Comparative Study of Phenolic Profiles, Antioxidant and Antiproliferative Activities in Different Vegetative Parts of Ramie (Boehmeria nivea L.)

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
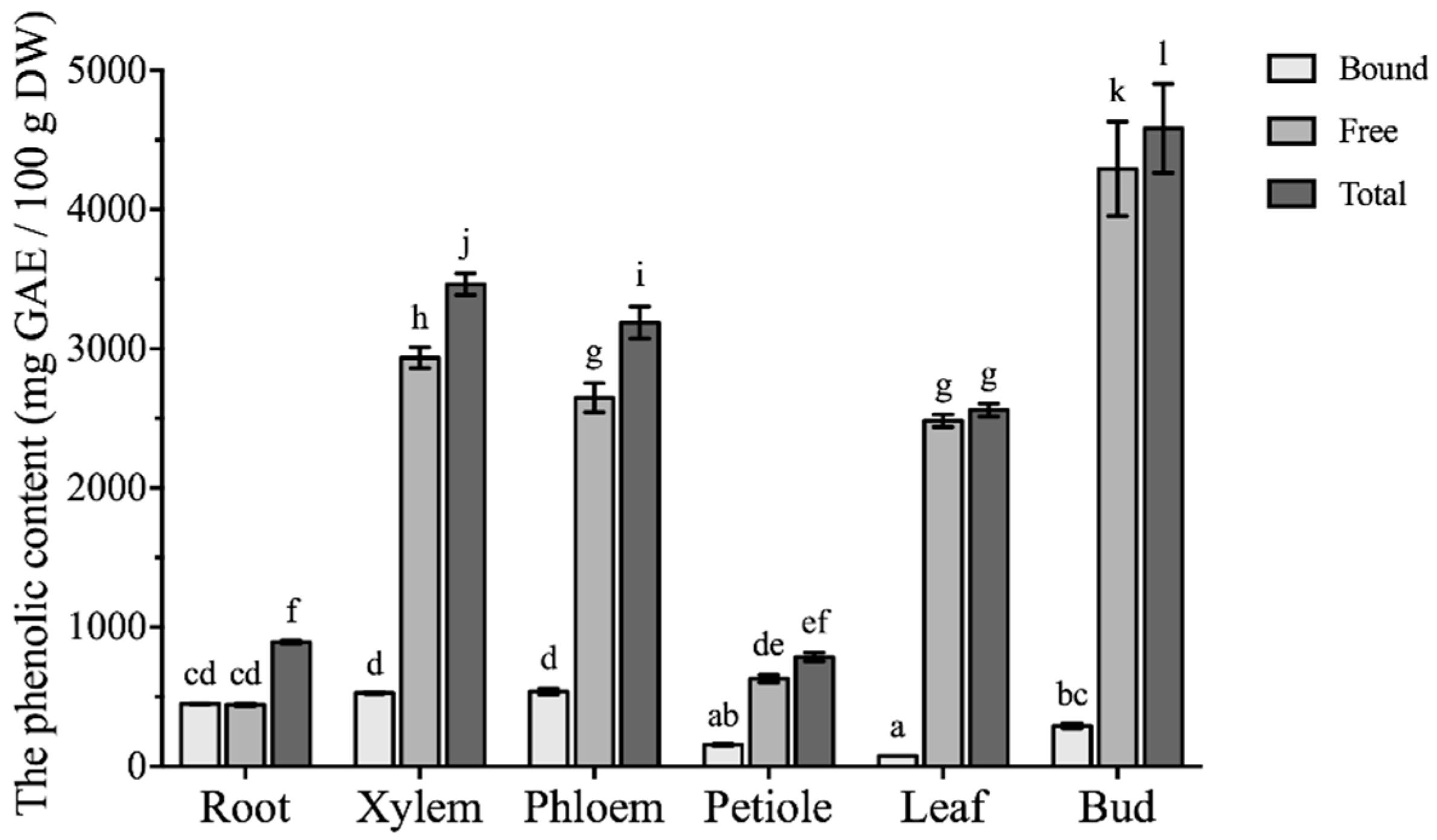
2.1. Comparison of Phenolic Content in Different Parts of Boehmeria nivea L.
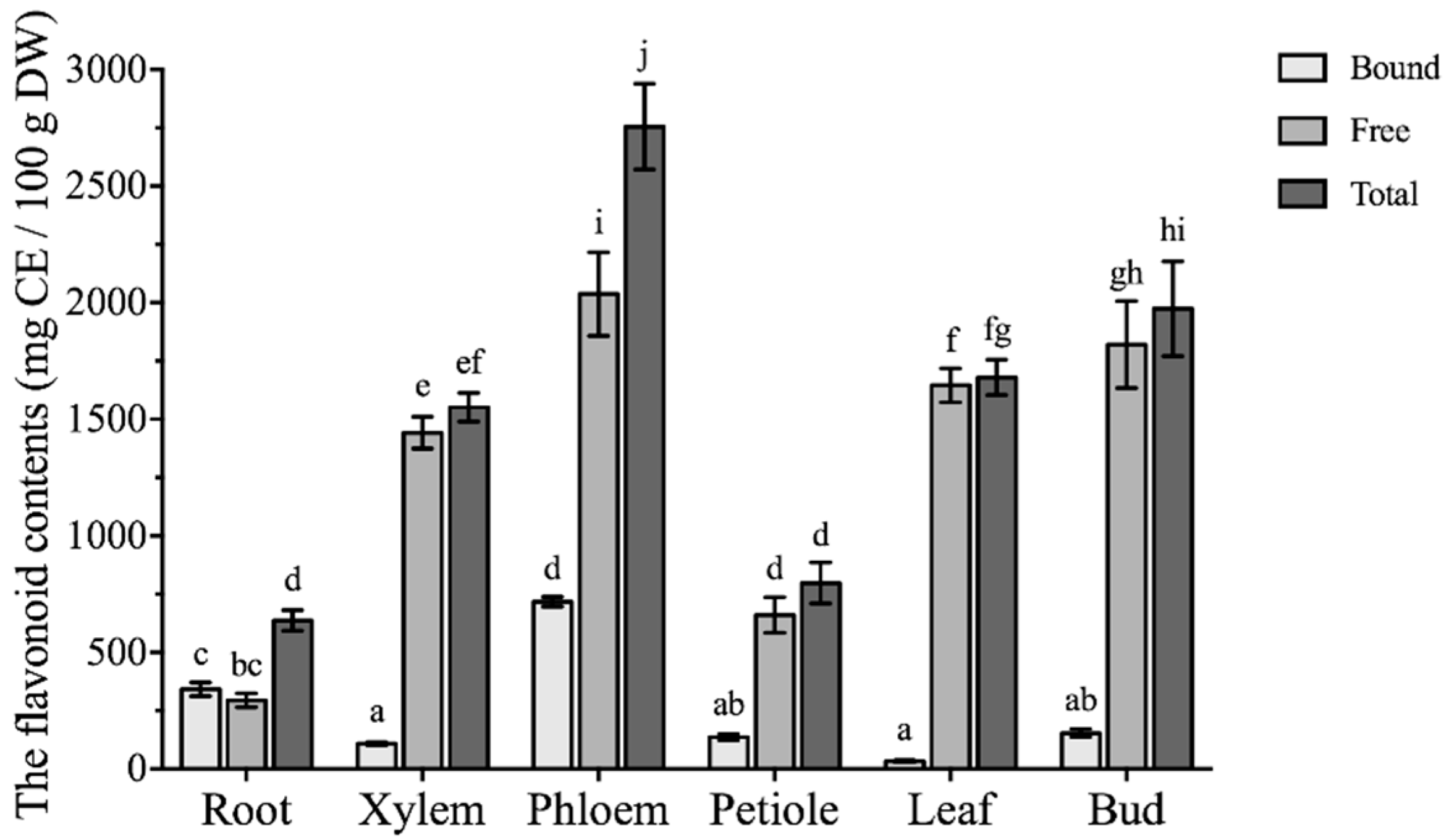
2.2. Flavonoid Contents in Different Parts of Boehmeria nivea L.
2.3. Phenolic Composition of Different Parts of Boehmeria nivea L.
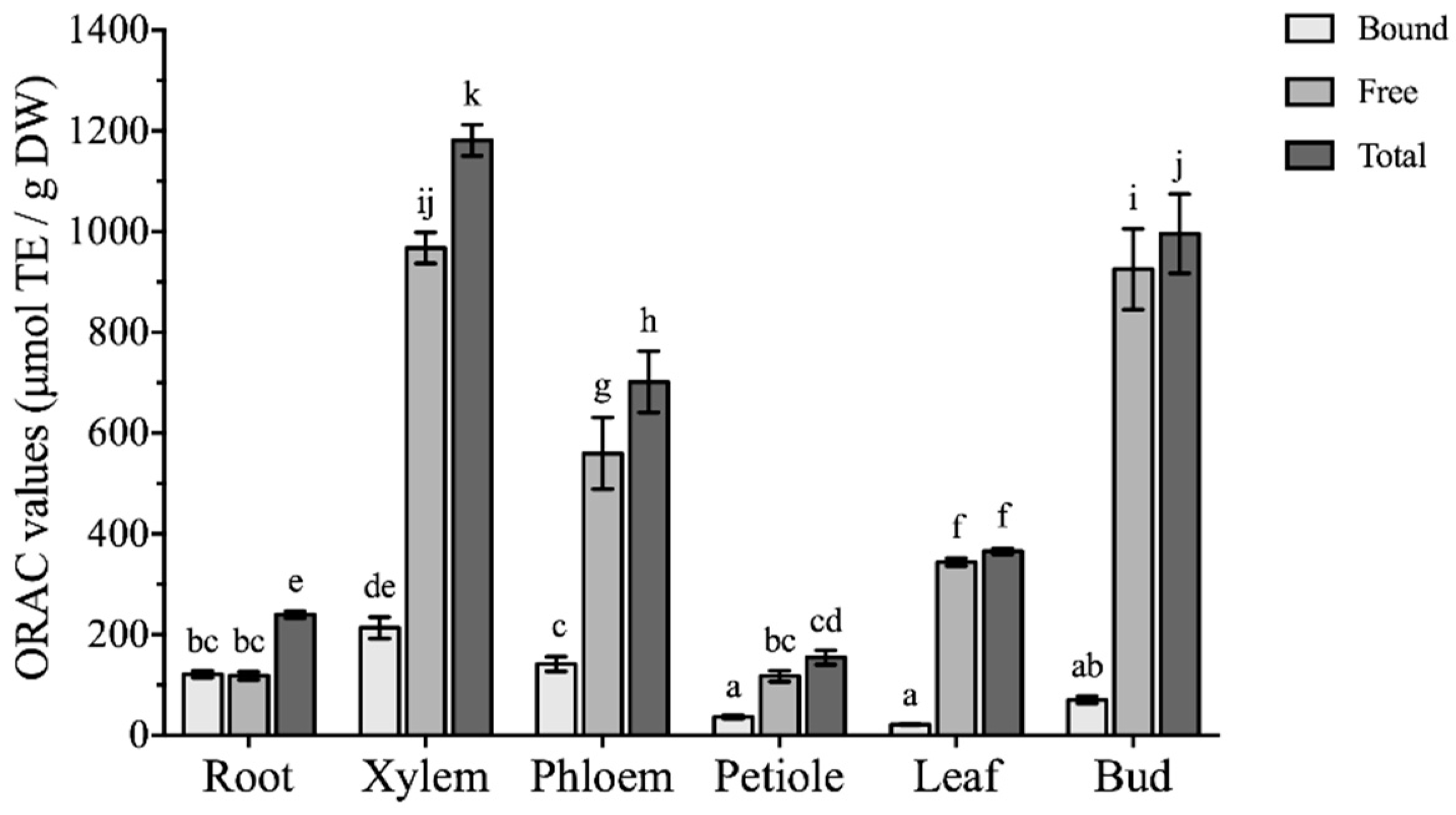
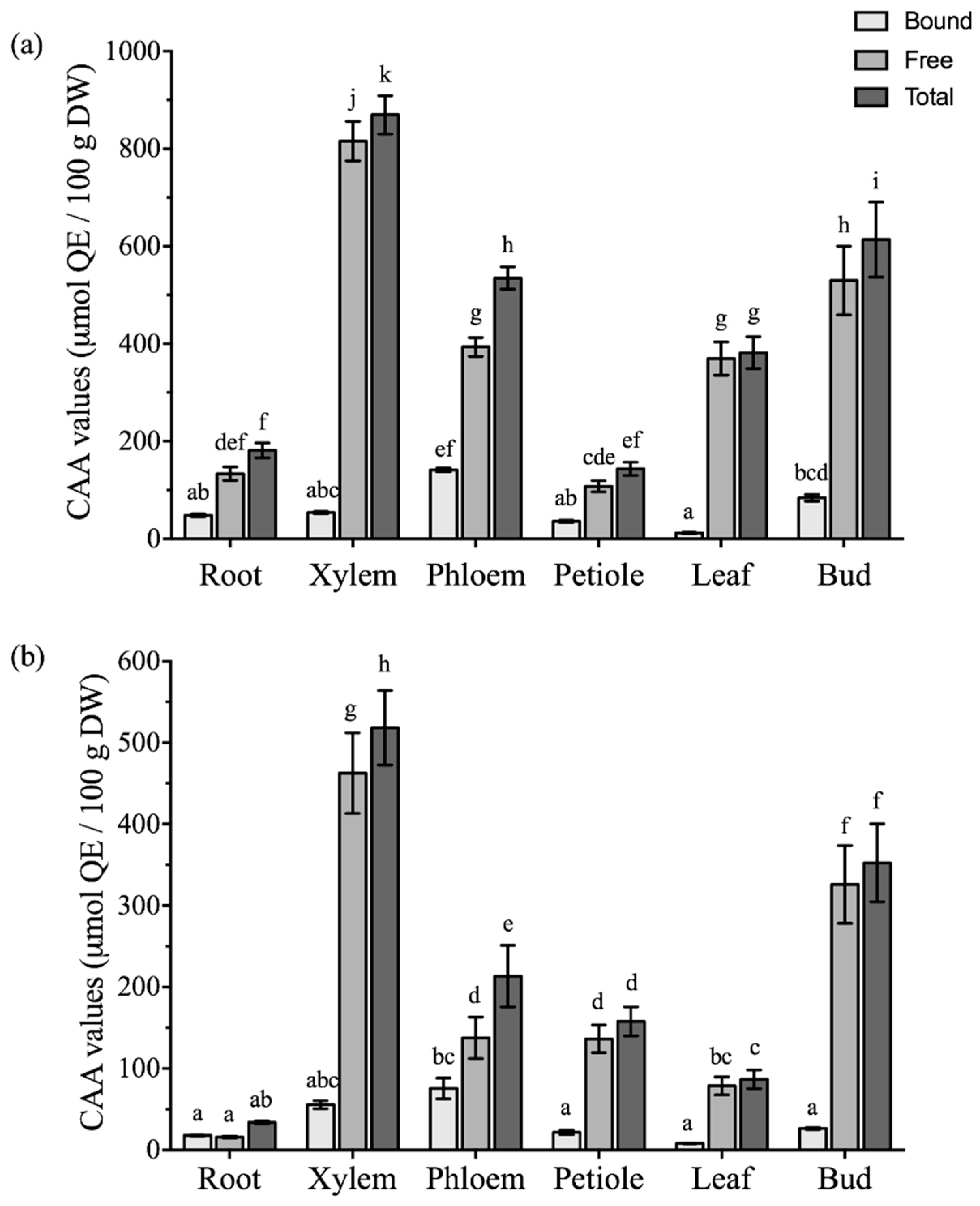
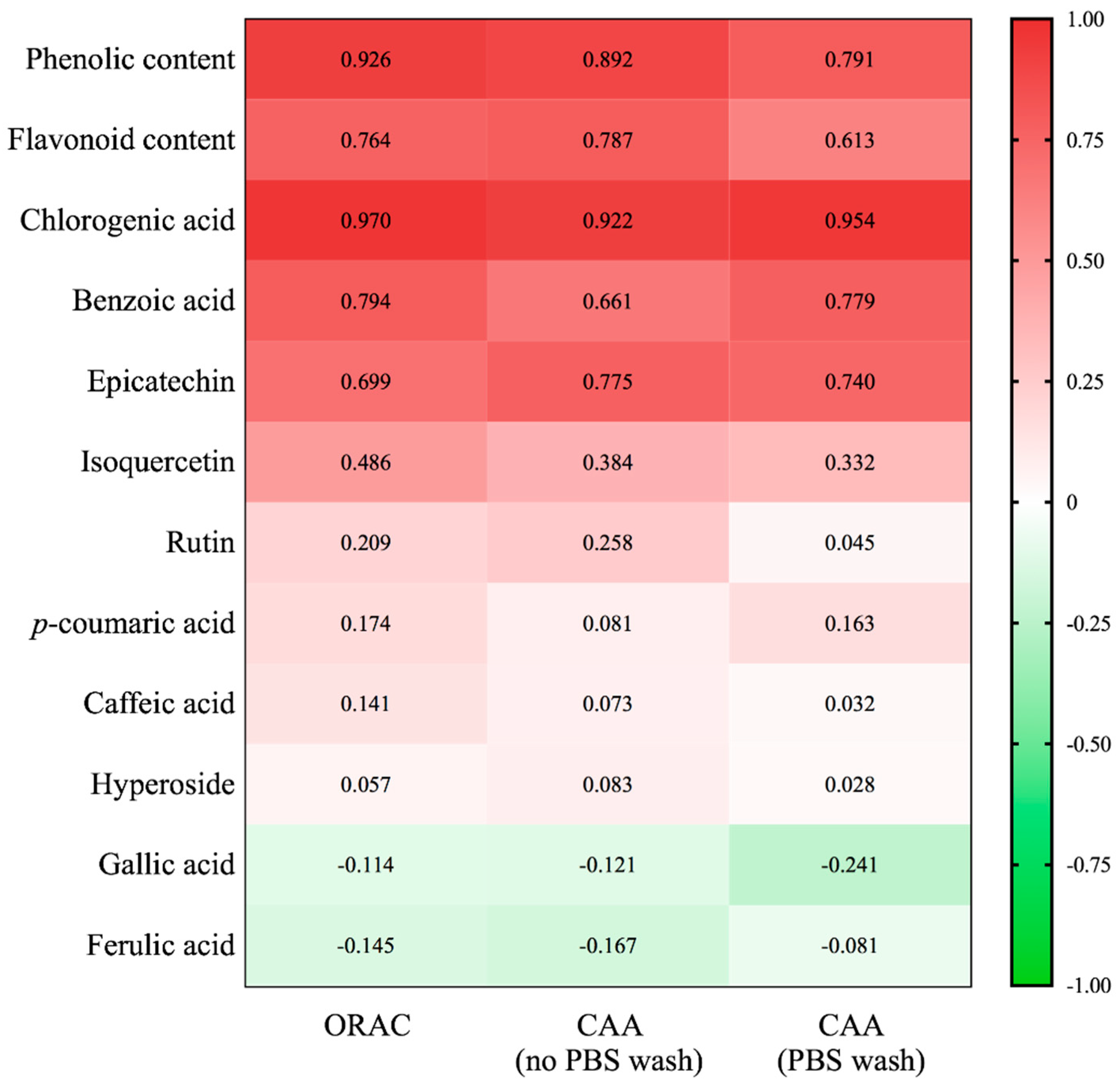
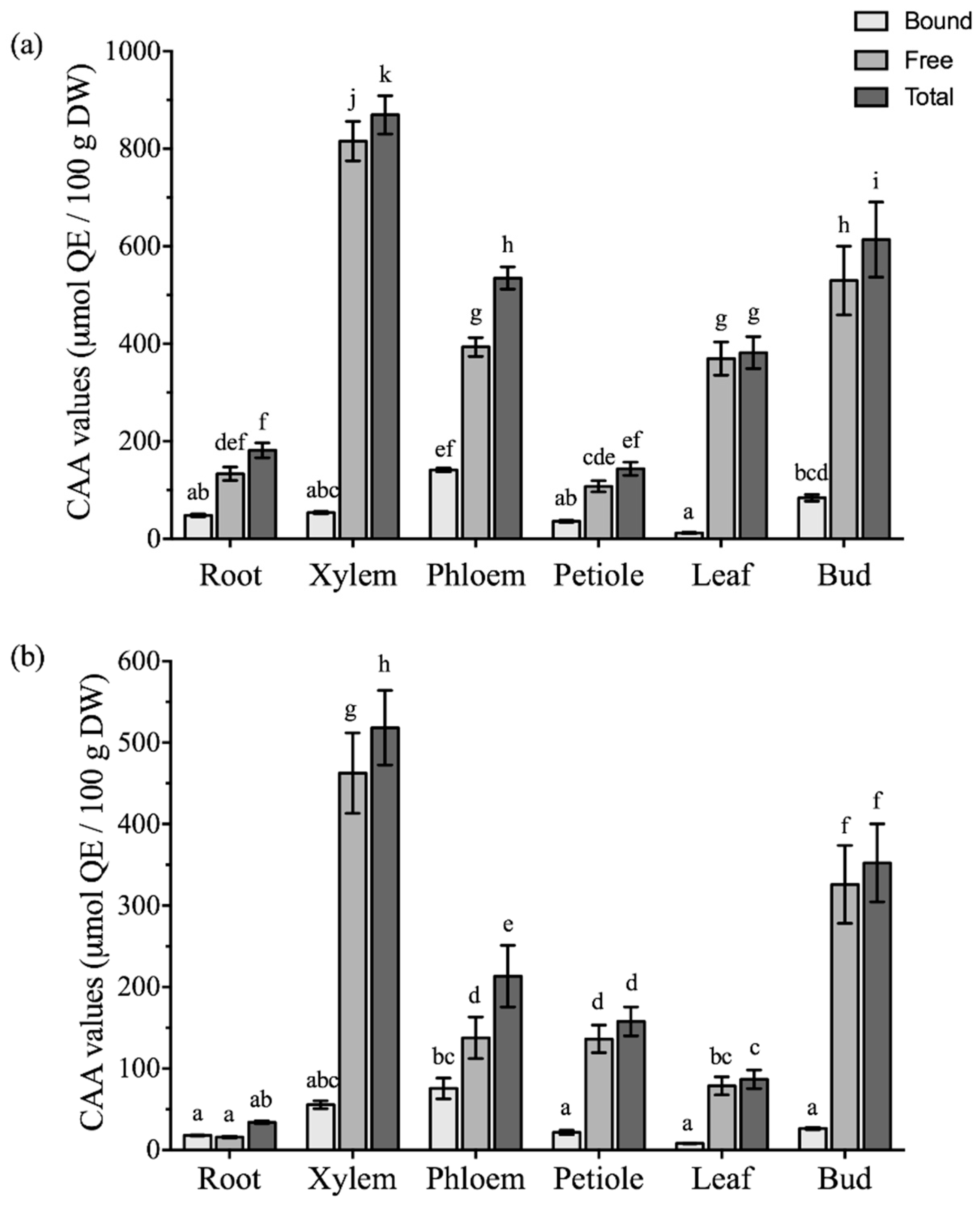
2.4. Comparison of Antioxidant Activities among Different Parts of Boehmeria nivea L.
2.5. Comparison of Antiproliferative Activities among Different Parts of Boehmeria nivea L.
3. Materials and Methods
3.1. Plant Materials
3.2. Preparation of Extracts
3.3. Determination of Total Phenolic and Flavonoid Contents
3.4. HPLC-PAD Analysis of Phenolic Compounds
3.5. Oxygen Radical Absorbance Capacity (ORAC) Assay
3.6. Cell Culture
3.7. Cellular Antioxidant Activity (CAA)
3.8. Antiproliferative Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 28 May 2018).
- Kipriotis, E.; Heping, X.; Vafeiadakis, T.; Kiprioti, M.; Alexopoulou, E. Ramie and kenaf as feed crops. Ind. Crop Prod. 2015, 68, 126–130. [Google Scholar] [CrossRef]
- Kiruthika, A.V. A review on physico-mechanical properties of bast fibre reinforced polymer composites. J. Build. Eng. 2017, 9, 91–99. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Bioactivities of phenolics by focusing on suppression of chronic diseases: A review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef]
- Giglio, R.V.; Patti, A.M.; Cicero, A.F.G.; Lippi, G.; Rizzo, M.; Toth, P.P.; Banach, M. Polyphenols: Potential use in the prevention and treatment of cardiovascular diseases. Curr. Pharm. Design 2018, 24, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Jacobo-Velazquez, D.A.; Cisneros-Zevallos, L. Recent advances in plant phenolics. Molecules 2017, 22, 1249. [Google Scholar] [CrossRef]
- Wang, Q.; Rehman, M.; Peng, D.; Liu, L. Antioxidant capacity and alpha-glucosidase inhibitory activity of leaf extracts from ten ramie cultivars. Ind. Crop Prod. 2018, 122, 430–437. [Google Scholar] [CrossRef]
- Lee, J.; Kim, A.R.; Lee, J.J. Ramie leaf extracts suppresses adipogenic differentiation in 3T3-L1 cells and pig preadipocytes. Asian Austral. J. Anim. 2016, 29, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Wang, X.; Lee, D.G.; Kim, Y.-M.; Jung, Y.-S.; Kim, H.B.; Kim, H.Y.; Cho, E.J.; Lee, S. Various biological activities of ramie (Boehmeria nivea). J. Appl. Biol. Chem. 2014, 57, 279–286. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, A.R.; Lee, J.J. Effects of ramie leaf extract on blood glucose and lipid metabolism in db/db mice. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 639–645. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, G.; Wang, H.; Cheng, C.; Zang, G.; Guo, X.; Liu, R.H. Phytochemical profiles and antioxidant activities in six species of ramie leaves. PLoS ONE 2014, 9, e108140. [Google Scholar] [CrossRef] [PubMed]
- Nho, J.W.; Hwang, I.G.; Kim, H.Y.; Lee, Y.R.; Woo, K.S.; Hwang, B.Y.; Chang, S.J.; Lee, J.; Jeong, H.S. Free radical scavenging, angiotensin I-converting enzyme (ACE) inhibitory, and in vitro anticancer activities of ramie (Boehmeria nivea) leaves extracts. Food Sci. Biotechnol. 2010, 19, 383–390. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; Chinese Medical Science and Technology Press: Beijing, China, 2010; pp. 176–177. [Google Scholar]
- Cheynier, V. Phenolic compounds: From plants to foods. Phytochem. Rev. 2012, 11, 153–177. [Google Scholar] [CrossRef]
- Liu, R.H.; Felice, D.L. Antioxidants and whole food phytochemicals for cancer prevention. In Antioxidant Measurement and Applications; Shahidi, F., Ho, C.T., Eds.; American Chemical Society: Washington, DC, USA, 2007; Volume 956, pp. 15–34. [Google Scholar]
- Liu, R.H. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78, A18–A25. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Estrada, B.A.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Saura-Calixto, F. Concept and health-related properties of nonextractable polyphenols: The missing dietary polyphenols. J. Agric. Food Chem. 2012, 60, 11195–11200. [Google Scholar] [CrossRef] [PubMed]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef]
- Lee, D.G.; Cho, S.; Lee, J.; Yang, S.; Jung, Y.-S.; Kim, H.B.; Cho, E.J.; Sanghyun, L. Quantitative analysis of the flavonoid content in the leaves of Boehmeria nivea and related commercial products. Nat. Prod. Sci. 2015, 21, 66–70. [Google Scholar]
- Santana-Galvez, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef]
- Upadhyay, R.; Rao, L.J.M. An outlook on chlorogenic acids-occurrence, chemistry, technology, and biological activities. Crit. Rev. Food Sci. 2013, 53, 968–984. [Google Scholar] [CrossRef]
- Boz, H. p-Coumaric acid in cereals: Presence, antioxidant and antimicrobial effects. Int. J. Food Sci. Technol. 2015, 50, 2323–2328. [Google Scholar] [CrossRef]
- Pei, K.H.; Ou, J.Y.; Huang, J.Q.; Ou, S.Y. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.J.; Ou, B.X.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate flourescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Fraga, C.G.; Oteiza, P.I.; Galleano, M. Plant bioactives and redox signaling: (-)-Epicatechin as a paradigm. Mol. Aspects Med. 2018, 61, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Apple peels as a value-added food ingredient. J. Agric. Food Chem. 2003, 51, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Wu, X.Z.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, G.; Fu, X.; Liu, R.H. Phytochemical profiles and antioxidant activity of different varieties of Adinandra tea (Adinandra Jack). J. Agric. Food Chem. 2015, 63, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Oxidants and Antioxidants, Pt A; Packer, L., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Gong, E.S.; Luo, S.J.; Li, T.; Liu, C.M.; Zhang, G.W.; Chen, J.; Zeng, Z.C.; Liu, R.H. Phytochemical profiles and antioxidant activity of brown rice varieties. Food Chem. 2017, 227, 432–443. [Google Scholar] [CrossRef] [PubMed]
- He, X.J.; Liu, D.; Liu, R.H. Sodium borohydride/chloranil-based assay for quantifying total flavonoids. J. Agric. Food Chem. 2008, 56, 9337–9344. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qiu, C.; Abbasi, A.M.; Chen, G.; You, L.; Li, T.; Fu, X.; Wang, Y.; Guo, X.; Liu, R.H. Effect of germination on vitamin C, phenolic compounds and antioxidant activity in flaxseed (Linum usitatissimum L.). Int. J. Food Sci. Tech. 2015, 50, 2545–2553. [Google Scholar] [CrossRef]
- Zhang, M.W.; Zhang, R.F.; Zhang, F.X.; Liu, R.H. Phenolic profiles and antioxidant activity of black rice bran of different commercially available varieties. J. Agric. Food Chem. 2010, 58, 7580–7587. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.H.; Guo, X.B.; Brennan, C.S.; Li, T.; Fu, X.; Chen, G.; Liu, R.H. Effect of germination on lignan biosynthesis, and antioxidant and antiproliferative activities in flaxseed (Linum usitatissimum L.). Food Chem. 2016, 205, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Liu, R.H. Effect of 2 alpha-hydroxyursolic acid on NF-kappa B activation induced by TNF-alpha in human breast cancer MCF-7 cells. J. Agric. Food Chem. 2008, 56, 8412–8417. [Google Scholar] [CrossRef] [PubMed]
- Felice, D.L.; Sun, J.; Liu, R.H. A modified methylene blue assay for accurate cell counting. J. Funct. Foods 2009, 1, 109–118. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetative Parts | Phenolic Acids | Flavonoids | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chlorogenic Acid | Caffeic Acid | P-Coumaric Acid | Ferulic Acid | Gallic Acid | Benzoic Acid | Epicatechin | Rutin | Isoquercetin | Hyperoside | ||
| Root | Bound | 14.10 ± 2.16 a * | 102.7 ± 1.8 h | 2087 ± 58 j | 31.30 ± 2.47 c | 10.15 ± 2.53 c | nd # | nd | 12.93 ± 2.35 bc | 31.61 ± 2.86 b | nd |
| Free | 43.48 ± 3.03 bc | nd | 60.86 ± 4.37 ab | 5.50 ± 0.18 a | 12.50 ± 1.38 c | 7.34 ± 1.16 a | nd | 35.07 ± 2.81 d | nd | 48.32 ± 2.75 d | |
| Total † | 57.58 ± 1.78 c | 102.7 ± 1.8 h | 2148 ± 56 k | 36.80 ± 2.35 d | 22.64 ± 3.79 d | 7.34 ± 1.16 a | nd | 48.00 ± 5.16 e | 31.61 ± 2.86 b | 48.32 ± 2.75 d | |
| Xylem | Bound | nd | 9.23±0.67 cd | 4114 ± 50 l | 74.19 ± 0.98 e | nd | 52.25 ± 1.38 c | nd | 29.60 ± 0.53 d | 42.59 ± 4.71 c | 16.31 ± 1.03 b |
| Free | 1799 ± 25 h | nd | 41.19 ± 2.33 a | 5.11 ± 0.16 a | 1.92 ± 0.39 a | 41.87 ± 1.86 b | 2540 ± 61 c | nd | nd | 13.77 ± 0.69 b | |
| Total | 1799 ± 25 h | 9.23±0.67 cd | 4155 ± 52 l | 79.31 ± 1.06 f | 1.92 ± 0.39 a | 94.12 ± 3.16 e | 2540 ± 61 c | 29.60 ± 0.53 d | 42.59 ± 4.71 c | 30.09 ± 0.50 c | |
| Phloem | Bound | 9.40 ± 0.60 a | 196.5 ± 2.4 k | 112.63 ± 1.58 c | 77.35 ± 1.17 ef | nd | nd | 32.49 ± 5.30 a | 30.67 ± 1.31 d | nd | nd |
| Free | 743.3 ± 24.2 f | 2.70 ± 0.09 a | 53.78 ± 3.12 ab | 6.84 ± 0.06 a | 5.41 ± 0.56 b | nd | 1426 ± 74 b | nd | nd | nd | |
| Total | 752.7 ± 24.8 f | 199.2 ± 2.5 j | 166.4 ± 2.1 d | 84.18 ± 1.12 g | 5.41 ± 0.56 b | nd | 1459 ± 79 b | 30.67 ± 1.31 d | nd | nd | |
| Petiole | Bound | 20.76 ± 1.24 ab | 4.87 ± 0.07 ab | 275.7 ± 3.3 e | 120.1 ± 3.8 j | nd | nd | nd | 10.31 ± 0.9 ab | 5.37 ± 0.10 a | nd |
| Free | 66.59 ± 1.86 cd | 7.11 ± 0.34 bc | 68.38 ± 0.44 abc | 8.98 ± 0.15 a | nd | nd | nd | 8.51 ± 0.39 ab | 5.16 ± 0.03 a | nd | |
| Total | 87.35 ± 2.17 d | 11.98 ± 0.42 d | 344.1 ± 3.7 f | 129.1 ± 3.7 k | nd | nd | nd | 18.82 ± 0.64 c | 10.53 ± 0.12 a | nd | |
| Leaf | Bound | nd | 8.36 ± 0.09 c | 367.74 ± 2.28 f | 90.41 ± 4.35 h | nd | nd | nd | 2.81 ± 0.13 a | 3.61 ± 0.03 a | nd |
| Free | 121.0 ± 2.6 e | 7.83 ± 0.71 c | 62.14 ± 0.59 ab | 7.73 ± 0.14 a | nd | nd | 2.03 ± 0.37 a | 254.6 ± 9.57 h | 97.39 ± 4.16 d | 2.04 ± 0.18 a | |
| Total | 121.0 ± 2.6 e | 16.19 ± 0.80 e | 429.9 ± 1.8 g | 98.13 ± 4.33 i | nd | nd | 2.03 ± 0.37 a | 257.4 ± 9.69 h | 101.0 ± 4.2 d | 2.04 ± 0.18 a | |
| Bud | Bound | 5.97 ± 0.11 a | 67.52 ± 1.44 f | 901.5 ± 10.4 h | 86.28 ± 4.5 gh | 2.68 ± 0.6 ab | nd | 3.47 ± 0.33 a | 9.23 ± 0.35 ab | 2.71 ± 0.03 a | nd |
| Free | 1586 ± 29 g | 80.29 ± 3.00 g | 101.2 ± 0.6 bc | 13.46 ± 0.17 b | nd | 82.23 ± 0.96 d | nd | 135.1 ± 8.0 f | 160.5 ± 12.7 e | nd | |
| Total | 1592 ± 30 g | 147.8 ± 2.9 i | 1003 ± 11 i | 99.75 ± 4.33 i | 2.68 ± 0.6 ab | 82.23 ± 0.96 d | 3.47 ± 0.33 a | 144.3 ± 8.4 g | 163.2 ± 12.7 e | nd | |
| A375 | KB | HepG2 | |
|---|---|---|---|
| Root | 3.11 ± 0.10 b * | 1.30 ± 0.07 b | 1.41 ± 0.07 a |
| Xylem | 2.84 ± 0.08 b | 1.64 ± 0.01 c | 0.96 ± 0.02 a |
| Phloem | 11.06 ± 0.43 c | 3.62 ± 0.22 d | 1.09 ± 0.07 a |
| Petiole | ND # | ND # | 2.28 ± 0.35 b |
| Leaf | ND # | ND # | 3.46 ± 0.54 c |
| Bud | 0.06 ± 0.01 a | 0.13 ± 0.01 a | 0.92 ± 0.14 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Qiu, C.; Chen, L.; Abbasi, A.M.; Guo, X.; Liu, R.H. Comparative Study of Phenolic Profiles, Antioxidant and Antiproliferative Activities in Different Vegetative Parts of Ramie (Boehmeria nivea L.). Molecules 2019, 24, 1551. https://doi.org/10.3390/molecules24081551
Wang H, Qiu C, Chen L, Abbasi AM, Guo X, Liu RH. Comparative Study of Phenolic Profiles, Antioxidant and Antiproliferative Activities in Different Vegetative Parts of Ramie (Boehmeria nivea L.). Molecules. 2019; 24(8):1551. https://doi.org/10.3390/molecules24081551
Chicago/Turabian StyleWang, Hong, Caisheng Qiu, Ling Chen, Arshad Mehmood Abbasi, Xinbo Guo, and Rui Hai Liu. 2019. "Comparative Study of Phenolic Profiles, Antioxidant and Antiproliferative Activities in Different Vegetative Parts of Ramie (Boehmeria nivea L.)" Molecules 24, no. 8: 1551. https://doi.org/10.3390/molecules24081551
APA StyleWang, H., Qiu, C., Chen, L., Abbasi, A. M., Guo, X., & Liu, R. H. (2019). Comparative Study of Phenolic Profiles, Antioxidant and Antiproliferative Activities in Different Vegetative Parts of Ramie (Boehmeria nivea L.). Molecules, 24(8), 1551. https://doi.org/10.3390/molecules24081551