Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes



Abstract
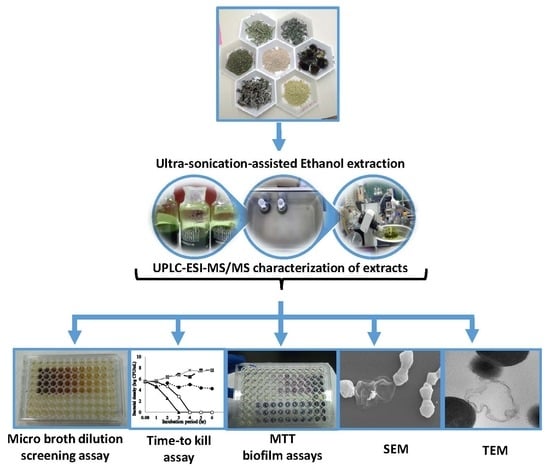
1. Introduction
2. Results
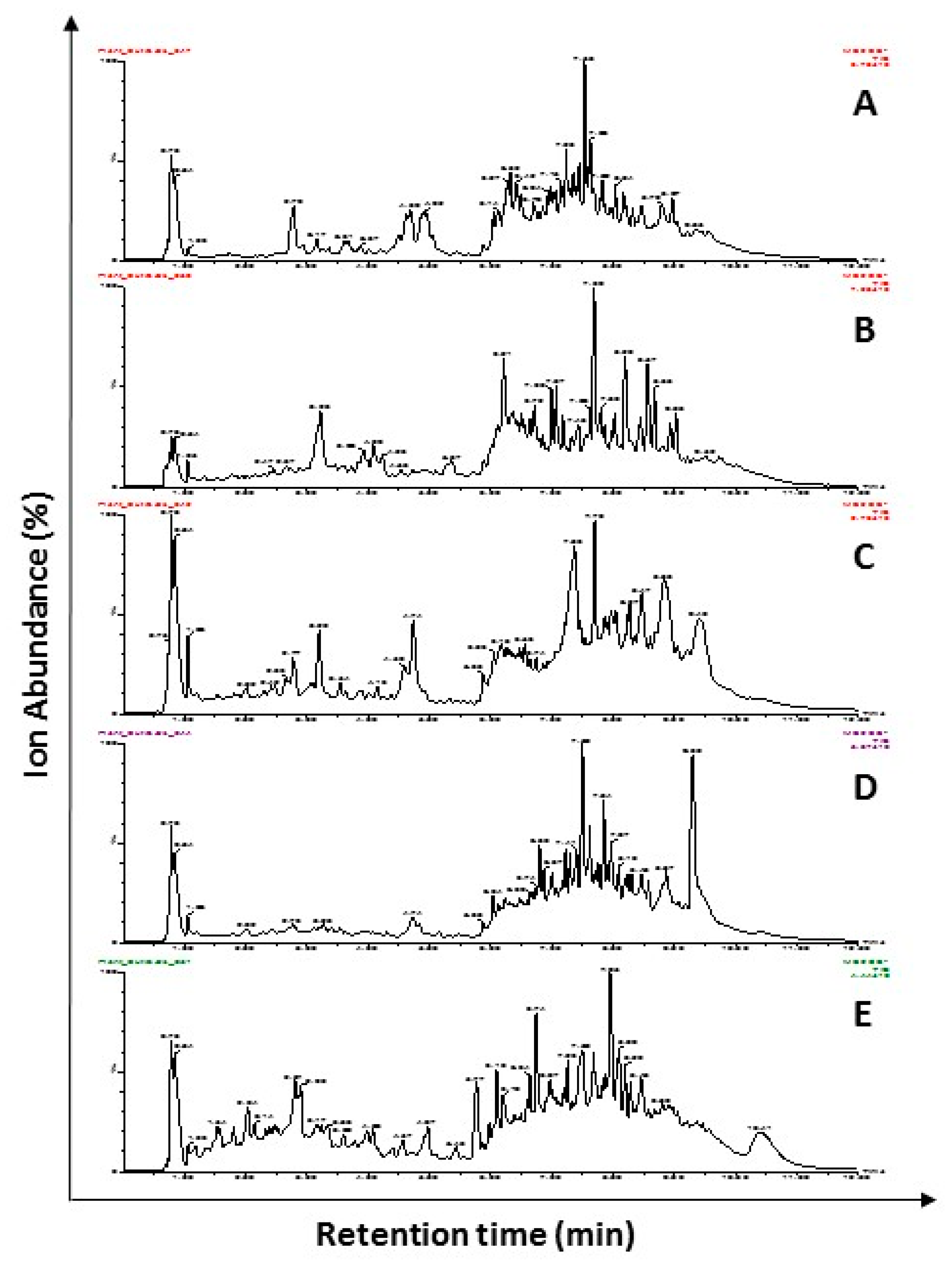
2.1. Characterization of Chemical Constituents Using UPLC-ESI-MS/MS
2.2. Inhibitory Effects of Plant Extracts against Planktonic S. pyogenes Growth
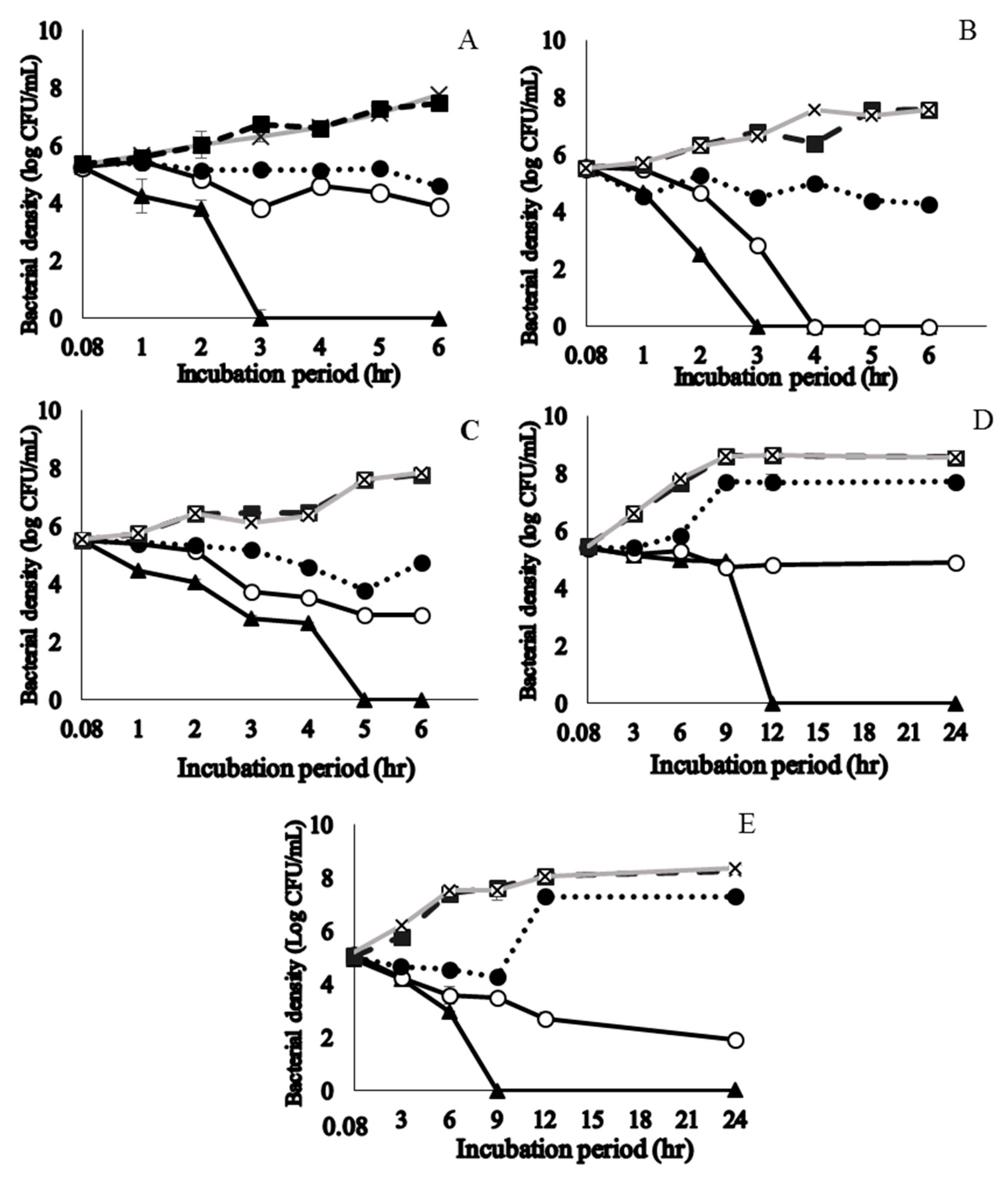
2.3. Time to Kill S. pyogenes
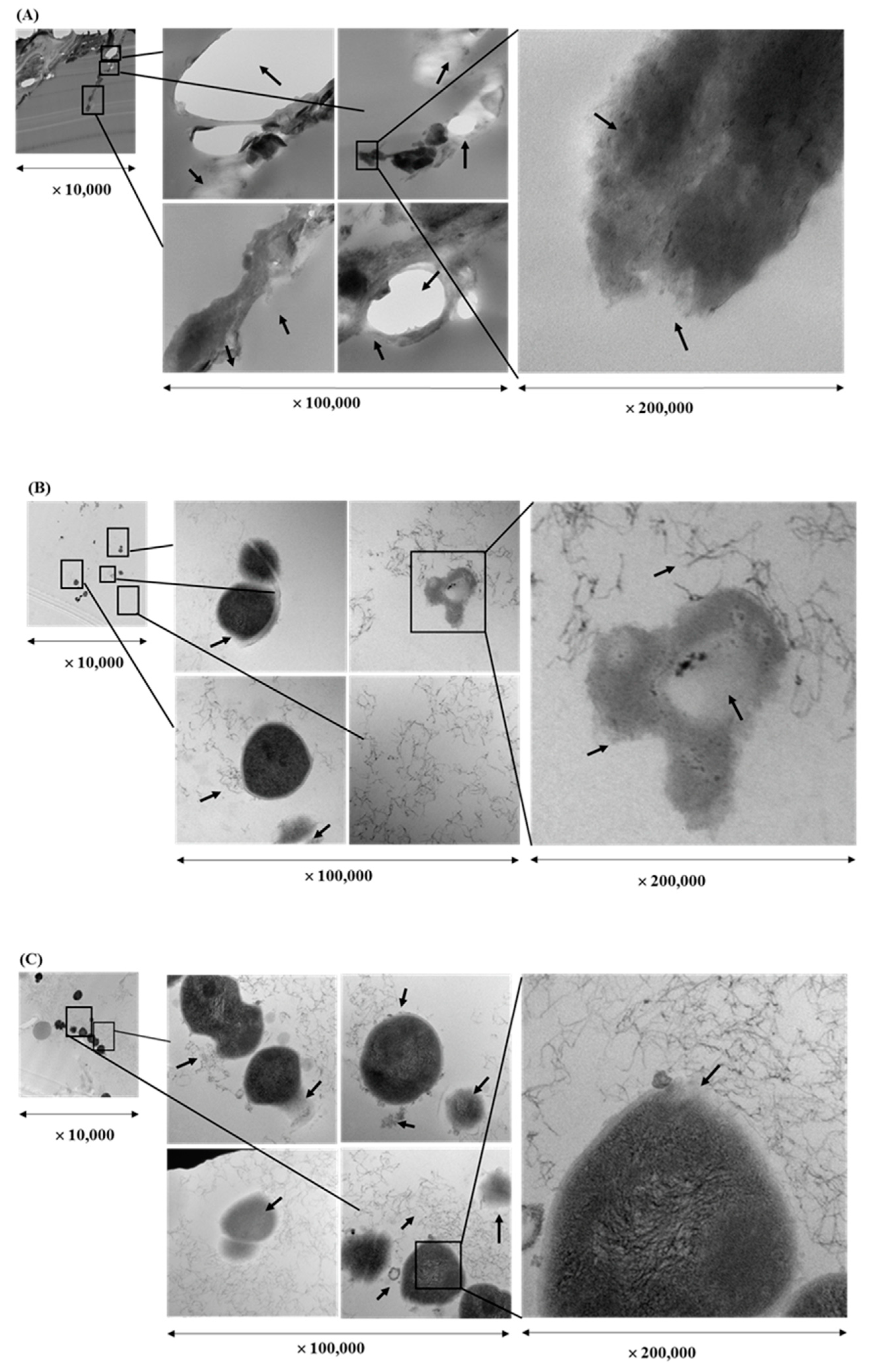
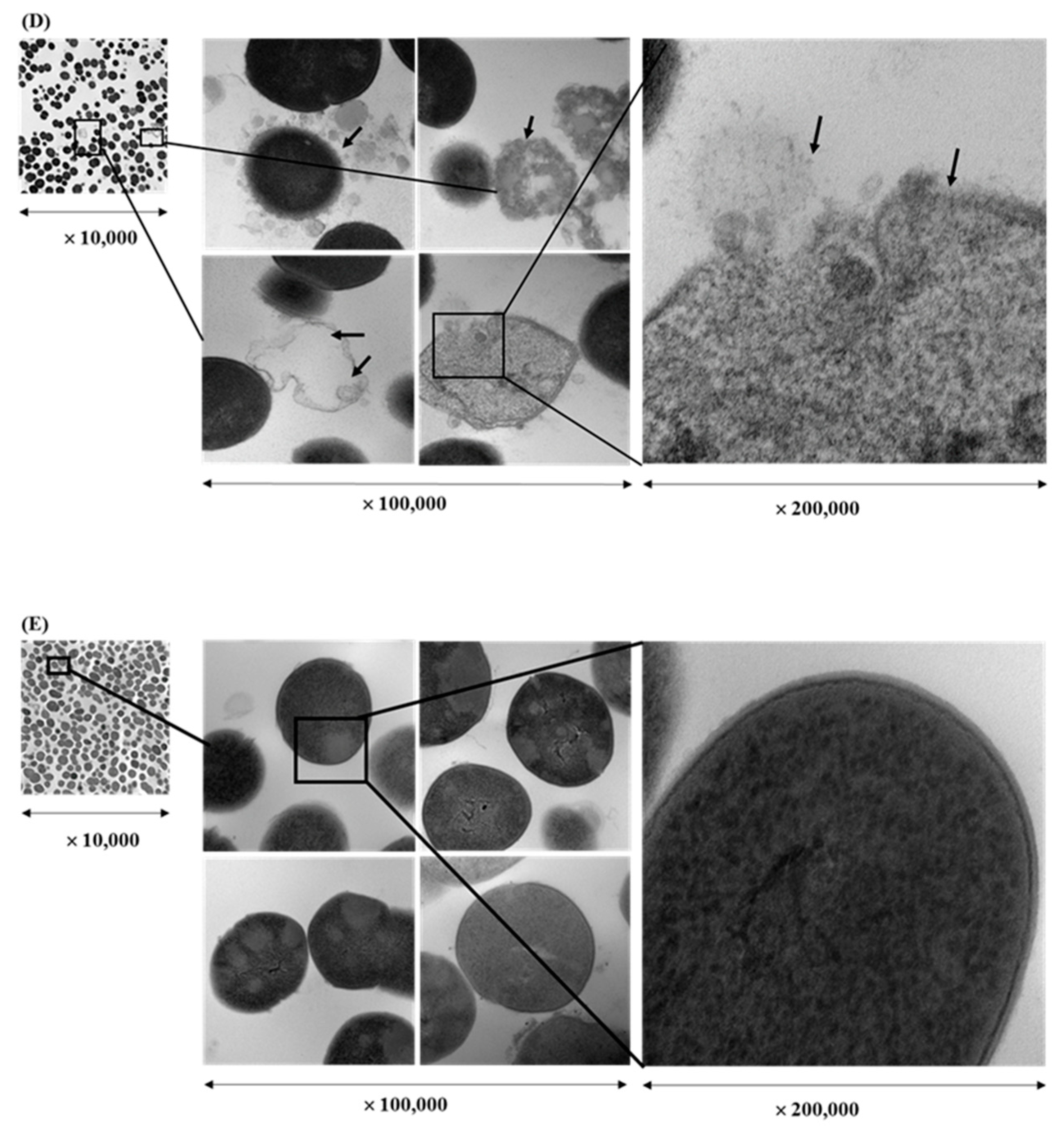
2.4. Transmission Electron Microscopy (TEM) Visualization of Fixed Biofilms of S. pyogenes
2.5. Anti-Biofilm Formation Activity
2.6. SEM Visualization of Fixed Biofilms of S. pyogenes
3. Discussion
3.1. Bactericidal Effect and Cell Integrity Changes by Plant Extracts
3.2. Anti-Biofilm Formation Ability of Plant Extracts
4. Materials and Methods
4.1. Collection of Plant Materials
4.2. Preparation of Ultrasonic-Assisted Ethanol Extracts (EE)
4.3. Phytochemical Analysis: Mass Spectrometric Characterization
4.4. Bacterial Strains and Culture Maintenance
4.5. Anti-Bacterial Activity
4.5.1. Determination of Minimum Inhibitory Concentrations (MICs)
4.5.2. Determination of Minimum Bactericidal Concentrations (MBCs)
4.6. Time to Kill Assay
4.7. TEM Observations of Bacterial Cell Wall Damage
4.8. Anti-Biofilm Formation Activity
4.8.1. Determination of Minimum Biofilm Inhibitory Concentration (MBIC)
4.8.2. SEM Visualization for Biofilm Morphology
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Bisno, A.L.; Peter, G.S.; Kaplan, E.L. Diagnosis of strep throat in adults: Are clinical criteria really good enough? Clin. Infect Dis. 2002, 35, 126–129. [Google Scholar] [CrossRef]
- Martin, J.M. The Mysteries of Streptococcal Pharyngitis. Curr. Treat Options Pediatr. 2015, 1, 180–189. [Google Scholar] [CrossRef]
- Shulman, S.T.; Bisno, A.L.; Clegg, H.W.; Gerber, M.A.; Kaplan, E.L.; Lee, G.; Martin, J.M.; Van Beneden, C. Clinical practice guideline for the diagnosis and management of group A streptococcal pharyngitis: 2012 update by the Infectious Diseases Society of America. Clin. Infect Dis. 2012, 55, 1279–1282. [Google Scholar] [CrossRef]
- Llor, C.; Bjerrum, L. Antimicrobial resistance: Risk associated with antibiotic overuse and initiatives to reduce the problem. Ther. Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Palla, A.H.; Khan, R.A.; Gilani, A.H.; Marra, F. Over prescription of antibiotics for adult pharyngitis is prevalent in developing countries but can be reduced using McIsaac modification of Centor scores: A cross-sectional study. BMC Pulm. Med. 2012, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Abachi, S.; Lee, S.; Rupasinghe, H.P.V. Molecular mechanisms of inhibition of streptococcus species by phytochemicals. Molecules 2016, 21, 215. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.M.; Anderson, M.; Schoop, S.R.; Hudson, J.B. Bactericidal and anti-inflammatory properties of a standardized Echinacea extract (Echinaforce): Dual actions against respiratory bacteria. Phytomedicine 2010, 17, 563–568. [Google Scholar] [CrossRef]
- Zuo, G.-Y.; Yang, C.-X.; Han, J.; Li, Y.-Q.; Wang, G.-C. Synergism of prenylflavonoids from Morus alba root bark against clinical MRSA isolates. Phytomedicine 2018, 39, 93–99. [Google Scholar] [CrossRef]
- Wijesundara, N.M.; Rupasinghe, H.P.V. Essential oils from Origanum vulgare and Salvia officinalis exhibit antibacterial and anti-biofilm activities against Streptococcus pyogenes. Micro. Pathogenesis 2018, 117, 118–127. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.E.; Ghareeb, D.A.; Sarhan, E.E.; Abu-Serie, M.M.; El Demellawy, M.A. In vitro biological assessment of Berberis vulgaris and its active constituent, berberine: Antioxidants, anti-acetylcholinesterase, anti-diabetic and anticancer effects. BMC Complement. Altern. Med. 2013, 13, 218. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Aligiannis, N.; Halabalaki, M.; Skaltsounis, A.-L.; Glowniak, K.; Kalpoutzakis, E. Influence of extraction procedures on phenolic content and antioxidant activity of Cretan barberry herb. Food Chem. 2013, 138, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Suau, R.; Rico, R.; López-Romero, J.M.; Nájera, F.; Cuevas, A. Isoquinoline alkaloids from Berberis Vulgaris subsp. Australis. Phytochemistry 1998, 49, 2545–2549. [Google Scholar] [CrossRef]
- Lee, K.-G.; Shibamoto, T. Antioxidant property of aroma extract isolated from clove buds [Syzygium aromaticum (L.) Merr. et Perry]. Food Chem. 2001, 74, 443–448. [Google Scholar] [CrossRef]
- de Oliveira, M.S.; da Costa, W.A.; Pereira, D.S.; Botelho, J.R.S.; de Alencar Menezes, T.O.; de Aguiar Andrade, E.H.; da Silva, S.H.M.; da Silva Sousa Filho, A.P.; de Carvalho, R.N. Chemical composition and phytotoxic activity of clove (Syzygium aromaticum) essential oil obtained with supercritical CO2. J. Supercrit. Fluid. 2016, 118, 185–193. [Google Scholar] [CrossRef]
- Cao, J.-L.; Wei, J.-C.; Hu, Y.-J.; He, C.-W.; Chen, M.-W.; Wan, J.-B.; Li, P. Qualitative and quantitative characterization of phenolic and diterpenoid constituents in Danshen (Salvia miltiorrhiza) by comprehensive two-dimensional liquid chromatography coupled with hybrid linear ion trap Orbitrap mass. J. Chromatogr. A 2016, 1427, 79–89. [Google Scholar] [CrossRef]
- Yang, M.; Liu, A.H.; Guan, S.H.; Sun, J.H.; Xu, M.; Guo, D. Characterization of tanshinones in the roots of Salvia miltiorrhiza (Dan-shen) by high-performance liquid chromatography with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Sp. 2006, 20, 1266–1280. [Google Scholar] [CrossRef]
- Hudec, J.; Burdová, M.; Kobida, L.U.; Komora, L.; Macho, V.; Kogan, G.; Turianica, I.; Kochanová, R.; Ložek, O.; Habán, M.; et al. Antioxidant Capacity Changes and Phenolic Profile of Echinacea purpurea, Nettle (Urtica dioica L.), and Dandelion (Taraxacum officinale) after Application of Polyamine and Phenolic Biosynthesis Regulators. J. Agric. Food Chem. 2007, 55, 5689–5696. [Google Scholar] [CrossRef]
- Kim, H.-O.; Durance, T.D.; Scaman, C.H.; Kitts, D.D. Retention of Caffeic Acid Derivatives in Dried Echinacea purpurea. J. Agric. Food Chem. 2000, 48, 4182–4186. [Google Scholar] [CrossRef]
- Tsai, Y.-L.; Chiou, S.-Y.; Chan, K.-C.; Sung, J.-M.; Lin, S.-D. Caffeic acid derivatives, total phenols, antioxidant and antimutagenic activities of Echinacea purpurea flower extracts. LWT-Food Sci. Technol. 2012, 46, 169–176. [Google Scholar] [CrossRef]
- Vimalanathan, S.; Kang, L.; Amiguet, V.T.; Livesey, J.; Arnason, J.T.; Hudson, J. Echinacea purpurea aerial parts contain multiple antiviral compounds. Pharm. Biol. 2005, 43, 740–745. [Google Scholar] [CrossRef]
- Hsouna, A.B.; Hamdi, N. Phytochemical composition and antimicrobial activities of the essential oils and organic extracts from pelargonium graveolens growing in Tunisia. Lipids Health Dis. 2012, 11, 167. [Google Scholar] [CrossRef]
- Račková, L.; Cupáková, M.; Tažký, A.; Mičová, J.; Kolek, E.; Košt’álová, D. Redox properties of ginger extracts: Perspectives of use of Zingiber officinale Rosc. as antidiabetic agent. Interdiscip. Toxicol. 2013, 6, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, Y.S.; Aktumsek, A.; Duran, A. Studies on antioxidant activity, volatile compound and fatty acid composition of different parts of Glycyrrhiza echinata L. EXCLI J. 2012, 11, 178–187. [Google Scholar]
- Montoro, P.; Maldini, M.; Russo, M.; Postorino, S.; Piacente, S.; Pizza, C. Metabolic profiling of roots of liquorice (Glycyrrhiza glabra) from different geographical areas by ESI/MS/MS and determination of major metabolites by LC-ESI/MS and LC-ESI/MS/MS. J. Pharm. Biomed. 2011, 54, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Simons, R.; Vincken, J.P.; Bakx, E.J.; Verbruggen, M.A.; Gruppen, H. A rapid screening method for prenylated flavonoids with ultra-high-performance liquid chromatography/electrospray ionization mass spectrometry in licorice root extracts. Rapid Commun. Mass Sp. 2009, 23, 3083–3093. [Google Scholar] [CrossRef] [PubMed]
- Koldas, S.; Demirtas, I.; Ozen, T.; Demirci, M.A.; Behcet, L. Phytochemical screening, anticancer and antioxidant activities of Origanum vulgare L. ssp viride (Boiss.) Hayek, a plant of traditional usage. J. Sci. Food Agric. 2015, 95, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Ličina, B.Z.; Stefanović, O.D.; Vasić, S.M.; Radojević, I.D.; Dekić, M.S.; Čomić, L.R. Biological activities of the extracts from wild growing Origanum vulgare L. Food Control 2013, 33, 498–504. [Google Scholar]
- Milos, M.; Mastelic, J.; Jerkovic, I. Chemical composition and antioxidant effect of glycosidically bound volatile compounds from oregano (Origanum vulgare L. ssp. hirtum). Food Chem. 2000, 71, 79–83. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.; Saraiva, J.A.; Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare) extracts and essential oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Maalej, A.; Bouallagui, Z.; Hadrich, F.; Isoda, H.; Sayadi, S. Assessment of Olea europaea L. fruit extracts: Phytochemical characterization and anticancer pathway investigation. Biomed. Pharmacother. 2017, 90, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Exarchou, V.; Nenadis, N.; Tsimidou, M.; Gerothanassis, I.P.; Troganis, A.; Boskou, D. Antioxidant Activities and Phenolic Composition of Extracts from Greek Oregano, Greek Sage, and Summer Savory. J. Agric. Food Chem. 2002, 50, 5294–5299. [Google Scholar] [CrossRef]
- Hamidpour, M.; Hamidpour, R.; Hamidpour, S.; Shahlari, M. Chemistry, Pharmacology, and Medicinal Property of Sage (Salvia) to Prevent and Cure Illnesses such as Obesity, Diabetes, Depression, Dementia, Lupus, Autism, Heart Disease, and Cancer. J. Tradit. Complement. Med. 2014, 4, 82–88. [Google Scholar] [CrossRef]
- Šulniūtė, V.; Pukalskas, A.; Venskutonis, P.R. Phytochemical composition of fractions isolated from ten Salvia species by supercritical carbon dioxide and pressurized liquid extraction methods. Food Chem. 2017, 224, 37–47. [Google Scholar] [CrossRef]
- Veličković, D.T.; Milenović, D.M.; Ristić, M.S.; Veljković, V.B. Kinetics of ultrasonic extraction of extractive substances from garden (Salvia officinalis L.) and glutinous (Salvia glutinosa L.) sage. Ultrason. Sonochem. 2006, 13, 150–156. [Google Scholar] [CrossRef]
- Yilar, M.; Kadioglu, I.; Telci, I. Chemical Composition and Antifungal Activity of Salvia Officinalis (L.), S. Cryptantha (Montbret Et Aucher Ex Benth.), S. Tomentosa (Mill.) Plant Essential Oils and Extracts. Fresen. Environ. Bull. 2018, 27, 1695–1706. [Google Scholar]
- Barsett, H.; Smestad Paulsen, B. Separation, isolation and characterization of acidic polysaccharides from the inner bark of Ulmus glabra Huds. Carbohyd. Polym. 1992, 17, 137–144. [Google Scholar] [CrossRef]
- Al Hashmi, L.S.; Hossain, M.A.; Weli, A.M.; Al-Riyami, Q.; AlSabahi, J.N. Gas chromatography–mass spectrometry analysis of different organic crude extracts from the local medicinal plant of Thymus vulgaris L. Asian Pac. J. Trop. Med. 2013, 3, 69–73. [Google Scholar] [CrossRef]
- Habashy, N.H.; Abu Serie, M.M.; Attia, W.E.; Abdelgaleil, S.A.M. Chemical characterization, antioxidant and anti-inflammatory properties of Greek Thymus vulgaris extracts and their possible synergism with Egyptian Chlorella vulgaris. J. Funct. Foods 2018, 40, 317–328. [Google Scholar] [CrossRef]
- Köksal, E.; Bursal, E.; Gülçin, İ.; Korkmaz, M.; Çağlayan, C.; Gören, A.C.; Alwasel, S.H. Antioxidant activity and polyphenol content of Turkish thyme (Thymus vulgaris) monitored by liquid chromatography and tandem mass spectrometry. Int. J. Food Prop. 2017, 20, 514–525. [Google Scholar] [CrossRef]
- Pereira, O.R.; Peres, A.M.; Silva, A.M.S.; Domingues, M.R.M.; Cardoso, S.M. Simultaneous characterization and quantification of phenolic compounds in Thymus x citriodorus using a validated HPLC–UV and ESI–MS combined method. Food Res. Int. 2013, 54, 1773–1780. [Google Scholar] [CrossRef]
- Zhang, B.; Wijesundara, N.M.; Abbey, L.; Rupasinghe, H.P.V. Growing medium amendments effect on growth, secondary metabolites and anti-streptococcal activity of two species of Plectranthus. J. Appl. Res. Med. Aromat. Plants 2017, 5, 53–59. [Google Scholar] [CrossRef]
- Mohd, A.; Rosina, K.; Rupasinghe, H.P.V. Application of Medicinal Plants as a Source for Therapeutic Agents Against Streptococcus pyogenes Infections. Curr. Drug Metab. 2018, 19, 695–703. [Google Scholar]
- Zacchino, S.A.; Butassi, E.; Liberto, M.D.; Raimondi, M.; Postigo, A.; Sortino, M. Plant phenolics and terpenoids as adjuvants of antibacterial and antifungal drugs. Phytomedicine 2017, 37, 27–48. [Google Scholar] [CrossRef] [PubMed]
- Wijesundara, N.M.; Sekhon-Loodu, S.; Rupasinghe, H.P.V. Phytochemical-rich medicinal plant extracts suppress bacterial antigens-induced inflammation in human tonsil epithelial cells. Peer J. 2017, 5, e3469. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.M.; Ramaiah, S. Pharmacological importance of Echinacea purpurea. Int. J. Pharm. Biol. Sci. 2011, 2, 304–314. [Google Scholar]
- Fournomiti, M.; Kimbaris, A.; Mantzourani, I.; Plessas, S.; Theodoridou, I.; Papaemmanouil, V.; Kapsiotis, I.; Panopoulou, M.; Stavropoulou, E.; Bezirtzoglou, E.E.; et al. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microb. Ecol. Health Dis. 2015, 26, 23289. [Google Scholar] [CrossRef]
- Abu-Darwish, M.S.; Cabral, C.; Ferreira, I.V.; Goncalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Al-bdour, T.H.; Salgueiro, L. Essential oil of common sage (Salvia officinalis L.) from Jordan: Assessment of safety in mammalian cells and its antifungal and anti-inflammatory potential. Biomed. Res. Int. 2013, 2013, 538940. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods--a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, J.; Li, Y.J.; Zheng, Y.F.; Li, P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Sedighinia, F.; Safipour Afshar, A.; Soleimanpour, S.; Zarif, R.; Asili, J.; Ghazvini, K. Anti-bacterial activity of Glycyrrhiza glabra against oral pathogens: An in vitro study. Avicenna J. Phytomed. 2012, 2, 118–124. [Google Scholar] [PubMed]
- Zadeh, J.B.; Kor, Z.M.; Goftar, M.K. Licorice (Glycyrrhiza glabra Linn) as a valuable medicinal plant. Int. J. Adv. Biol. Biomed. Res. 2014, 1, 196–199. [Google Scholar]
- Nzeako, B.C.; Al-Kharousi, Z.S.; Al-Mahrooqui, Z. Antimicrobial activities of clove and thyme extracts. Sultan Qaboos Univ. Med. J. 2006, 6, 33–39. [Google Scholar] [PubMed]
- Campos, F.M.; Couto, J.A.; Figueiredo, A.R.; Toth, I.V.; Rangel, A.O.; Hogg, T.A. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Teethaisong, Y.; Pimchan, T.; Srisawat, R.; Hobbs, G.; Eumkeb, G. Boesenbergia rotunda (L.) Mansf. extract potentiates the antibacterial activity of some β-lactams against β-lactam-resistant staphylococci. J. Glob. Antimicrob. Re. 2018, 12, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Terao, Y.; Okuni, H.; Ninomiya, K.; Sakata, H.; Ikebe, K.; Maeda, Y.; Kawabata, S. Biofilm formation or internalization into epithelial cells enable Streptococcus pyogenes to evade antibiotic eradication in patients with pharyngitis. Microb. Pathog. 2011, 51, 58–68. [Google Scholar] [CrossRef]
- Post, J.C.; Stoodley, P.; Hall-Stoodley, L.; Ehrlich, G.D. The role of biofilms in otolaryngologic infections. Curr. Opin. Otolaryngol. Head Neck Surg. 2004, 12, 185–190. [Google Scholar] [CrossRef]
- Conley, J.; Olson, M.E.; Cook, L.S.; Ceri, H.; Phan, V.; Davies, H.D. Biofilm formation by group a streptococci: Is there a relationship with treatment failure? J. Clin. Microbiol. 2003, 41, 4043–4048. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Manetti, A.G.; Zingaretti, C.; Falugi, F.; Capo, S.; Bombaci, M.; Bagnoli, F.; Gambellini, G.; Bensi, G.; Mora, M.; Edwards, A.M.; et al. Streptococcus pyogenes pili promote pharyngeal cell adhesion and biofilm formation. Mol. Microbiol. 2007, 64, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Darsini, D.T.P.; Srinivasan, P.; Guna, G.; Manimekalai, K.; Dineshbabu, J. In vitro anti-biofilm activity of Piper longum and Piper nigrum against clinical isolates of S. pyogenes isolated from pharyngitis patients. Int. J. Pharm. 2015, 6, 122–132. [Google Scholar]
- Mutalib, L.Y.; Nuraddin, S.M.; Aka, S.T.H. Phytochemical screening, antibacterial and antibiofilm evaluation of Lagenaria siceraria fruit growing in Kurdistan region\Iraq. J. Pharmacogn Phytochem. 2015, 4, 45–49. [Google Scholar]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically: Approved Standard; Clinical and Laboratory Standards Institute: Valley, Wayne, PA, USA, 2006. [Google Scholar]
Sample Availability: No samples of this study is available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract Name | Potential Major Phytochemicals ([M − H+]−, RT) | References |
|---|---|---|
| Barberry roots | Jatrorrhizine (337.38, 8.04), Palmatine (341.40, 9.28), Berbamine (608.7, 7.79), Quercetin (301.23, 7.49), Rutin (609.52, 4.73), and Kaempferol (285.23, 6.50). | [12,13,14] |
| Clove flower buds | Eugenol (163.20, 4.40), β-Ocimene (135.25, 4.25), p-Allyl phenol (133.17, 0.84), and Kaempferol (285.23, 6.50). | [15,16] |
| Danshen roots * | NI in this study Danshenxinkun A, Protocatechuic acid, Caffeic acid, Rosmarinic acid, Tanshinone VI, 17-Hydroxycryptotanshinone, Salvianolic acid, and Salvianolic acid A. | [17,18] |
| Purple coneflower flowers | Caftaric acid (311.23, 2.44), Chlorogenic acid (353.31, 2.78), Caffeic acid (179.16, 3.27), Cynarin (515.46, 6.02), Echinacoside (785.73, 6.56), and Cichoric acid (473.37, 4.78). | [19,20,21,22] |
| Purple coneflower leaves | Chlorogenic acid (353.31, 2.78), Caffeic acid (179.16, 3.27), Cynarin (515.46, 6.02) Echinacoside (785.73, 6.56), and Cichoric acid (473.37, 4.78). | [19,22] |
| Purple coneflower stems | Caftaric acid (311.23, 2.44), Chlorogenic acid (353.31, 2.78) and Cichoric acid (473.37, 4.78), Quercetin (301.23. 7.49), Caryophyllene oxide (375.45, 7.40), and Liquirtin (419.39, 6.21). | [19,22] |
| Geranium leaves | Geraniol (151.24, 6.62). | [23] |
| Ginger rhizome | Gingerol (273.38, 7.18), Galanolactone (317.45, 8.94), Zingerone (193.22, 4.96), α-Humulene (203. 24, 2.50), and α or β-Caryophyllene (203.36, 2.52). | [24] |
| Licorice root | Naringin (579.54, 6.28), Asparegene (131.12, 1.63), Rosmarinic acid (359.31, 6.23) and Liquirtin (419.39, 6.21). | [25,26,27] |
| Oregano flowering shoots | p-Cymene and/or Borneol and/or 1,8 Cineole, (153.24, 2.20), Thymohydroquinone (165.22, 7.17), Rosmarinic acid (359.31, 6.23) and Naringin (579.54, 6.28). | [11,28,29,30,31] |
| Olive leaves * | NI in this study. Gallic acid, Hydroxytyrosol, Tyrosol, Caffeic acid, p-Coumaric, Oleuropein, Luteolin, and Quercetin. | [32] |
| Sage leaves | Borneol and/or Linalool and/or α-Terpineol and/or β-Pinene (135.24, 3.05 or 4.01), Asparagine (131.12, 3.65), γ-Terpinene and/or Mycrene and/or β-Pinene and/or α-Pinene (135.24, 4.24 and Kaempferol (285.23, 6.50). | [11,33,34,35,36,37] |
| Slippery elm inner barks | Oleanolic acid (455.71, 7.2), Ursolic acid and/or Betulinic acid (455.71, 7.50 or 9.6) and β-carotene (535.87, 5.29). | [38] |
| Thyme flowering shoots | Thymol and/or Carvacrol (149.21, 6.65), Thymohydroquinone (165.22, 7.17), γ-Terpinene, Mycrene, and α-Pinene (135.24, 4.24) and Kaempferol (285.23, 6.50). | [39,40,41,42] |
| Plant Source/Reference | ATCC 19615 | ATCC 49399 | Clinical Isolate | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC (μg/mL) | MBC (μg/mL) | MBC/MIC | MIC (μg/mL) | MBC (μg/mL) | MBC/MIC | MIC (μg/mL) | MBC (μg/mL) | MBC/MIC | |
| Barberry Roots | 250 | 500 | 2 | 250 | 500 | 2 | 250 | 500 | 2 |
| Clove Flower Buds | 500 | 1000 | 2 | 500 | 1000 | 2 | 500 | 1000 | 2 |
| Dan shen Roots | 250 | >250 | - | 250 | >250 | - | NA | NA | - |
| Purple Coneflower Flower | 62.5 | 125 | 2 | 62.5 | 125 | 2 | 62.5 | 125 | 2 |
| Purple Coneflower Leaves | >1000 | >1000 | - | >1000 | >1000 | - | NA | NA | - |
| Purple Coneflower Stem | 62.5 | 125 | 2 | 62.5 | 125 | 2 | 125 | 250 | 2 |
| Geranium Leaves | >1000 | >1000 | - | >1000 | >1000 | - | NA | NA | - |
| Ginger Rhizomes | >1000 | >1000 | - | >1000 | >1000 | - | NA | NA | - |
| Licorice Roots | 62.5 | 125 | 2 | 62.5 | 125 | 2 | 62.5 | 125 | 2 |
| Olive Leaves | >1000 | >1000 | - | >1000 | >1000 | - | NA | NA | - |
| Oregano Flowering Shoots | 500 | 1000 | 2 | 500 | 1000 | 2 | 500 | 1000 | 2 |
| Sage Leaves | 62.5 | 125 | 2 | 62.5 | 125 | 2 | 62.5 | 125 | 2 |
| Slippery elm Inner barks | 62.5 | 125 | 1 | 62.5 | 125 | 2 | 62.5 | 125 | 1 |
| Thyme Flowering shoots | 500 | 1000 | 2 | 500 | 1000 | 500 | 1000 | - | |
| Penicillin | 0.008 | 0.016 | 2 | 0.008 | 0.016 | 2 | 0.008 | 0.016 | 2 |
| Plant | MBIC (μg/mL) | ||
|---|---|---|---|
| ATCC 19615 | ATCC 49399 | Clinical | |
| Licorice Roots | 250 (4 × MIC) | 250 (4 × MIC) | 62.5 (1 × MIC) |
| Purple Coneflower Stems | 125 (2 × MIC) | 250 (4 × MIC) | 250 (2 × MIC) |
| Purple Coneflower Flowers | 125 (2 × MIC) | 31.5 (1 × MIC) | 62.5 (1 × MIC) |
| Sage Leaves | 125 (2 × MIC) | 125 (2 × MIC) | 125 (2 × MIC) |
| Slippery elm Inner Bark | 62.5 (1 × MIC) | 62.5 (1 × MIC) | 125 (2 × MIC) |
| Penicillin G | 0.0156 (2 × MIC) | 0.0625 (8 × MIC) | 0.0625 (8 × MIC) |
| Plant Name | Family | Parts Used | Voucher No. | GPS Location of Harvested Area | |
|---|---|---|---|---|---|
| Common | Botanical | ||||
| Barberry | Berberis vulgaris L. | Berberidaceae | Root | - | Purchased |
| Clove | Syzygium aromaticum L. | Myrtaceae | Flower bud | - | Purchased |
| Danshen | Salvia miltiorrhiza Bunge. | Lamiaceae | Roots | - | Purchased |
| Echinacea/Purple cone-flower | Echinacea purpurea L. | Asteraceae | Leaves, stem, flower | 13009 | 45°22′20.8″ N 63°15′43.8″ W |
| Ginger | Zingiber officinale L. | Zingiberaceae | Rhizome | - | Purchased |
| Licorice | Glycyrrhiza glabra L. | Papilionaceae | Root | - | Purchased |
| Oregano | Origanum vulgare L. | Lamiaceae | Flowering shoots and leaves | 13012 | 45°22′23.3″ N 63°15′45.2″ W |
| Olive | Olea europeus L. | Oleaceae | Leaves | - | Purchased |
| Rose geranium | Pelargonium graveolens L. | Geraniaceae | Leaves | 13010 | 45°22′23.3″ N 63°15′45.2″ W |
| Sage | Salvia officinalis L. | Lamiaceae | leaves, root | 13011 | 45°22′23.3″ N 63°15′45.2″ W |
| Slippery elm | Ulmus rubra Muhl. | Ulmaceae | Inner bark | - | Purchased |
| Thyme | Thymus vulgaris L. | Lamiaceae | Flowering shoots and leaves | 13013 | 45°22′23.3″ N 63°15′45.2″ W |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijesundara, N.M.; Rupasinghe, H.P.V. Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes. Molecules 2019, 24, 1165. https://doi.org/10.3390/molecules24061165
Wijesundara NM, Rupasinghe HPV. Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes. Molecules. 2019; 24(6):1165. https://doi.org/10.3390/molecules24061165
Chicago/Turabian StyleWijesundara, Niluni M., and H. P. Vasantha Rupasinghe. 2019. "Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes" Molecules 24, no. 6: 1165. https://doi.org/10.3390/molecules24061165
APA StyleWijesundara, N. M., & Rupasinghe, H. P. V. (2019). Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes. Molecules, 24(6), 1165. https://doi.org/10.3390/molecules24061165