Genotype-Related Differences in the Phenolic Compound Profile and Antioxidant Activity of Extracts from Olive (Olea europaea L.) Leaves

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction Yield and Total Phenolic Content
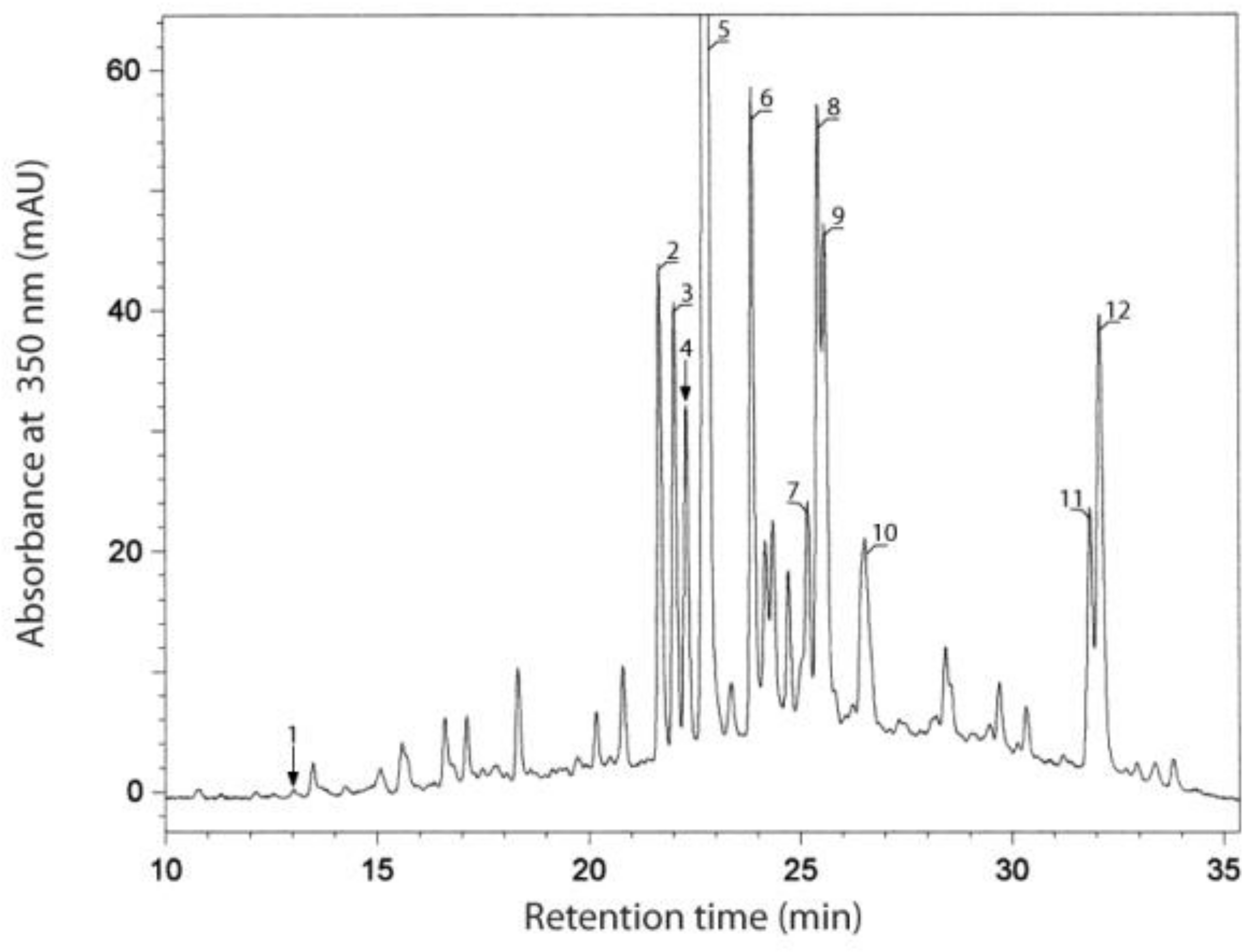
2.2. Identification and Quantification of Phenolic Compounds
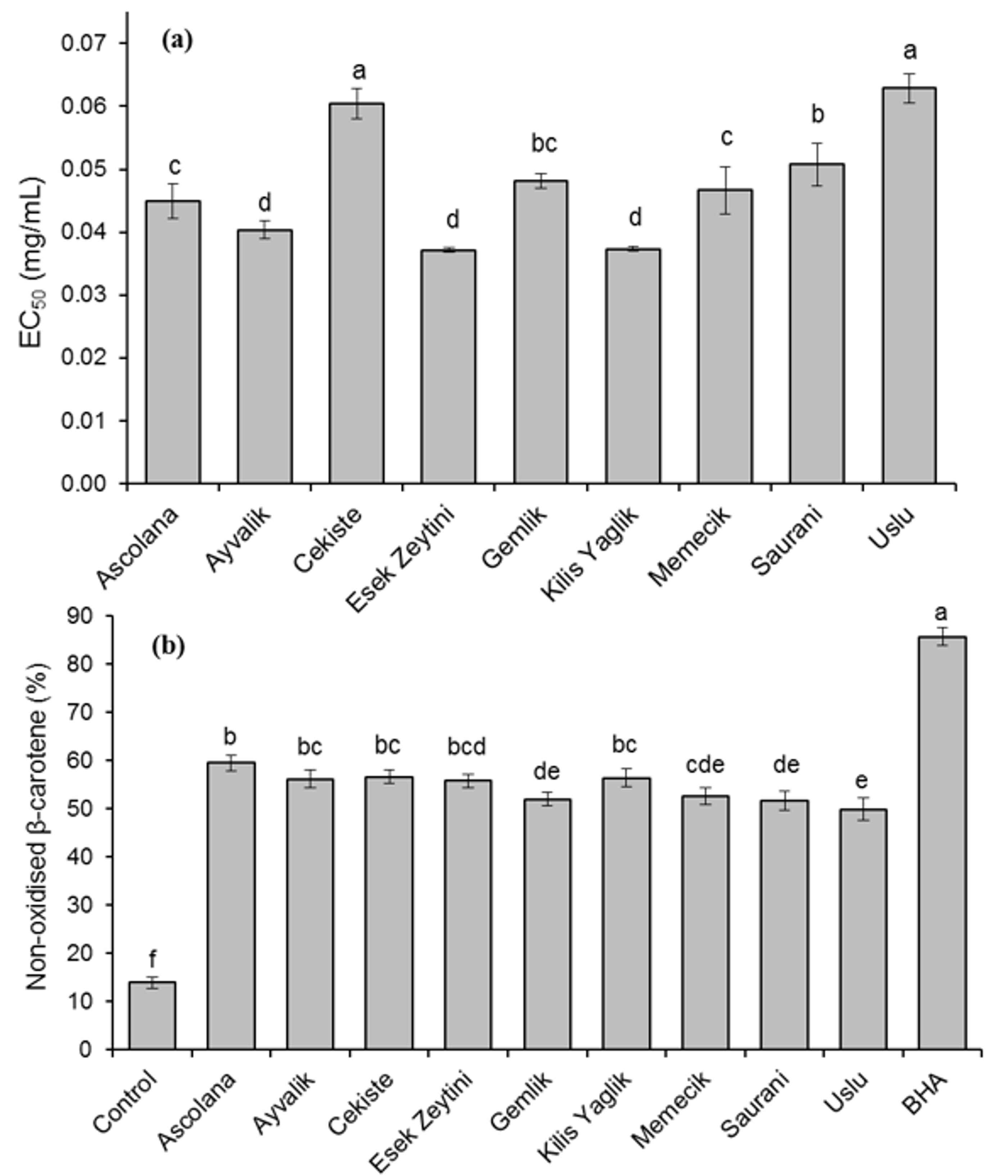
2.3. Antioxidant Activity of Olive Leaf Extracts
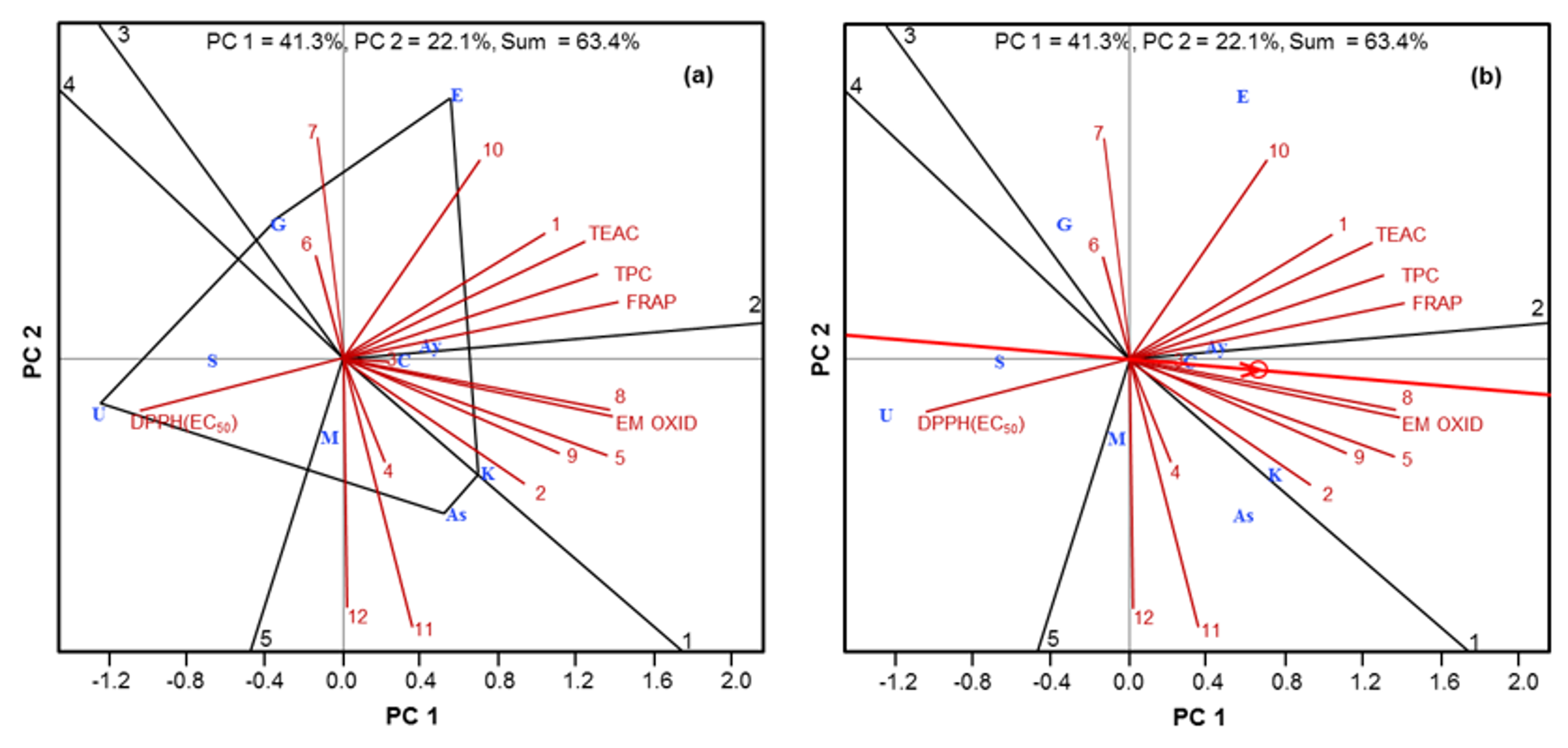
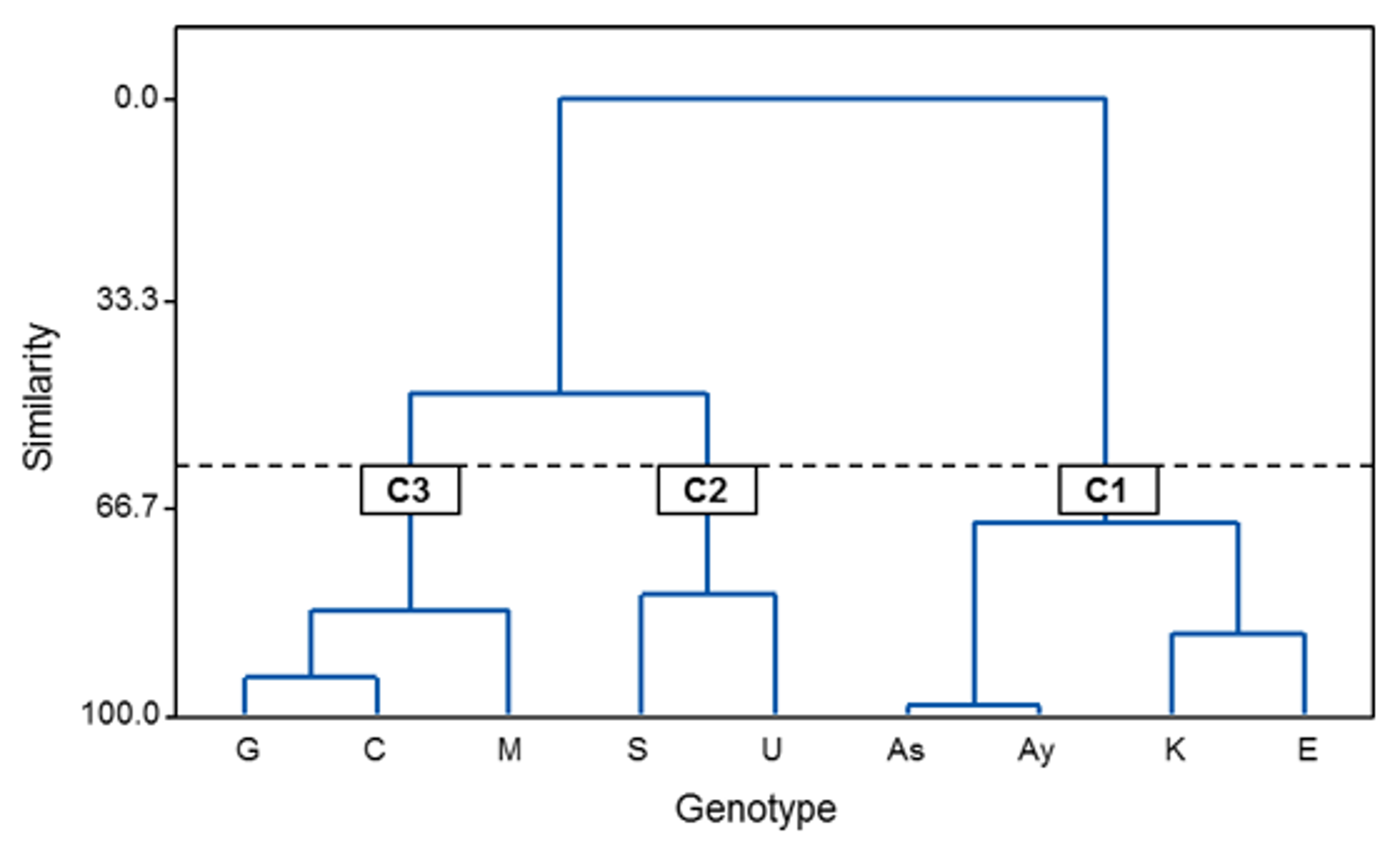
2.4. Chemometric Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals and Reagents
3.3. Extract Preparation
3.4. Determination of Total Phenolic Compound Content
3.5. Phenolic Compounds Analysis
3.6. Determination of Trolox Equivalent Antioxidant Capacity
3.7. Determination of DPPH Radical Scavenging Activity
3.8. Determination of Ferric-Reducing Antioxidant Power
3.9. β-Carotene-Linoleic Acid Emulsion Oxidation
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Romero-García, J.M.; Niño, L.; Martínez-Patiño, C.; Álvarez, C.; Castro, E.; Negro, M.J. Biorefinery based on olive biomass. State of the art and future trends. Bioresour. Technol. 2014, 159, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.A.; Pimentel, F.B.; Costa, A.S.G.; Alves, R.C.; Oliveira, M.B.P.P. Olive by-products for functional and food applications: Challenging opportunities to face environmental constraints. Innov. Food Sci. Emerg. Technol. 2016, 35, 139–148. [Google Scholar] [CrossRef]
- Spinelli, R.; Picchi, G. Industrial harvesting of olive tree pruning residue for energy biomass. Bioresour. Technol. 2010, 101, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.; Pimentel, F.B.; Oliveira, M.B.P.P. Olive by-products: Challenge application in cosmetic industry. Ind. Crops Prod. 2015, 70, 116–124. [Google Scholar] [CrossRef]
- Souilem, S.; Fki, I.; Kobayashi, I.; Khalid, N.; Neves, M.; Isoda, H.; Sayadi, S.; Nakajima, M. Emerging technologies for recovery of value-added components from olive leaves and their applications in food/feed industries. Food Bioprocess Technol. 2017, 10, 229–248. [Google Scholar] [CrossRef]
- Talhaoui, N.; Taamalli, A.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Phenolic compounds in olive leaves: Analytical determination, biotic and abiotic influence, and health benefits. Food Res. Int. 2015, 77, 92–108. [Google Scholar] [CrossRef]
- Ryan, D.; Antolovich, M.; Prenzler, P.; Robards, K.; Lavee, S. Biotransformations of phenolic compounds in Olea europaea L. Sci. Hortic. 2002, 92, 147–176. [Google Scholar] [CrossRef]
- Brahmi, F.; Mechri, B.; Dhibi, M.; Hammami, M. Variations in phenolic compounds and antiradical scavenging activity of Olea europaea leaves and fruits extracts collected in two different seasons. Ind. Crops Prod. 2013, 49, 256–264. [Google Scholar] [CrossRef]
- Benavente-García, O.; Castillo, J.; Lorente, J.; Ortuño, A.; Del Rio, J.A. Antioxidant activity of phenolics extracted from Olea europaea L. leaves. Food Chem. 2000, 68, 457–462. [Google Scholar] [CrossRef]
- Lee, O.H.; Lee, B.Y. Antioxidant and antimicrobial activities of individual and combined phenolics in Olea europaea leaf extract. Bioresour. Technol. 2010, 101, 3751–3754. [Google Scholar] [CrossRef]
- Di Donna, L.; Mazzotti, F.; Naccarato, A.; Salerno, R.; Tagarelli, A.; Taverna, D.; Sindona, G. Secondary metabolites of Olea europaea leaves as markers for the discrimination of cultivars and cultivation zones by multivariate analysis. Food Chem. 2010, 121, 492–496. [Google Scholar] [CrossRef]
- Ortega-García, F.; Peragón, J. Phenol metabolism in the leaves of the olive tree (Olea europaea L.) cv. Picual, Verdial, Arbequina, and Frantoio during ripening. J. Agric. Food Chem. 2010, 58, 12440–12448. [Google Scholar] [CrossRef]
- Orak, H.H.; Isbilir, S.S.; Yagar, H. Determination of antioxidant properties of lyophilized olive leaf water extracts obtained from 21 different cultivars. Food Sci. Biotechnol. 2012, 21, 1065–1074. [Google Scholar] [CrossRef]
- Talhaoui, N.; Gómez-Caravaca, A.M.; León, L.; De la Rosa, R.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Determination of phenolic compounds of ’Sikitita’ olive leaves by HPLC-DAD-TOF-MS. Comparison with its parents ’Arbequina’ and ’Picual’ olive leaves. LWT Food Sci. Technol. 2014, 58, 28–34. [Google Scholar] [CrossRef]
- Michel, T.; Khlif, I.; Kanakis, P.; Termentzi, A.; Allouche, N.; Halabalaki, M.; Skaltsounis, A.L. UHPLC-DAD-FLD and UHPLC-HRMS/MS based metabolic profiling and characterization of different Olea europaea organs of Koroneiki and Chetoui varieties. Phytochem. Lett. 2015, 11, 424–439. [Google Scholar] [CrossRef]
- Talhaoui, N.; Gómez-Caravaca, A.M.; Roldán, C.; León, L.; De la Rosa, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Chemometric analysis for the evaluation of phenolic patterns in olive leaves from six cultivars at different growth stages. J. Agric. Food Chem. 2015, 63, 1722–1729. [Google Scholar] [CrossRef]
- Herrero, M.; Temirzoda, T.N.; Segura-Carretero, A.; Quirantes, R.; Plaza, M.; Ibañez, E. New possibilities for the valorization of olive oil by-products. J. Chromatogr. A 2011, 1218, 7511–7520. [Google Scholar] [CrossRef]
- Stanković, M.; Ćurčić, S.; Zlatić, N.; Bojović, B. Ecological variability of the phenolic compounds of Olea europaea L. leaves from natural habitats and cultivated conditions. Biotechnol. Biotechnol. Equip. 2017, 31, 499–504. [Google Scholar] [CrossRef]
- Goldsmith, C.D.; Vuong, Q.V.; Sadeqzadeh, E.; Stathopoulos, C.E.; Roach, P.D.; Scarlett, C.J. Phytochemical properties and anti-proliferative activity of Olea europaea L. leaf extracts against pancreatic cancer cells. Molecules 2015, 20, 12992–13004. [Google Scholar] [CrossRef]
- Blasi, F.; Urbani, E.; Simonetti, M.S.; Chiesi, C.; Cossignani, L. Seasonal variations in antioxidant compounds of Olea europaea leaves collected from different Italian cultivars. J. Appl. Bot. Food Qual. 2016, 89, 202–207. [Google Scholar]
- Goulas, V.; Papoti, V.T.; Exarchou, V.; Tsimidou, M.Z.; Gerothanassis, I.P. Contribution of flavonoids to the overall radical scavenging activity of olive (Olea europaea L.) leaf polar extracts. J. Agric. Food Chem. 2010, 58, 3303–3308. [Google Scholar] [CrossRef] [PubMed]
- Briante, R.; Patumi, M.; Terenziani, S.; Bismuto, E.; Febbraio, F.; Nucci, R. Olea europaea L. leaf extract and derivatives: Antioxidant properties. J. Agric. Food Chem. 2002, 50, 4934–4940. [Google Scholar] [CrossRef]
- Silva, S.; Gomes, L.; Leitão, F.; Coelho, A.V.; Boas, L.V. Phenolic compounds and antioxidant activity of Olea europaea L. fruits and leaves. Int. J. Food Sci. Technol. 2006, 12, 385–396. [Google Scholar] [CrossRef]
- Tóth, G.; Alberti, Á.; Sólyomváry, A.; Barabás, C.; Boldizsár, I.; Noszála, B. Phenolic profiling of various olive bark-types and leaves: HPLC–ESI/MS study. Ind. Crops Prod. 2015, 67, 432–438. [Google Scholar] [CrossRef]
- Taamalli, A.; Arráez-Román, D.; Ibañez, E.; Zarrouk, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Optimization of microwave-assisted extraction for the characterization of olive leaf phenolic compounds by using HPLC-ESI-TOF-MS/IT-MS2. J. Agric. Food Chem. 2012, 60, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Janiak, M.A.; Slavova-Kazakova, A.; Kancheva, V.D.; Ivanova, M.; Tsrunchev, T.; Karamać, M. Effects of γ-irradiation of wild thyme (Thymus serpyllum L.) on the phenolic compounds profile of its ethanolic extract. Pol. J. Food Nutr. Sci. 2017; 67, 309–315. [Google Scholar]
- Pereira, A.P.; Ferreira, I.C.F.R.; Marcelino, F.; Valentão, P.; Andrade, P.B.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J.A. Phenolic compounds and antimicrobial activity of olive (Olea europaea L. cv. Cobrançosa) leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Petridis, A.; Therios, I.; Samouris, G. Genotypic variation of total phenol and oleuropein concentration and antioxidant activity of 11 Greek olive cultivars (Olea europaea L.). Hortscience 2012, 47, 339–342. [Google Scholar] [CrossRef]
- Orak, H.H.; Karamać, M.; Orak, A.; Amarowicz, R. Antioxidant potential and phenolic compounds of some widely consumed Turkish white bean (Phaseolus vulgaris L.) varieties. Pol. J. Food Nutr. Sci. 2016, 66, 253–260. [Google Scholar] [CrossRef]
- Karamać, M.; Orak, H.H.; Amarowicz, R.; Orak, A.; Piekoszewski, W. Phenolic contents and antioxidant capacities of wild and cultivated white lupin (Lupinus albus L.) seeds. Food Chem. 2018, 258, 1–7. [Google Scholar] [CrossRef]
- Yan, W.; Rajcan, I. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–655. [Google Scholar] [CrossRef]
- Hossain, M.B.; Patras, A.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N.P. Application of principal component and hierarchical cluster analysis to classify different spices based on in vitro antioxidant activity and individual polyphenolic antioxidant compounds. J. Funct. Food. 2011, 3, 179–189. [Google Scholar] [CrossRef]
- Wang, L.J.; Wu, J.; Wang, H.X.; Li, S.S.; Zheng, X.C.; Du, H.; Xu, Y.J.; Wang, L.S. Composition of phenolic compounds and antioxidant activity in the leaves of blueberry cultivars. J. Funct. Food. 2015, 16, 295–304. [Google Scholar] [CrossRef]
- Amarowicz, R.; Karamać, M.; Weidner, S.; Abe, S.; Shahidi, F. Antioxidant activity of wheat caryopses and embryos extracts. J. Food Lipids 2002, 9, 201–210. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Miller, H.E. A simplified method for the evaluation of antioxidants. J. Am. Oil Chem. Soc. 1971, 48, 91. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Extract Yield (%) | TPC mg GAE/g) | TEAC (mmol TE/g) | FRAP (mmol Fe2+/g) |
|---|---|---|---|---|
| Ascolona | 29.58 | 236 ± 4.8 b | 0.83 ± 0.03 e | 1.79 ± 0.037 c |
| Ayvalik | 29.87 | 242 ± 3.2 b | 0.98 ± 0.08 b | 1.78 ± 0.027 c |
| Cekiste | 25.84 | 206 ± 0.4 d | 0.96 ± 0.06 c | 1.48 ± 0.048 e |
| Esek Zeytini | 29.79 | 268 ± 3.0 a | 1.01 ± 0.04 a | 2.12 ± 0.016 a |
| Gemlik | 27.39 | 199 ± 1.2 e | 0.93 ± 0.04 d | 1.42 ± 0.024 e |
| Kilis Yaglik | 28.52 | 225 ± 8.1 c | 1.01 ± 0.08 a | 1.99 ± 0.069 b |
| Memecik | 27.11 | 209 ± 3.4 d | 0.84 ± 0.10 d | 1.60 ± 0.074 d |
| Saurani | 24.63 | 197 ± 1.7 e | 0.75 ± 0.05 f | 1.23 ± 0.052 f |
| Uslu | 24.46 | 110 ± 4.3 f | 0.70 ± 0.14 g | 1.04 ± 0.019 g |
| No | Compound | Ascolana | Ayvalik | Cekiste | Esek Zeytini | Gemlik | Kilis Yaglik | Memecik | Saurani | Uslu |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Hydroxytyrosol | 2.32 ± 0.12 c | 1.96 ± 0.10 d | 3.38 ± 0.17 b | 4.03 ± 0.20 a | 2.52 ± 0.13 c | 2.04 ± 0.10 d | 2.10 ± 0.11 d | 2.44 ± 0.12 c | 1.33 ± 0.07 e |
| 2 | Luteolin glycoside 1 * | 1.15 ± 0.06 c | 1.15 ± 0.06 c | 1.83 ± 0.09 a | 0.76 ± 0.07 d | 0.87 ± 0.04 d | 1.89 ± 0.09 a | 1.48 ± 0.04 b | 0.89 ± 0.04 d | 0.45 ± 0.02 e |
| 3 | Verbascoside | 6.28 ± 0.24 c | 0.72 ± 1.05 f | 0.45 ± 0.12 f | 4.89 ± 0.74 d | 19.55 ± 0.98 b | 21.07 ± 0.20 a | 3.47 ± 0.17 e | 3.25 ± 0.31 e | 3.92 ± 0.16 de |
| 4 | Luteolin glycoside 2 * | 0.84 ± 0.04 a | 0.47 ± 0.02 c | 0.37 ± 0.02 d | 0.22 ± 0.01 ef | 0.63 ± 0.03 b | 0.51 ± 0.03 c | 0.20 ± 0.01 f | 0.67 ± 0.03 b | 0.25 ± 0.01 e |
| 5 | Luteolin 7-O-glucoside | 8.11 ± 0.41 a | 7.43 ± 0.37 b | 6.84 ± 0.34 b | 5.69 ± 0.28 c | 5.23 ± 0.26 cd | 8.67 ± 0.43 a | 4.96 ± 0.25 de | 4.50 ± 0.23 e | 3.23 ± 0.16 f |
| 6 | Apigenin glycoside ** | 0.57 ± 0.03 ef | 0.64 ± 0.03 d | 0.88 ± 0.01 b | 0.54 ± 0.03 f | 1.48 ± 0.07 a | 0.42 ± 0.02 g | 0.77 ± 0.04 c | 0.63 ± 0.03 de | 0.38 ± 0.02 g |
| 7 | Apigenin 7-O-glucoside | 0.20 ± 0.01 d | 0.40 ± 0.02 b | 0.21 ± 0.01 d | 0.40 ± 0.02 b | 0.59 ± 0.03 a | 0.08 ± 0.04 e | 0.18 ± 0.01 d | 0.29 ± 0.01 c | 0.18 ± 0.01 e |
| 8 | Luteolin 4′-O-glucoside | 3.72 ± 0.19 a | 3.64 ± 0.18 ab | 3.57 ± 0.18 ab | 3.54 ± 0.18 ab | 2.08 ± 0.10 c | 3.90 ± 0.20 a | 3.39 ± 0.17 b | 1.58 ± 0.08 d | 1.89 ± 0.09 c |
| 9 | Luteolin glycoside 3 *** | 2.98 ± 0.15 b | 2.60 ± 0.13 c | 3.61 ± 0.18 a | 1.91 ± 0.10 e | 1.92 ± 0.10 e | 2.63 ± 0.13 c | 2.28 ± 0.11 d | 1.41 ± 0.07 f | 1.27 ± 0.06 f |
| 10 | Oleuropein | 57.6 ± 2.9 b | 46.3 ± 2.3 c | 38.5 ± 1.9 d | 98.0 ± 4.9 a | 23.1 ± 1.2 e | 44.7 ± 2.2 c | 38.2 ± 1.9 d | 21.0 ± 1.0 e | 22.2 ± 1.1 e |
| 11 | Luteolin glycoside 4 * | 0.69 ± 0.03 a | 0.54 ± 0.03 cd | 0.51 ± 0.02 de | 0.31 ± 0.02 f | 0.50 ± 0.02 de | 0.61 ± 0.03 b | 0.59 ± 0.03 bc | 0.49 ± 0.03 de | 0.47 ± 0.02 e |
| 12 | Quercetin | 1.92 ± 0.10 a | 1.07 ± 0.05 c | 0.63 ± 0.03 d | 0.43 ± 0.02 e | 0.47 ± 0.02 e | 1.36 ± 0.07 b | 1.86 ± 0.09 a | 1.24 ± 0.06 bc | 1.22 ± 0.06 bc |
| TPC | TEAC | FRAP | DPPH (EC50) | Emulsion Oxidation a | |
|---|---|---|---|---|---|
| Hydroxytyrosol | 0.614 | 0.555 | 0.461 | −0.225 | 0.369 |
| Luteolin glycoside 1 | 0.359 | 0.518 | 0.395 | −0.190 | 0.525 |
| Verbascoside | 0.043 | 0.303 | 0.218 | −0.363 | −0.018 |
| Luteolin glycoside 2 | 0.179 | 0.107 | −0.014 | −0.148 | 0.352 |
| Luteolin 7-O-glucoside | 0.669 * | 0.666 * | 0.728 * | −0.570 | 0.903 ** |
| Apigenin glycoside | 0.040 | 0.201 | −0.204 | 0.117 | −0.186 |
| Apigenin 7-O-glucoside | 0.227 | 0.252 | −0.014 | −0.171 | −0.186 |
| Luteolin 4′-O-glucoside | 0.689 * | 0.699 * | 0.833 ** | −0.544 | 0.846 ** |
| Luteolin glycoside 3 | 0.464 | 0.552 | 0.436 | −0.095 | 0.806 ** |
| Oleuropein | 0.744 * | 0.664 * | 0.836 ** | −0.674 * | 0.608 |
| Luteolin glycoside 4 | −0.004 | −0.149 | 0.024 | −0.049 | 0.380 |
| Quercetin | −0.104 | −0.480 | −0.024 | −0.059 | 0.140 |
| TPC | 1 | 0.746* | 0.885 ** | −0.824 ** | 0.737 * |
| TEAC | 1 | 0.789 ** | −0.676* | 0.582 | |
| FRAP | 1 | −0.873 ** | 0.748 * | ||
| DPPH (EC50) | 1 | −0.485 |
| Cluster No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | TPC (mg GAE/g) | TEAC (mmol TE/g) | FRAP (mmol Fe2+/g) | DPPH EC50 (mg/mL) | Emulsion Oxidation (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg/g) | |||||||||||||||||
| C1 | 2.69 | 1.24 | 14.02 | 0.51 | 7.48 | 0.54 | 0.27 | 3.70 | 2.53 | 61.63 | 0.54 | 1.20 | 242.83 | 0.96 | 1.92 | 0.107 | 56.98 |
| C2 | 1.89 | 0.67 | 3.59 | 0.46 | 3.87 | 0.51 | 0.24 | 1.74 | 1.34 | 21.54 | 0.48 | 1.23 | 153.55 | 0.72 | 1.14 | 0.113 | 50.75 |
| C3 | 2.67 | 1.39 | 7.82 | 0.40 | 5.68 | 1.04 | 0.33 | 3.01 | 2.60 | 33.29 | 0.53 | 0.99 | 204.69 | 0.91 | 1.50 | 0.118 | 53.73 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orak, H.H.; Karamać, M.; Amarowicz, R.; Orak, A.; Penkacik, K. Genotype-Related Differences in the Phenolic Compound Profile and Antioxidant Activity of Extracts from Olive (Olea europaea L.) Leaves. Molecules 2019, 24, 1130. https://doi.org/10.3390/molecules24061130
Orak HH, Karamać M, Amarowicz R, Orak A, Penkacik K. Genotype-Related Differences in the Phenolic Compound Profile and Antioxidant Activity of Extracts from Olive (Olea europaea L.) Leaves. Molecules. 2019; 24(6):1130. https://doi.org/10.3390/molecules24061130
Chicago/Turabian StyleOrak, Hakime Hülya, Magdalena Karamać, Ryszard Amarowicz, Adnan Orak, and Kamila Penkacik. 2019. "Genotype-Related Differences in the Phenolic Compound Profile and Antioxidant Activity of Extracts from Olive (Olea europaea L.) Leaves" Molecules 24, no. 6: 1130. https://doi.org/10.3390/molecules24061130
APA StyleOrak, H. H., Karamać, M., Amarowicz, R., Orak, A., & Penkacik, K. (2019). Genotype-Related Differences in the Phenolic Compound Profile and Antioxidant Activity of Extracts from Olive (Olea europaea L.) Leaves. Molecules, 24(6), 1130. https://doi.org/10.3390/molecules24061130