 2.236068 | 1.788854 | 1.341641 | 2.683282 | 1.341641 | 1.639783 | 2.385139 | 0.745356 |
| p | 0.539137 | 0.021871 | 0.539137 | 1.000000 | 0.076042 | 0.220915 | 0.539137 | 0.021871 | 0.539137 | 0.303151 | 0.051218 | 1.000000 | |
| LIFE | EARLY APOPTOTIC | LATE APOPTOTIC | NECROTIC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | ||
| Xan-l | Z | 2.68328 | 1.34164 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.02187 | 0.53913 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Xan-n | Z | 1.34164 | 2.68328 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| P | 0.53913 | 0.02187 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Her | Z | 2.68328 | 1.34164 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.02187 | 0.53913 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | |
| Iso | Z | 1.34164 | 1.34164 | 2.683282 | 0.447214 | 2.236068 | 1.788854 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.53913 | 0.53913 | 0.021871 | 1.000000 | 0.076042 | 0.220915 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | |
| Ber | Z | 1.34164 | 1.34164 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.53913 | 0.53913 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | |
| Bya-n | Z | 2.68328 | 1.34164 | 1.341641 | 0.447214 | 2.236068 | 1.788854 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.02187 | 0.53913 | 0.539137 | 1.000000 | 0.076042 | 0.220915 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | |
| Bya-l | Z | 2.68328 | 1.34164 | 1.341641 | 2.385139 | 1.639783 | 0.745356 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.02187 | 0.53913 | 0.539137 | 0.051218 | 0.303151 | 1.000000 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Phel | Z | 1.34164 | 2.68328 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.53913 | 0.02187 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| LIFE | EARLY APOPTOTIC | LATE APOPTOTIC | NECROTIC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | HL60/ MX1 | HL60/ MX2 | MX1/ MX2 | ||
| Xan-l+C | Z | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | |
| Xan-n+C | Z | 2.086997 | 1.937926 | 0.149071 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 |
| P | 0.110665 | 0.157897 | 1.000000 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | |
| Her+C | Z | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 1.341641 | 2.683282 |
| p | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | |
| Iso+C | Z | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Ber+C | Z | 2.683282 | 1.341641 | 1.341641 | 0.745356 | 2.385139 | 1.639783 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.021871 | 0.539137 | 0.539137 | 1.000000 | 0.051218 | 0.303151 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Bya-n+C | Z | 2.683282 | 1.341641 | 1.341641 | 0.447214 | 2.236068 | 1.788854 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.021871 | 0.539137 | 0.539137 | 1.000000 | 0.076042 | 0.220915 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Bya-l+C | Z | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
| Phel+C | Z | 1.341641 | 2.683282 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 1.341641 | 2.683282 | 1.341641 | 1.341641 | 2.683282 | 1.341641 |
| p | 0.539137 | 0.021871 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | 0.539137 | 0.021871 | 0.539137 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubrak, T.; Czop, M.; Kołodziej, P.; Ziaja-Sołtys, M.; Bogucki, J.; Makuch-Kocka, A.; Aebisher, D.; Kocki, J.; Bogucka-Kocka, A. The Effect of Furanocoumarin Derivatives on Induction of Apoptosis and Multidrug Resistance in Human Leukemic Cells. Molecules 2019, 24, 1824. https://doi.org/10.3390/molecules24091824
Kubrak T, Czop M, Kołodziej P, Ziaja-Sołtys M, Bogucki J, Makuch-Kocka A, Aebisher D, Kocki J, Bogucka-Kocka A. The Effect of Furanocoumarin Derivatives on Induction of Apoptosis and Multidrug Resistance in Human Leukemic Cells. Molecules. 2019; 24(9):1824. https://doi.org/10.3390/molecules24091824
Chicago/Turabian StyleKubrak, Tomasz, Marcin Czop, Przemysław Kołodziej, Marta Ziaja-Sołtys, Jacek Bogucki, Anna Makuch-Kocka, David Aebisher, Janusz Kocki, and Anna Bogucka-Kocka. 2019. "The Effect of Furanocoumarin Derivatives on Induction of Apoptosis and Multidrug Resistance in Human Leukemic Cells" Molecules 24, no. 9: 1824. https://doi.org/10.3390/molecules24091824
APA StyleKubrak, T., Czop, M., Kołodziej, P., Ziaja-Sołtys, M., Bogucki, J., Makuch-Kocka, A., Aebisher, D., Kocki, J., & Bogucka-Kocka, A. (2019). The Effect of Furanocoumarin Derivatives on Induction of Apoptosis and Multidrug Resistance in Human Leukemic Cells. Molecules, 24(9), 1824. https://doi.org/10.3390/molecules24091824