

RETRACTED: Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
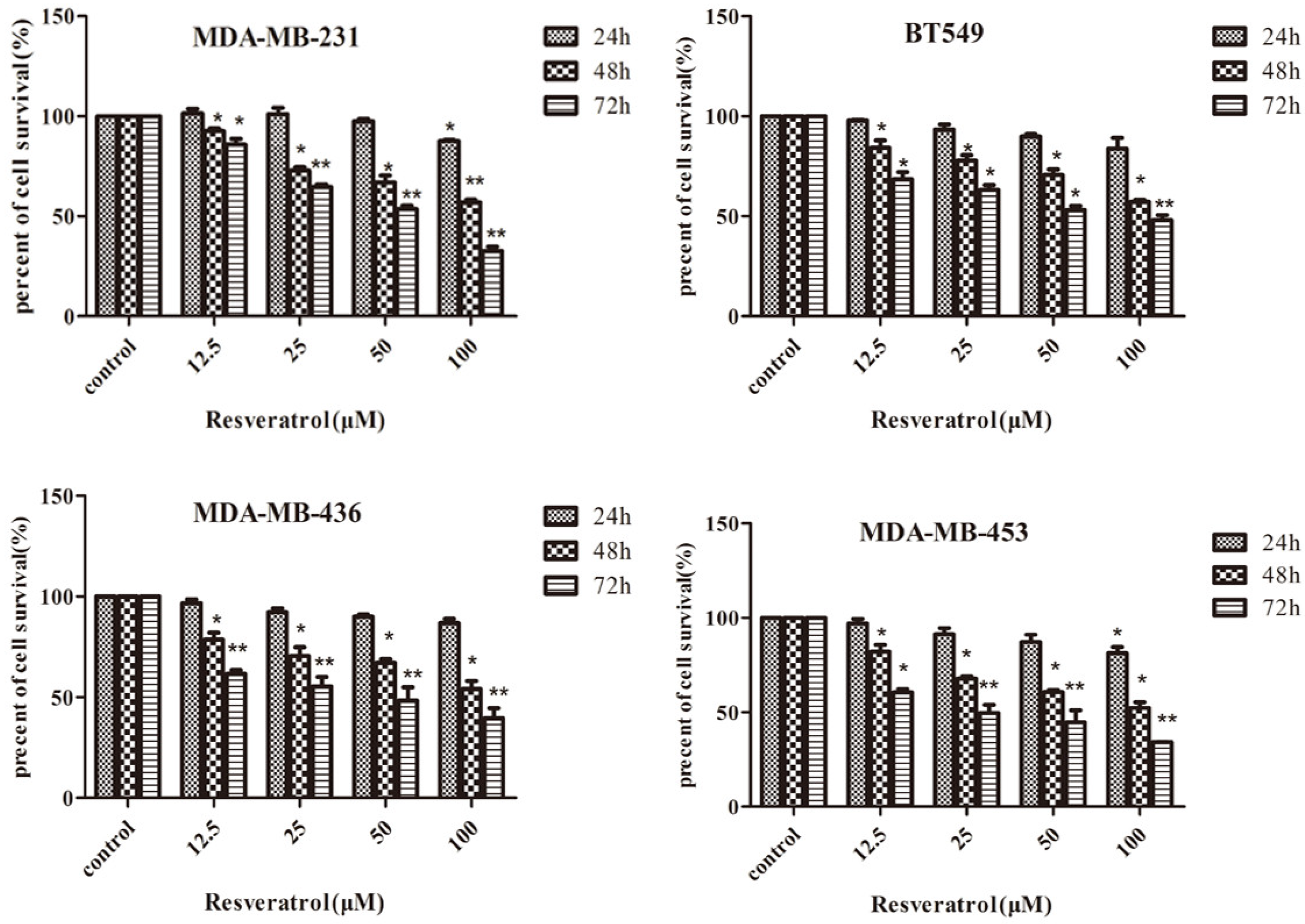
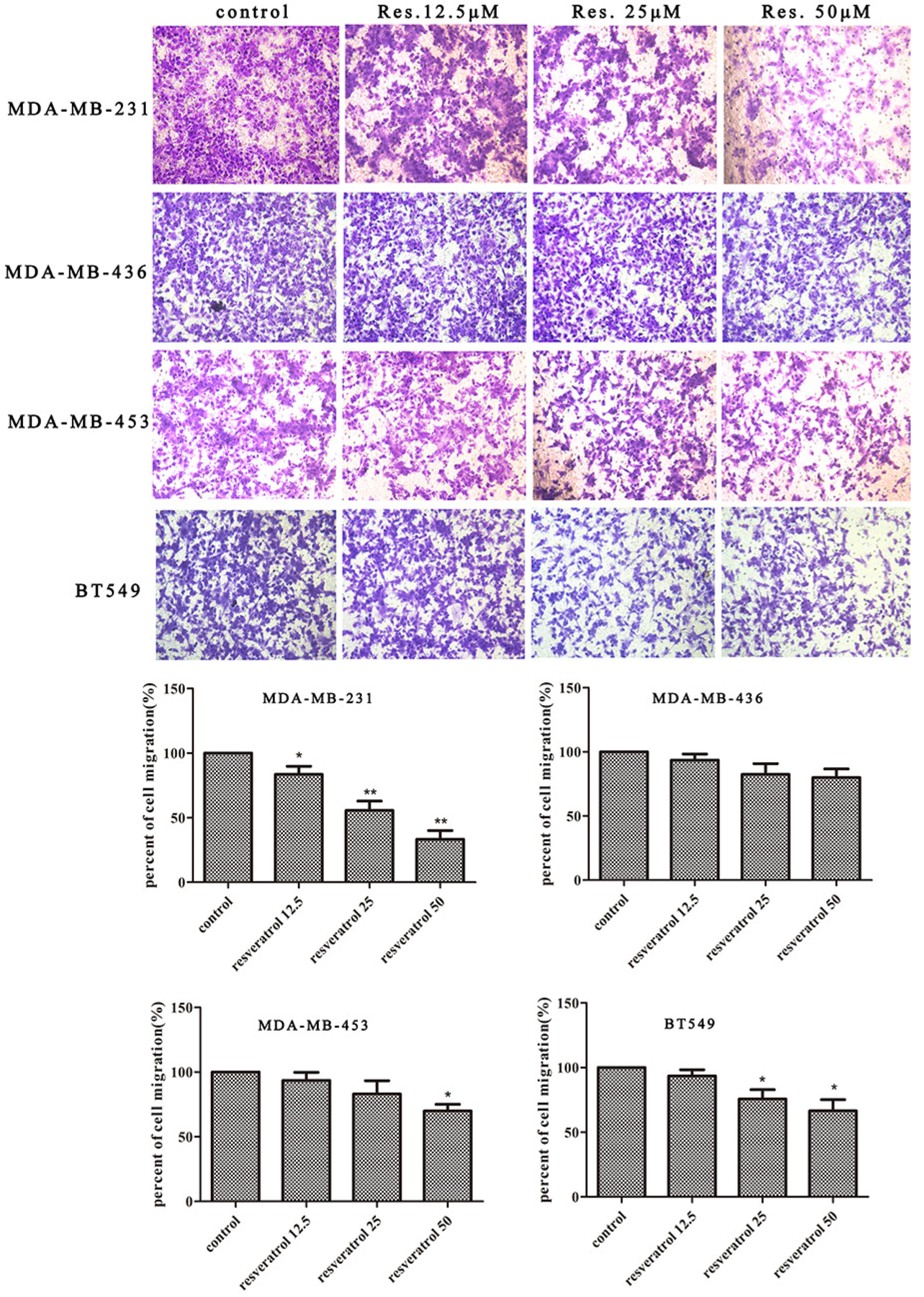
2.1. Resveratrol Inhibits the Migration of Breast Cancer Cells
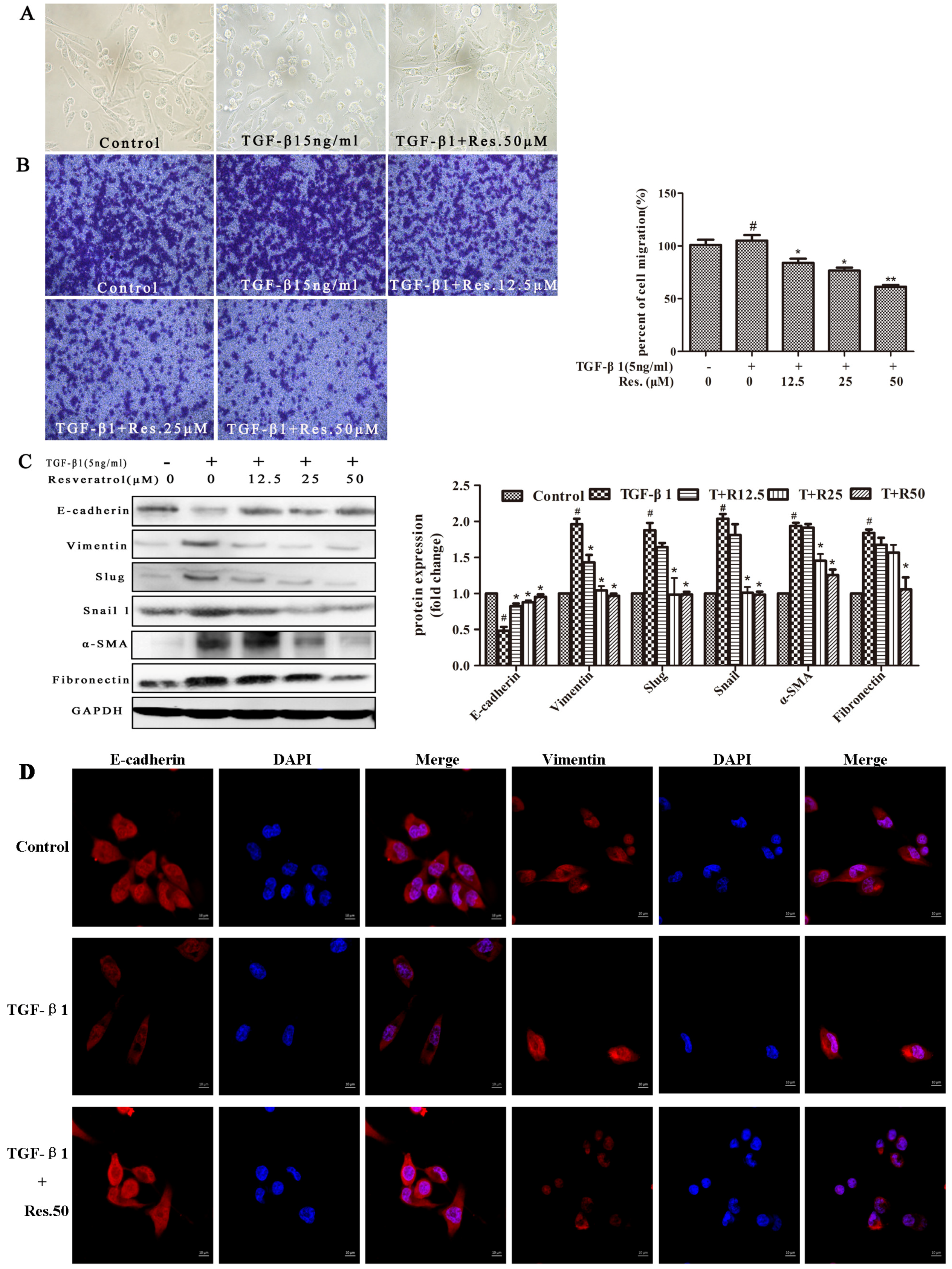
2.2. Resveratrol Reverses TGF-β1-Induced EMT in MDA231 Cells
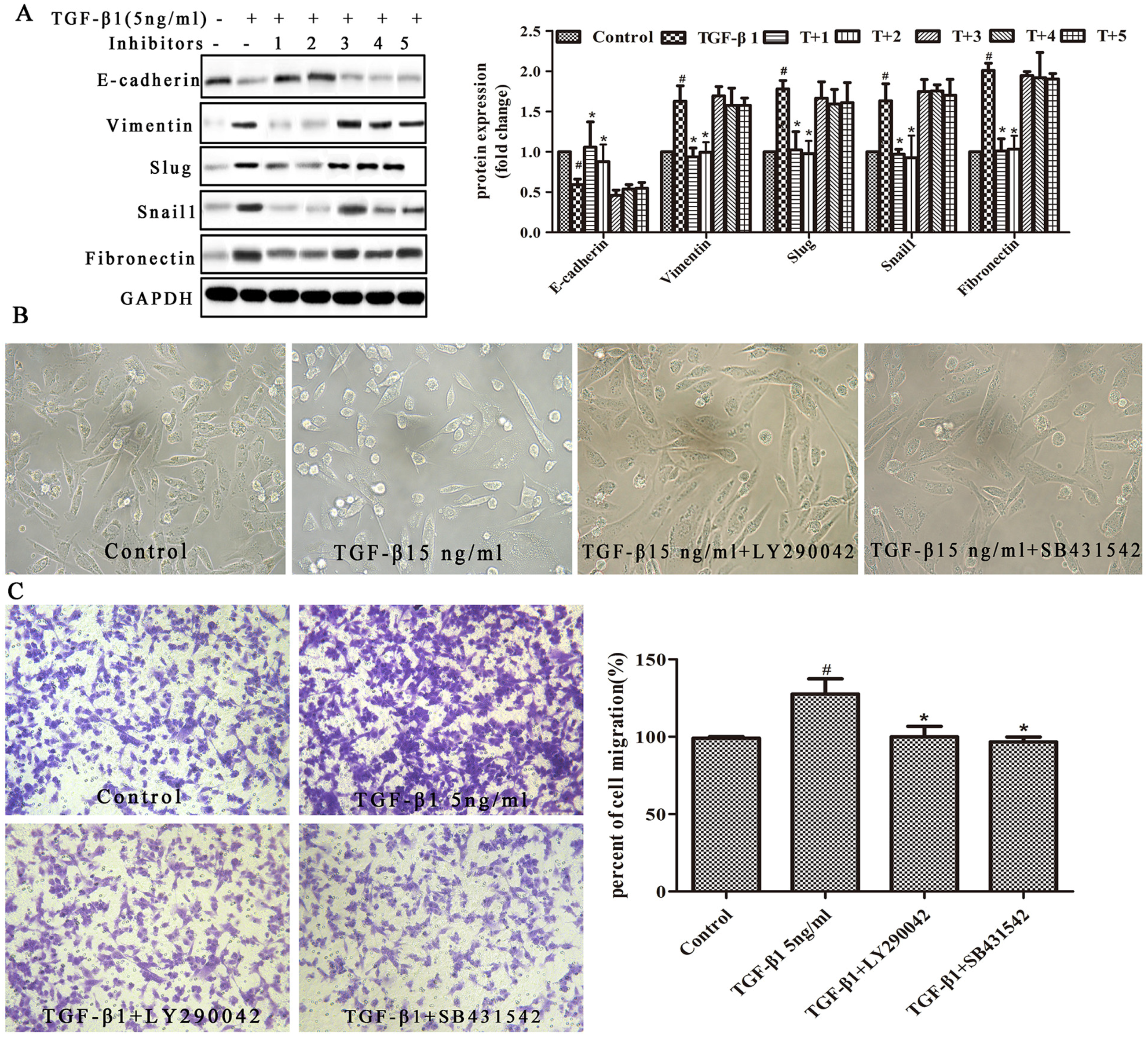
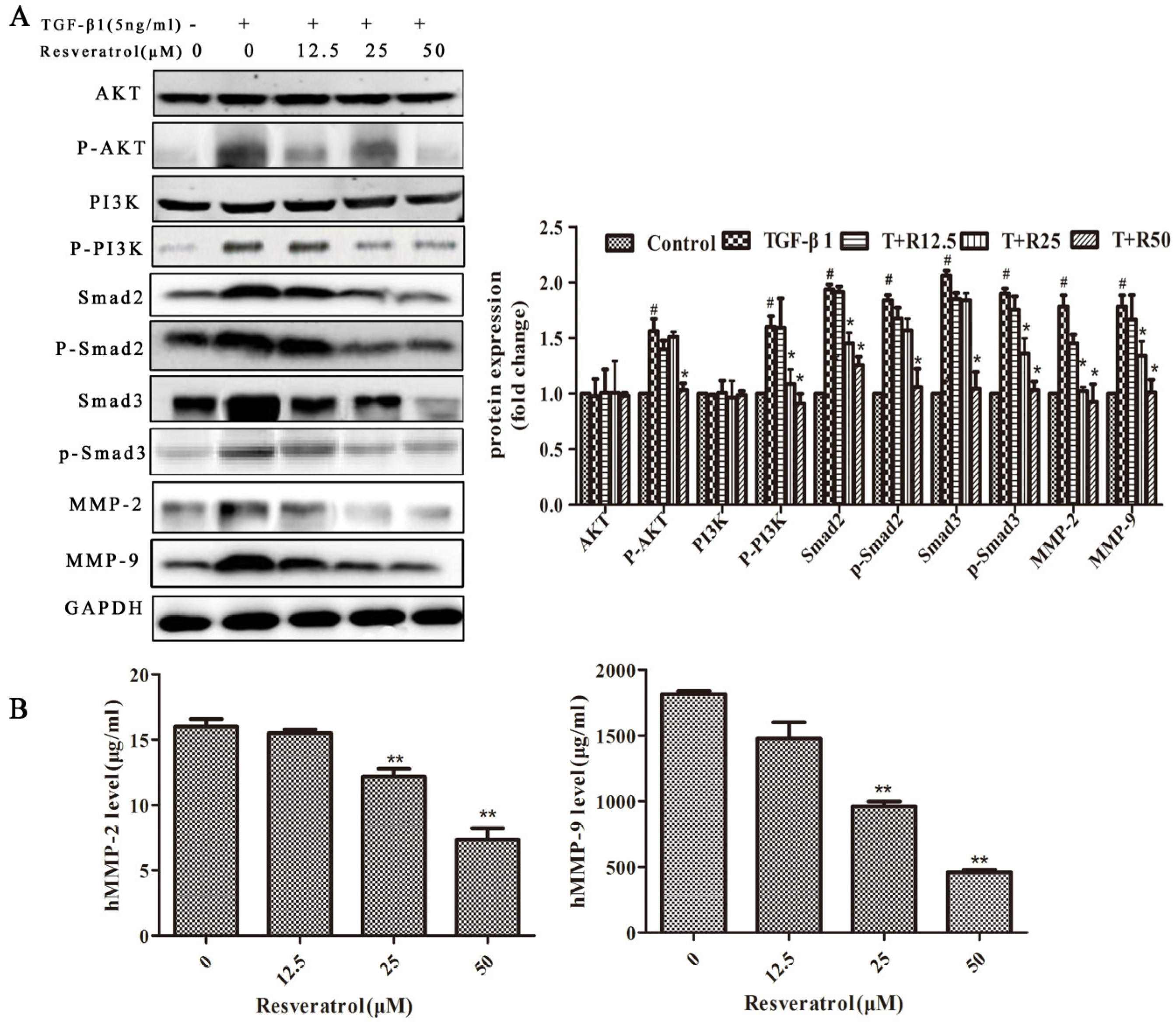
2.3. PI3K/AKT and Smad Signaling Mediates EMT in TGF-β1-Induced MDA231 Cells
2.4. Resveratrol Reverses TGF-β1-Induced EMT Through the PI3K/AKT, Smad, and MMP Signaling Pathways
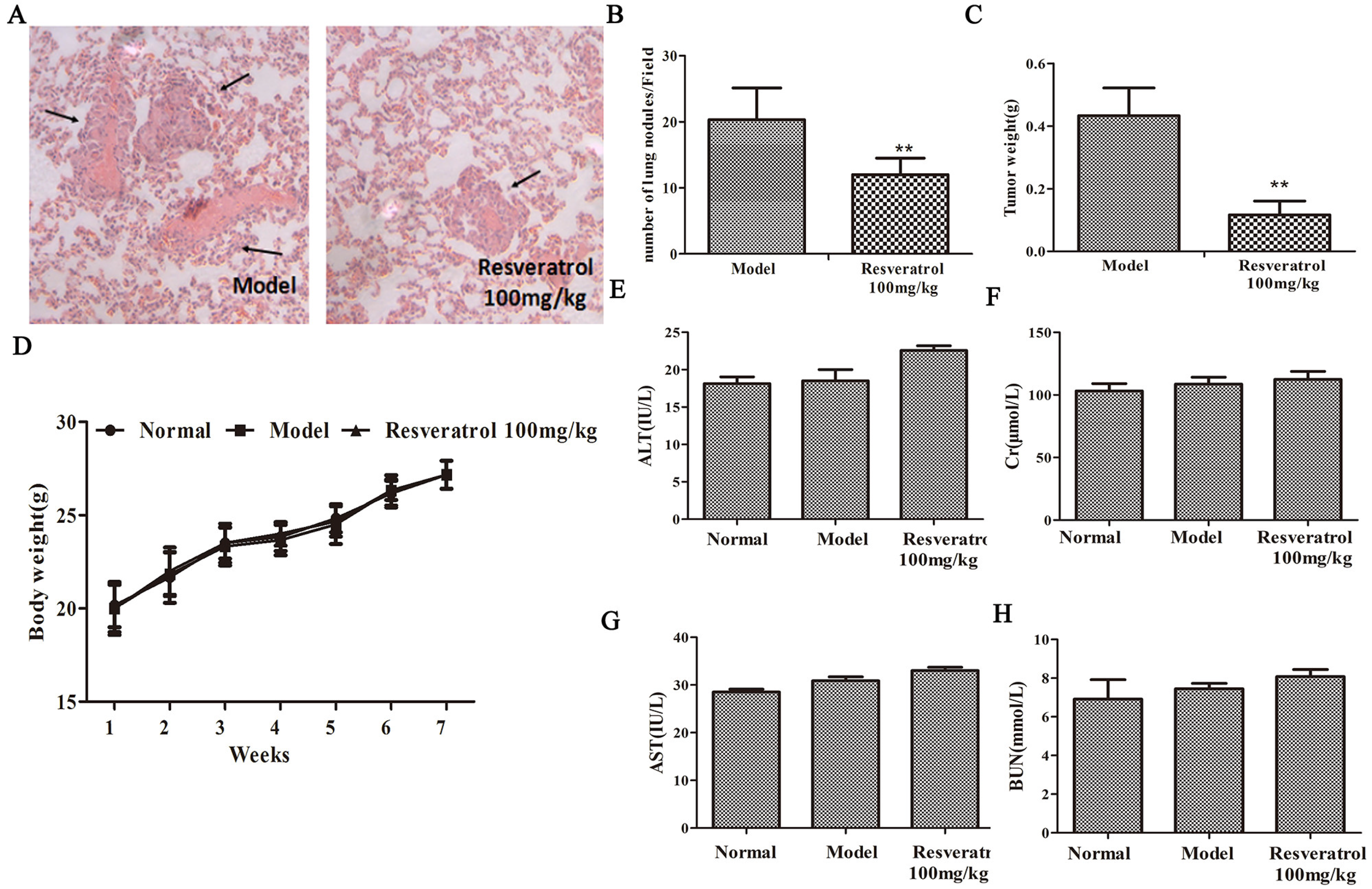
2.5. Effects of Resveratrol on MDA231 Xenografts
2.6. Resveratrol Does Not Cause Side Effects in Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Migration Assays
4.5. Effects of Resveratrol on Breast Cancer MDA231 Xenografts
4.6. Liver and Kidney Function Tests
4.7. ELISA
4.8. Immunofluorescence
4.9. Western Blot Analyses
4.10. Statistical Analyses
4.11. Ethics Approval and Consent to Participate
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TGF | transforming growth factor |
| EMT | epithelial-mesenchymal transition |
| ELISA | enzyme-linked immunosorbent assay |
| MMP-2 | metalloproteinase-2 |
| MMP-9 | metalloproteinase-9 |
| EGF | epidermal growth factor |
| VEGF | vascular endothelial growth factor |
| PDGF | platelet-derived growth factor |
| HGF | human growth factor |
| DMSO | dimethyl sulfoxide |
| MTS | 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium |
| H&E | haematoxylin and eosin |
| ALT | alanine aminotransferase |
| AST | aspartic transaminase |
| Cr | creatinine |
| BUN | urea nitrogen |
| JNK | JunN-terminalkinase |
| PDTC | pyrrolidinedithiocarbamate ammonium |
| PI3K | phosphatidylinositide-3 kinase |
| TNBC | triple-negative breast cancer |
| ECM | extracellular matrix |
| DAPI | 4′,6-diamidino-2-phenylindole |
| MDA231 | MDA-MB-231 |
| MD453 | MDA-MB-453 |
| MDA436 | MDA-MB-436 |
| BT549 | BT-549 |
References
- Coughlin, S.S.; Ekwueme, D.U. Breast cancer as a global health concern. Cancer Epidemiol. 2009, 33, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Mouh, F.Z.; Mzibri, M.E.; Slaoui, M.; Amrani, M. Recent progress in triple negative breast cancer research. Asian Pac. J. Cancer Prev. 2016, 17, 1595–1608. [Google Scholar] [CrossRef]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef]
- Singh, M.; Yelle, N.; Venugopal, C.; Singh, S.K. EMT: Mechanisms and therapeutic implications. Pharmacol. Ther. 2018, 182, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Guarino, M. Epithelial-mesenchymal transition and tumour invasion. Int. J. Biochem. Cell 2007, 39, 2153–2160. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, Q.; Zhou, Y.; Zheng, L.; Chiu, J.F.; He, Q.Y. Epidermal growth factor-induced epithelial-mesenchymal transition in human esophageal carcinoma cells-a model for the study of metastasis. Cancer Lett. 2010, 296, 88–95. [Google Scholar] [CrossRef]
- Ahmad, A.; Biersack, B.; Li, Y.; Kong, D.; Bao, B.; Schobert, R.; Padhye, S.B.; Sarkar, F. Targeted regulation of PI3K/Akt/mTOR/NF-κB signaling by indole compounds and their derivatives: Mechanistic details and biological implications for cancer therapy. Anticancer. Agents Med. Chem. 2013, 13, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Moreno, B.G.; Peinado, H.; Molina, P.; Olmeda, D.; Cubillo, E.; Santos, V.; Palacios, J.; Portillo, F.; Cano, A. The morphological and molecular features of the epithelial-to-mesenchymal transition. Nat. Protoc. 2009, 4, 1591–1613. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, X.J.; Xing, J. Signal Transduction pathways of EMT induced by TGF-β1, SHH, and WNT and their Crosstalks. J. Clin. Med. 2016, 5, 41. [Google Scholar] [CrossRef]
- Park, N.R.; Cha, J.H.; Jang, J.W.; Bae, S.H.; Jang, B.; Kim, J.H.; Hur, W.; Choi, J.Y.; Yoon, S.K. Synergistic effects of CD44 and TGF-β1 through AKT/GSK-3β/β-catenin signaling during epithelial-mesenchymal transition in liver cancer cells. Biochem. Biophys. Res. Commun. 2016, 477, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, X.; Hao, F.; Dong, A.; Chen, D. Targeting TGF-β1 inhibits invasion of anaplastic thyroid carcinoma cell through SMAD2-dependent S100A4-MMP-2/9 signalling. Am. J. Transl. Res. 2016, 8, 2196–2209. [Google Scholar]
- Huang, X.; Zhu, H.L. Resveratrol and its analogues: Promising antitumor agents. Agents Med. Chem. 2011, 11, 479–490. [Google Scholar]
- Rauf, A.; Imran, M.; Suleria, H.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zheng, Y.; Li, T.W.; Peng, H.; Fernandez, R.D.; Martínez-Chantar, M.L.; Rojas, A.L.; Mato, J.M.; Lu, S.C. Methionine adenosyltransferase 2B, HuR, and sirtuin 1 protein cross-talk impacts on the effect of resveratrol on apoptosis and growth in liver cancer cells. J. Biol. Chem. 2013, 288, 23161–23170. [Google Scholar] [CrossRef]
- Yousef, M.; Vlachogiannis, I.A.; Tsiani, E. Effects of resveratrol against lung cancer: In vitro and in vivo studies. Nutrients 2017, 9, 1231. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Zhang, H.J.; Tang, L.; Chen, H.X.; Wu, C.L.; Zhao, M.C.; Yao, Q.Y.; Chen, X.F.; Liu, G.T. Resveratrol inhibits TGF-β1-induced epithelial-to-mesenchymal transition and Suppresses lung cancer invasion and metastasis. Toxicology 2013, 303, 139–146. [Google Scholar] [CrossRef]
- Sinha, D.; Sarkar, N.; Biswas, J.; Bishayee, A. Resveratrol for breast cancer prevention and therapy: Preclinical evidence and molecular mechanisms. Semin. Cancer Biol. 2016, 40–41, 209–232. [Google Scholar] [CrossRef]
- Jie, H.T.; Li, S.H.; Chih, L.L.; Hui, M.H.; Min, H.P.; Tzong, D.W.; Wei, J.C. 3,5,4-Trimethoxystilbene, a natural methoxylated analog of resveratrol, inhibits breast cancer cell invasiveness by down regulation of PI3K/Akt and Wnt/β-catenin signaling cascades and reversal of epithelial–mesenchymal transition. Toxicol. Appl. Pharmacol. 2013, 272, 746–756. [Google Scholar]
- Sengupta, S.; Jana, S.; Biswas, S.; Mandal, P.K.; Bhattacharyya, A. Cooperative involvement of NFAT and SnoN mediates transforming growth factor-β (TGF-β) induced EMT in metastatic breast cancer (MDA-MB 231) cells. Clin. Exp. Metastasis. 2013, 30, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, M.K.; Choi, K.S.; Jeong, J.W.; Bae, S.K.; Kim, H.J.; Bae, M.K. Neuromedin B receptor antagonism inhibits migration, invasion, and epithelial-mesenchymal transition of breast cancer cells. Int. J. Oncol. 2016, 49, 334–342. [Google Scholar] [CrossRef]
- Shao, S.; Zhao, X.; Zhang, X.; Luo, M.; Zuo, X.; Huang, S.; Wang, Y.; Gu, S.; Zhao, X. Notch1 signaling regulates the epithelial-mesenchymal transition and invasion of breast cancer in a Slug-dependent manner. Mol. Cancer 2015, 14, 28. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar] [CrossRef]
- Jeandet, P.; Breuil, A.C.; Adrian, M.; Weston, L.A.; Debord, S.; Meunier, P.; Maume, G.; Bessis, R. HPLC analysis of grapevine phytoalexins coupling photodiode array detection and fluorometry. Anal. Chem. 1997, 69, 5172–5177. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Sobarzo-Sánchez, E.; Clément, C.; Nabavi, S.F.; Habtemariam, S.; Nabavi, S.M.; Cordelier, S. Engineering stilbene metabolic pathways in microbial cells. Biotechnol. Adv. 2018, 36, 2264–228323. [Google Scholar] [CrossRef]
- Fulda, S. Regulation of cell death and survival by resveratrol: Implications for cancer therapy. Anticancer Agents Med. Chem. 2012, 12, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Fasano, E.; Serini, S.; Mondella, N.; Trombino, S.; Celleno, L.; Lanza, P.; Cittadini, A.; Calviello, G. Antioxidant and anti-inflammatory effects of selected natural compounds contained in a dietary supplement on two human immortalized keratinocyte lines. Biomed. Res. Int. 2014, 327452. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Yi, L.; Jin, X.; Xie, Q.; Zhang, T.; Zhou, X.; Chang, H.; Fu, Y.J.; Zhu, J.D.; Zhang, Q.Y.; et al. Absorption of resveratrol by vascular endothelial cells through passive diffusion and an SGLT1-mediated pathway. J. Nutr. Biochem. 2013, 24, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Pichardo, L.; Cubano, L.A.; Dharmawardhane, S. Dietary grape polyphenol resveratrol increases mammary tumor growth and metastasis in immunocompromised mice. MC Complement. Altern. Med. 2013, 13, 6. [Google Scholar] [CrossRef]
- Bill, R.; Christofori, G. The relevance of EMT in breast cancer metastasis: Correlation or causality? FEBS Lett. 2015, 589, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.; Wu, G. Molecular mechanisms involved in podocyte EMT and concomitant diabetic kidney diseases: An update. Ren. Fail. 2017, 39, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Lombaerts, M.; Wezel, T.; Philippo, K.; Dierssen, J.W.F.; Zimmerman, R.M.E.; Oosting, J.; Van Eijk, R.; Eilers, P.H.; Water, B.; Cornelisse, C.J.; et al. E-cadherin transcriptional downregulation by promoter methylation but not mutation is related to epithelial-tomesenchymal transition in breast cancer cell lines. Br. J. Cancer. 2006, 94, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Jeremy, R.; James, G.; Rena, G.; Hemi, C.; Rosa, X.; David, F.; William, B.; Paula, M.; Nancy, K.; Stephen, B.; et al. E-cadherin expression is silenced by DNA hypermethylation in human breast and prostate carcinomas. Cancer Res. 1995, 55, 5195–5199. [Google Scholar]
- Shi, X.; Miao, S.; Wu, Y.; Zhang, W.; Zhang, X.; Ma, H.; Xin, H.; Feng, J.; Wen, A.; Li, Y. Resveratrol sensitizes tamoxifen in antiestrogen-resistant breast cancer cells with epithelial-mesenchymal transition features. Int. J. Mol. Sci. 2013, 14, 15655–15668. [Google Scholar] [CrossRef]
- Shay, G.; Lynch, C.C.; Fingleton, B. Moving targets: Emerging roles for MMPs in cancer progression and metastasis. Matrix Biol. 2015, 6, 200–206. [Google Scholar] [CrossRef]
- Yao, Z.; Yuan, T.; Wang, H.; Yao, S.; Zhao, Y.; Liu, Y.; Jin, S.; Chu, J.; Xu, Y.; Zhou, W.; et al. MMP-2 together with MMP-9 overexpression correlated with lymph node metastasis and poor prognosis in early gastric carcinoma. Tumour Biol. 2017, 39, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Costa, R.L.B.; Han, H.S.; Gradishar, W.J. Targeting the PI3K/AKT/mTOR pathway in triple-negative breast cancer: A review. Breast Cancer Res. Treat. 2018, 169, 397–406. [Google Scholar] [CrossRef]
- Bakin, A.V.; Tomlinson, A.K.; Bhowmick, N.A.; Moses, H.L.; Arteaga, C.L. Phosphatidylinositol 3-kinase function is required for transforming growth factor beta-mediated epithelial to mesenchymal transition and cell migration. J. Biol. Chem. 2000, 275, 36803–36810. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Kang, Y. Multilayer control of the EMT master regulators. Oncogene 2014, 33, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Attisano, L. The TGF-β superfamily signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Valcourt, U.; Kowanetz, M.; Niimi, H.; Heldin, C.H.; Moustakas, A. TGF-beta and the Smad signaling pathway support transcriptomic reprogramming during epithelial-mesenchymal cell transition. Mol. Biol. Cell. 2005, 16, 1987–2002. [Google Scholar] [CrossRef] [PubMed]
- Dzwonek, J.; Preobrazhenska, O.; Cazzola, S.; Conidi, A.; Schellens, A.; Dinther, M.; Stubbs, A.; Klippel, A.; Huylebroeck, D.; Dijke, P.; et al. Smad3 is a key nonredundant mediator of transforming growth factor β signaling in Nme mouse mammary epithelial cells. Mol. Cancer Res. 2009, 7, 1342–1353. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: All data in this study can be obtained from the corresponding author upon reasonable request. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zhou, Q.-M.; Lu, Y.-Y.; Zhang, H.; Chen, Q.-L.; Zhao, M.; Su, S.-B. RETRACTED: Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition. Molecules 2019, 24, 1131. https://doi.org/10.3390/molecules24061131
Sun Y, Zhou Q-M, Lu Y-Y, Zhang H, Chen Q-L, Zhao M, Su S-B. RETRACTED: Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition. Molecules. 2019; 24(6):1131. https://doi.org/10.3390/molecules24061131
Chicago/Turabian StyleSun, Yang, Qian-Mei Zhou, Yi-Yu Lu, Hui Zhang, Qi-Long Chen, Ming Zhao, and Shi-Bing Su. 2019. "RETRACTED: Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition" Molecules 24, no. 6: 1131. https://doi.org/10.3390/molecules24061131
APA StyleSun, Y., Zhou, Q.-M., Lu, Y.-Y., Zhang, H., Chen, Q.-L., Zhao, M., & Su, S.-B. (2019). RETRACTED: Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition. Molecules, 24(6), 1131. https://doi.org/10.3390/molecules24061131