Evidence for Effective Inhibitory Actions on Hyperpolarization-Activated Cation Current Caused by Ganoderma Triterpenoids, the Main Active Constitutents of Ganoderma Spores

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
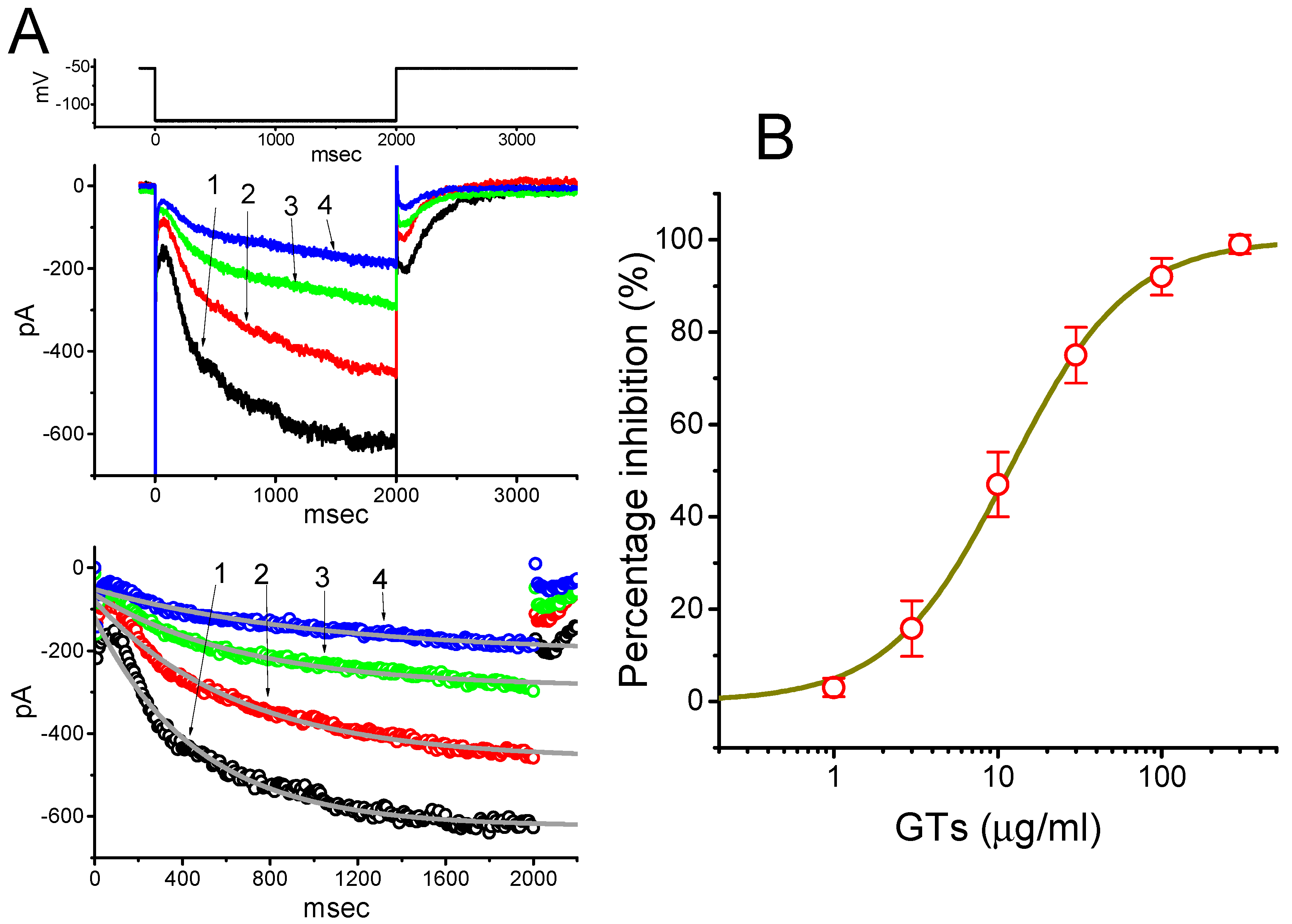
2.1. Inhibitory Effect of GTs on Hyperpolarization-Activated Cation Current (Ih) Recorded from Pituitary GH3 Cells
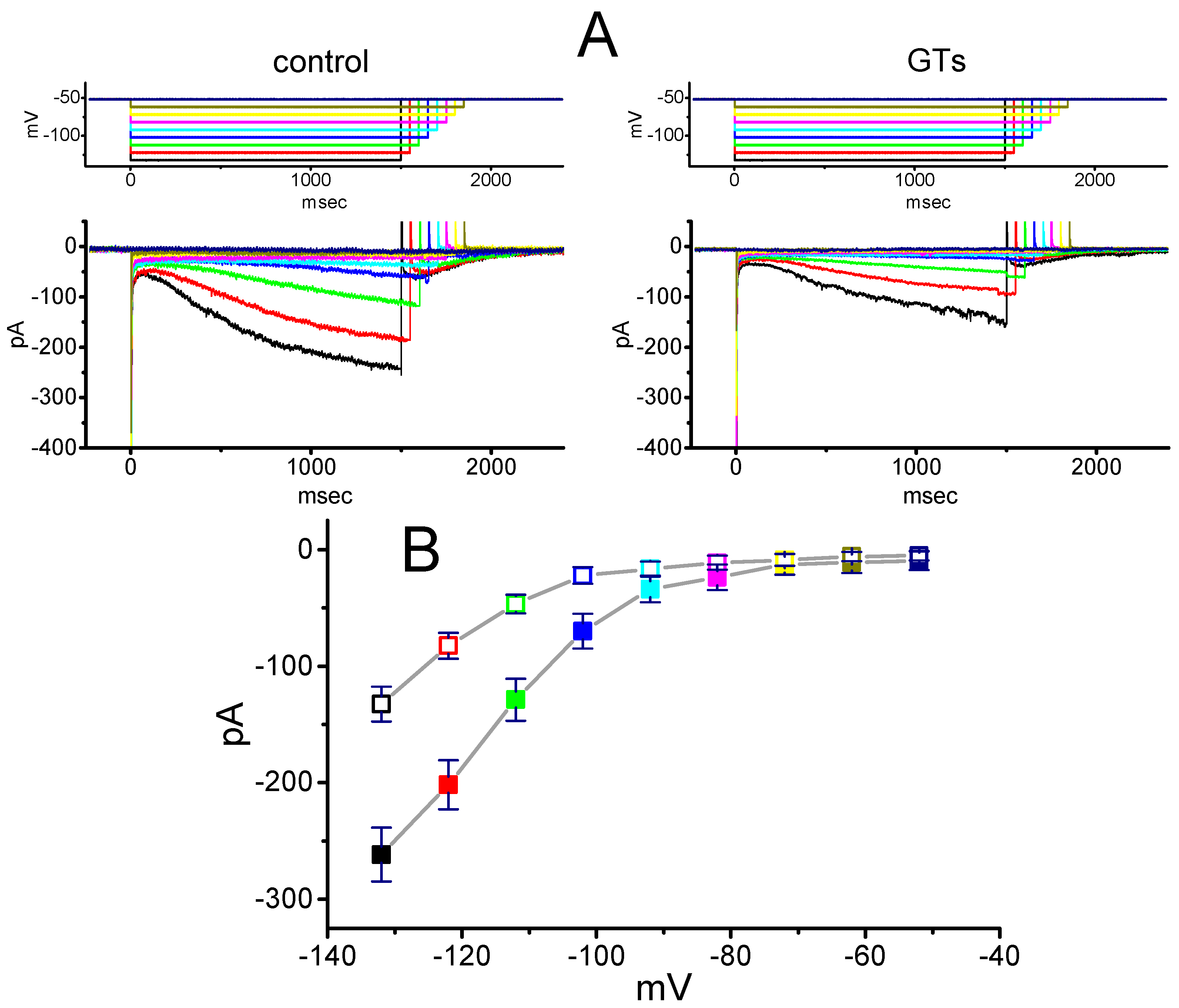
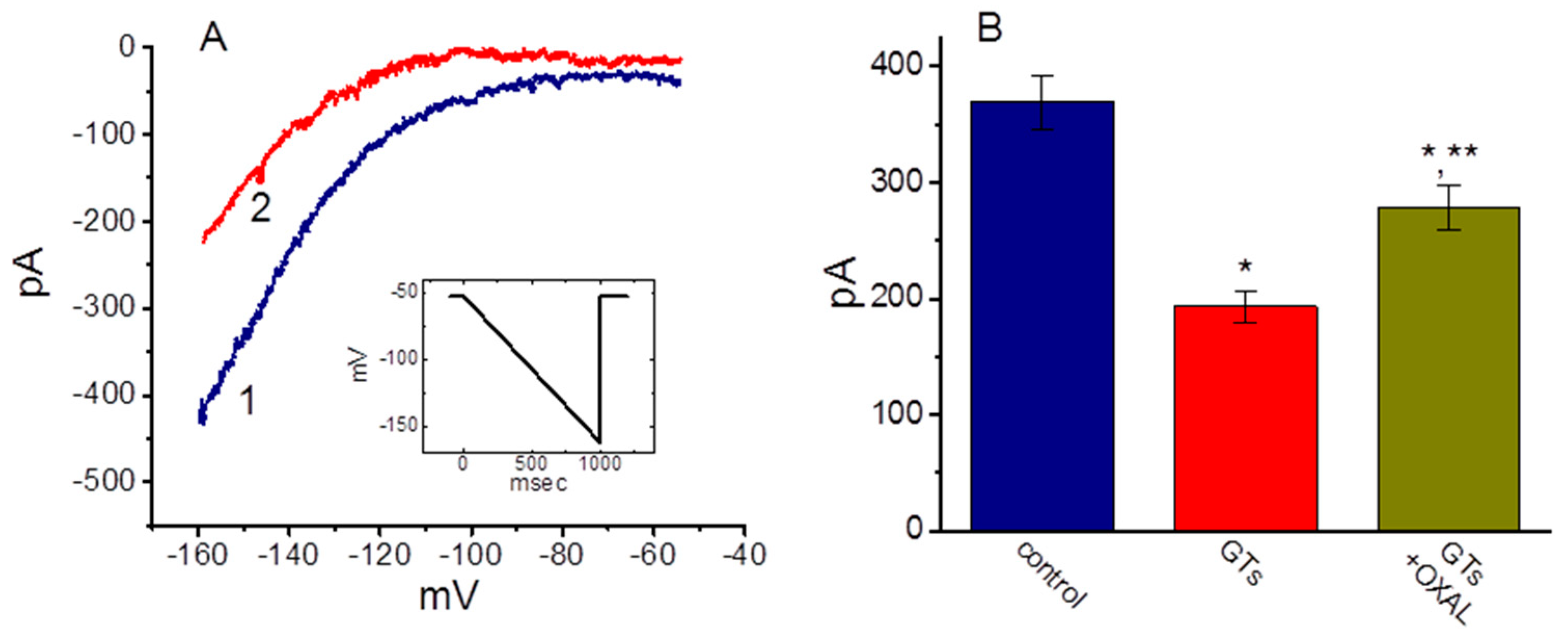
2.2. Effect of GTs on the Averaged I–V Relationship of Ih Recorded from GH3 Cells
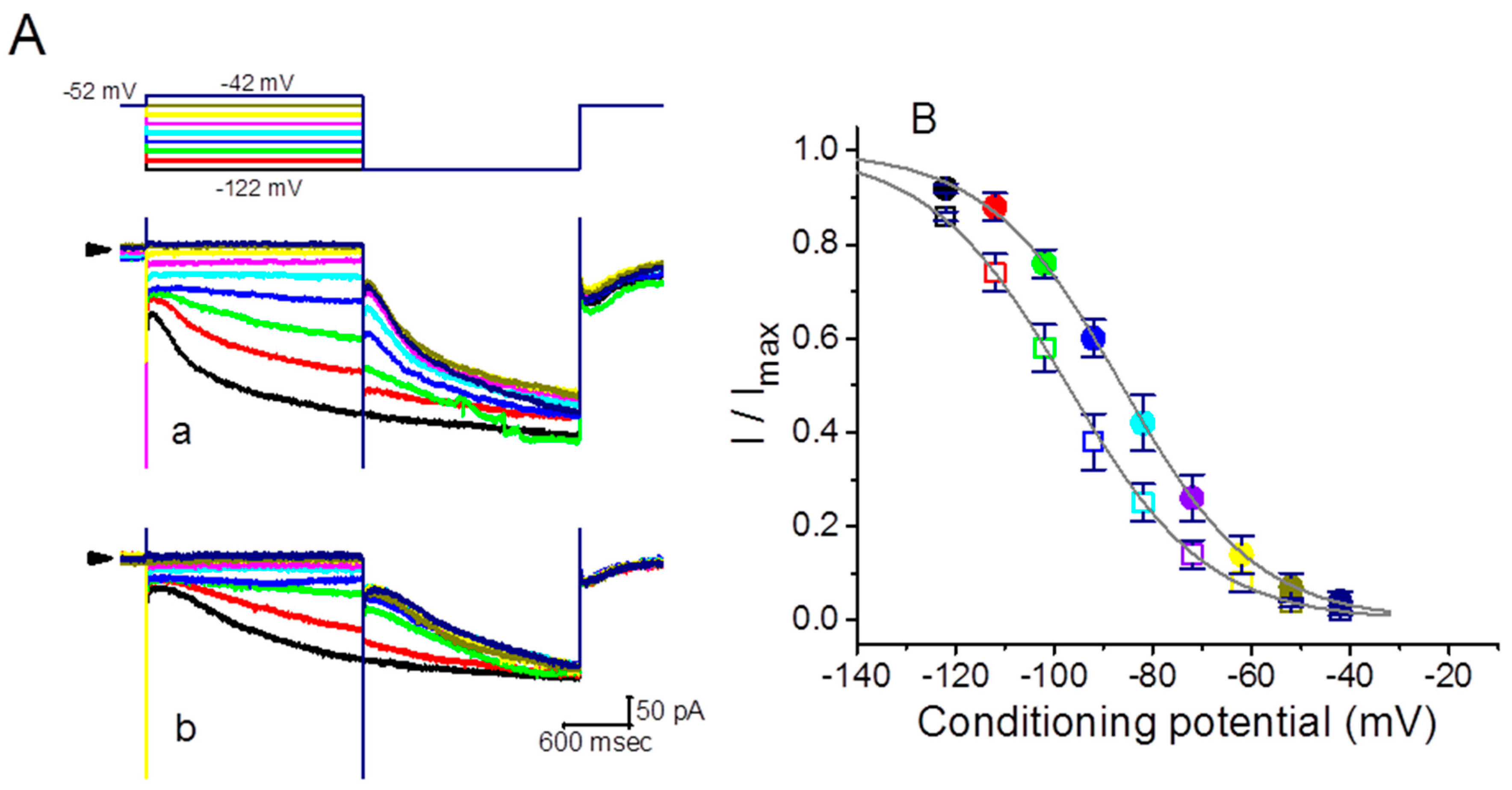
2.3. Modification of the Steady-State Activation Curve of Ih Produced by the Presence of GTs
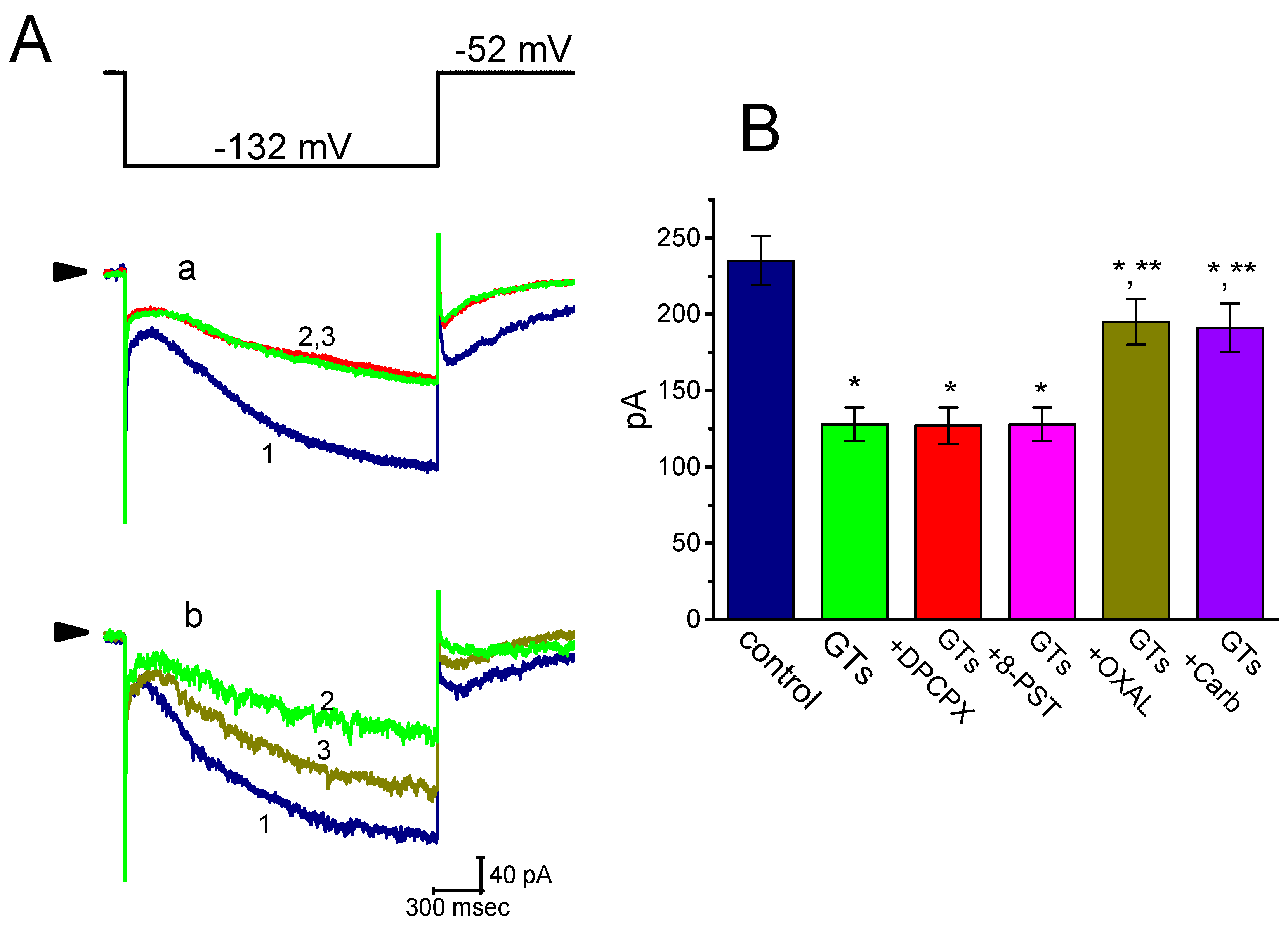
2.4. Comparisons Among the Effects of GTs, GTs Plus 8-Cyclopentyl-1,3-dipropylxanthine (DPCPX), GTs Plus 8-(p-Sulfophenyl)theophylline (8-PST), GTs Plus Oxaliplatin (OXAL) and GTs Plus Carboxiplatin (Carb) in Ih Amplitude
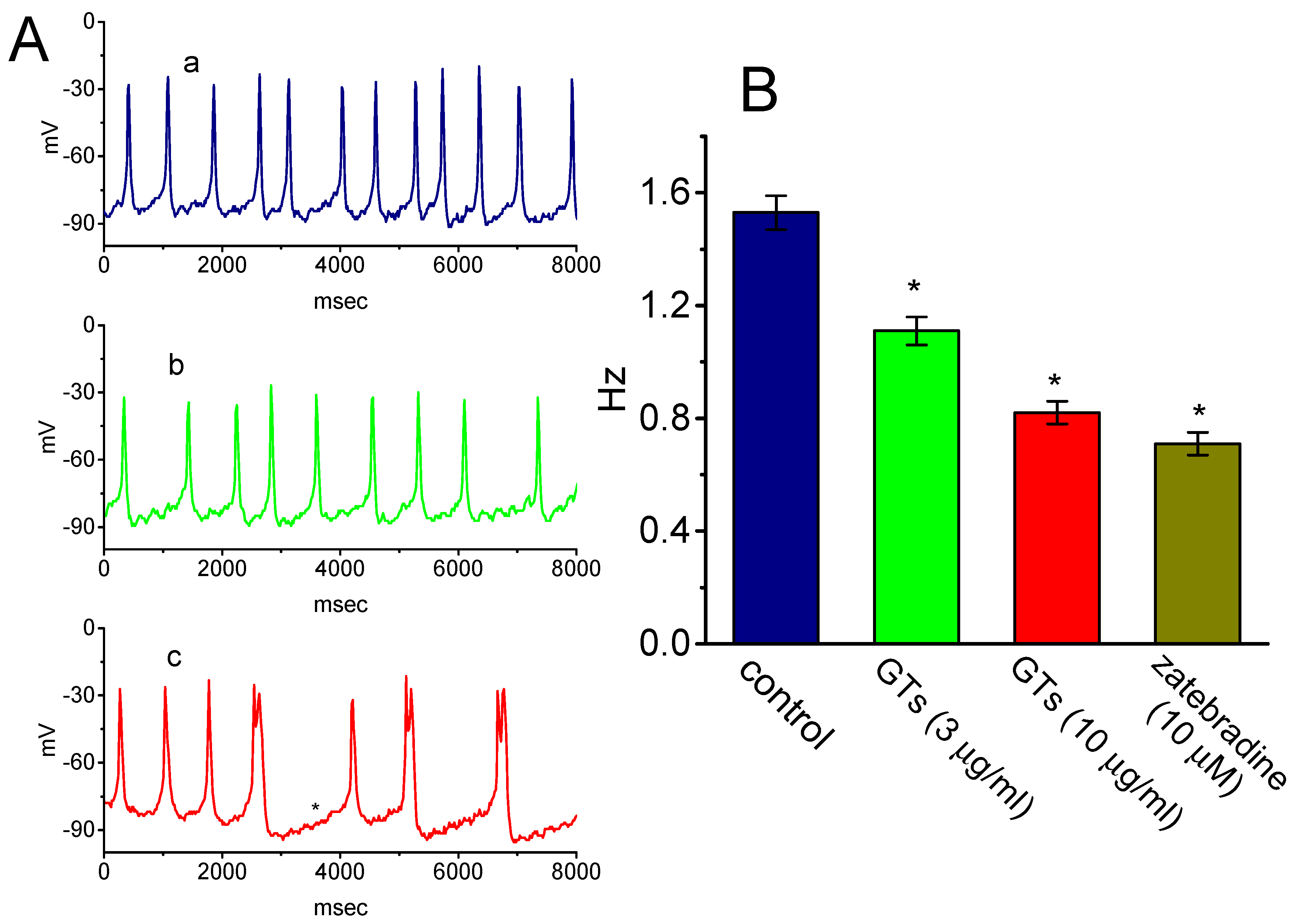
2.5. Effects of GTs on the Firing of Spontaneous Action Potentials (APs) Recorded from GH3 Cells
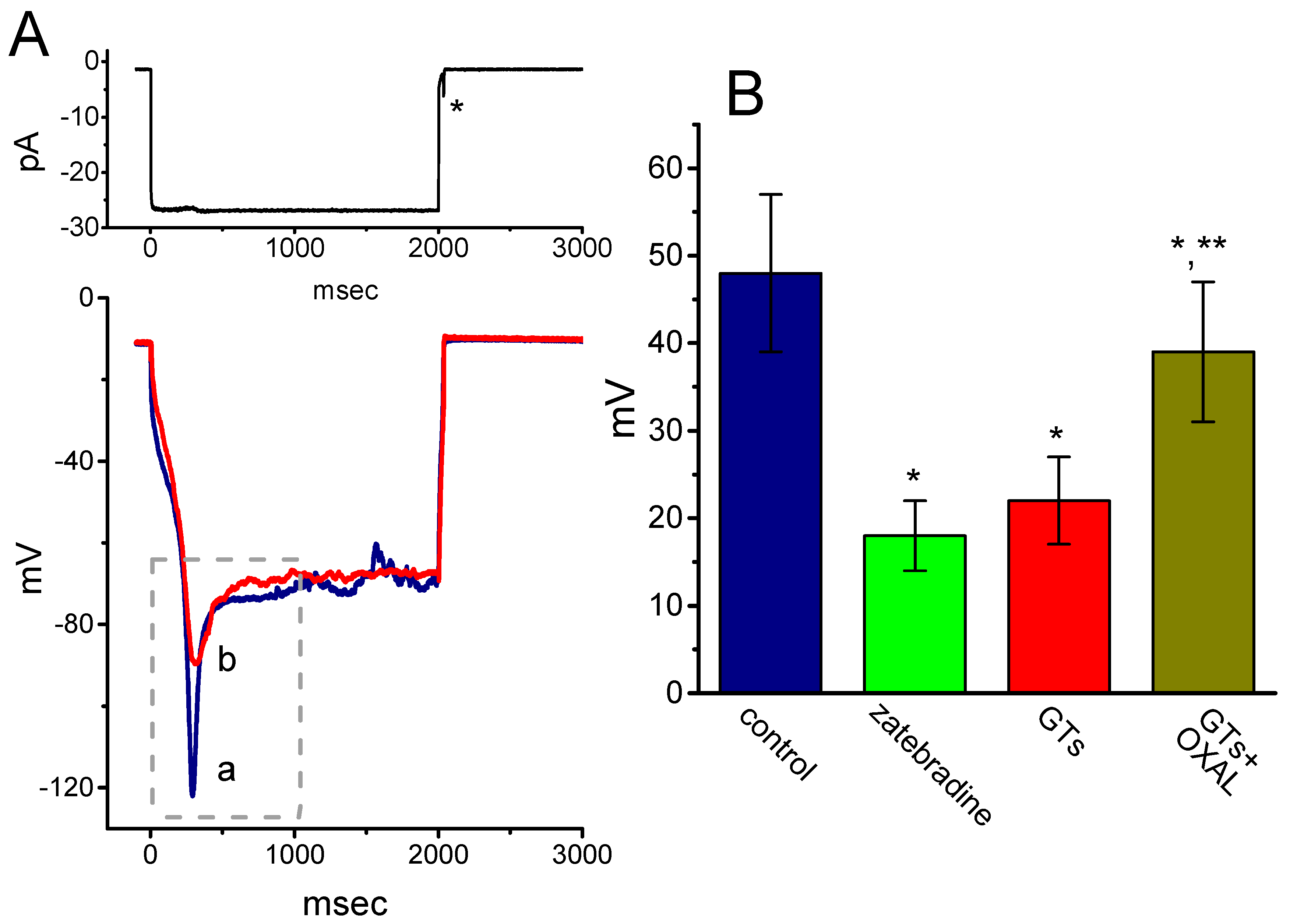
2.6. Effect of GTs on Sag Potential in GH3 Cells
2.7. Effect of GTs on Ih Recorded from HL-1 Cardiomyocytes
3. Materials and Methods
3.1. Chemicals, Drugs and Solutions
3.2. Cell Preparations
3.3. Electrophysiological Measurements
3.4. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | action potential |
| DPCPX | 8-cyclopentyl-1,3-dipropylxanthine |
| GL | Gandoderm lucidum (Língzhī or Reishi) |
| GTs | Ganoderma triterpenoids |
| HCN gene | hyperpolarization-activated cyclic nucleotide-gated gene |
| I-V | current versus voltage |
| IC50 | the concentration required for half-maximal inhibition |
| Ih | hyperpolarization-activated cation current |
| 8-PST | 8-(p-sulfophenyl)theophylline |
| τact | activation time constant |
References
- Shiao, M.S. Natural products of the medicinal fungus Ganoderma lucidum: Occurrence, biological activities, and pharmacological functions. Chem. Rec. 2003, 3, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.F.; Fu, H.Y.; Lu, Y.Y.; Han, Y.C.; Shen, Y.C.; Venkatakrishnan, K.; Wang, C.K.; Golovinskaia, O. Triterpenoids and polysaccharide peptides-enriched Ganoderma lucidum: A randomized, double-blind placebo-controlled crossover study of its antioxidation and hepatoprotective efficacy in healthy volunteers. Pharm. Biol. 2017, 55, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.F. Ganoderma lucidum: Persuasive biologically active constituents and their health endorsement. Biomed. Pharmacother. 2018, 107, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Cor, D.; Knez, Z.; Knez, H.M. Antitumour, Antimicrobial, Antioxidant and Antiacetylcholinesterase Effect of Ganoderma Lucidum Terpenoids and Polysaccharides: A Review. Molecules 2018, 23, 649. [Google Scholar] [CrossRef]
- Xin, H.; Fang, L.; Xie, J.; Qi, W.; Niu, Y.; Yang, F.; Cai, D.; Zhang, Y.; Wen, Z. Identification and Quantification of Triterpenoids in Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes), with HPLC-MS/MS Methods. Int. J. Med. Mushrooms 2018, 20, 919–934. [Google Scholar] [CrossRef]
- Liang, C.; Tian, D.; Liu, Y.; Li, H.; Zhu, J.; Li, M.; Xin, M.; Xia, J. Review of the molecular mechanisms of Ganoderma lucidum triterpenoids: Ganoderic acids A.; C2, D, F, DM, X and Y. Eur. J. Med. Chem. 2019, 174, 130–141. [Google Scholar] [CrossRef]
- Peng, X.; Li, L.; Dong, J.; Lu, S.; Lu, J.; Li, X.; Zhou, L.; Qiu, M. Lanostane-type triterpenoids from the fruiting bodies of Ganoderma applanatum. Phytochemistry 2019, 157, 103–110. [Google Scholar] [CrossRef]
- Kuok, Q.Y.; Yeh, C.Y.; Su, B.C.; Hsu, P.L.; Ni, H.; Liu, M.Y.; Mo, F.E. The triterpenoids of Ganoderma tsugae prevent stress-induced myocardial injury in mice. Mol. Nutr. Food Res. 2013, 57, 1892–1896. [Google Scholar]
- Ma, B.; Ren, W.; Zhou, Y.; Ma, J.; Ruan, Y.; Wem, C.N. Triterpenoids from the spores of Ganoderma lucidum. N. Am. J. Med. Sci. 2011, 3, 495–498. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Ip, F.C.; Zhang, D.M.; Chen, L.X.; Zhang, W.; Li, Y.L.; Ip, N.Y.; Ye, W.C. Triterpenoids with neurotrophic activity from Ganoderma lucidum. Nat. Prod. Res. 2011, 25, 1607–1613. [Google Scholar] [CrossRef]
- Chu, T.T.; Benzie, I.F.; Lam, C.W.; Fok, B.S.; Lee, K.K.; Tomlinson, B. Study of potential cardioprotective effects of Ganoderma lucidum (Lingzhi): Results of a controlled human intervention trial. Br. J. Nutr. 2012, 107, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Qu, Z.Q.; Zeng, Y.S.; Lin, Y.K.; Li, Y.; Chung, P.; Wong, R.; Hägg, U. Neuroprotective effect of preadministration with Ganoderma lucidum spore on rat hippocampus. Exp. Toxicol. Pathol. 2012, 64, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Sudheesh, N.P.; Ajith, T.A.; Janardhanan, K.K. Ganoderma lucidum ameliorate mitochondrial damage in isoproterenol-induced myocardial infarction in rats by enhancing the activities of TCA cycle enzymes and respiratory chain complexes. Int. J. Cardiol. 2013, 165, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Kan, Y.J.; Wu, Y.B.; Yi, J.; Chen, T.Q.; Wu, J.Z. Hepatoprotective effect of ganoderma triterpenoids against oxidative damage induced by tert-butyl hydroperoxide in human hepatic HepG2 cells. Pharm. Biol. 2016, 54, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Kirar, V.; Nehra, S.; Mishra, J.; Rakhee, R.; Saraswat, D.; Misra, K. Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes), as a Cardioprotectant in an Oxygen-Deficient Environment. Int. J. Med. Mushrooms. 2017, 19, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, J.; Ren, J.; Wang, W.; Xiong, W.; Zhang, Y.; Bao, L.; Liu, H. Triterpenes and Meroterpenes with Neuroprotective Effects from Ganoderma leucocontextum. Chem, Biodivers. 2018, 15, e1700567. [Google Scholar] [CrossRef]
- Hsu, P.L.; Lin, Y.C.; Ni, H.; Mo, F.E. Ganoderma Triterpenoids Exert Antiatherogenic Effects in Mice by Alleviating Disturbed Flow-Induced Oxidative Stress and Inflammation. Oxid. Med. Cell. Longev. 2018, 2018, 3491703. [Google Scholar] [CrossRef]
- Veena, R.K.; Ajith, T.A.; Janardhanan, K.K. Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes), Prevents Doxorubicin-Induced Cardiotoxicity in Rats. Int. J. Med. Mushrooms 2018, 20, 761–774. [Google Scholar] [CrossRef]
- Li, T.; Yu, H.; Song, Y.; Zhang, R.; Ge, M. Protective effects of Ganoderma triterpenoids on cadmium-induced oxidative stress and inflammatory injury in chicken livers. J. Trace Elem. Med. Biol. 2019, 52, 118–125. [Google Scholar] [CrossRef]
- Liu, Y.N.; Tong, T.; Zhang, R.R.; Liu, L.M.; Shi, M.L.; Ma, Y.C.; Liu, G.Q. Interdependent nitric oxide and hydrogen peroxide independently regulate the coix seed oil-induced triterpene acid accumulation in Ganoderma lingzhi. Mycologia 2019, 111, 529–540. [Google Scholar] [CrossRef]
- Lou, H.W.; Guo, X.Y.; Zhang, X.C.; Guo, L.Q.; Lin, J.F. Optimization of Cultivation Conditions of Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes) for the Highest Antioxidant Activity and Antioxidant Content. Int. J. Med. Mushrooms 2019, 21, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Han, F.; Luan, S.; Ai, R.; Zhang, P.; Li, H.; Chen, L. Triterpenoids from Ganoderma lucidum and Their Potential Anti-inflammatory Effects. J. Agric. Food Chem. 2019, 67, 5147–5158. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Fan, J.; Liu, Y.; Guo, W.; Cao, H.; Xiao, J.; Wang, Y.; Liu, B. Hepatoprotective activity of Ganoderma lucidum triterpenoids in alcohol-induced liver injury in mice, an iTRAQ-based proteomic analysis. Food Chem. 2019, 271, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Socala, K.; Nieoczym, D.; Grzywnowicz, K.; Stefaniuk, D.; Wlaz, P. Evaluation of Anticonvulsant, Antidepressant-, and Anxiolytic-like Effects of an Aqueous Extract from Cultured Mycelia of the Lingzhi or Reishi Medicinal Mushroom Ganoderma lucidum (Higher Basidiomycetes) in Mice. Int. J. Med. Mushrooms 2015, 17, 209–218. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Y. Anti-inflammatory, anti-nociceptive and sedative-hypnotic activities of lucidone D extracted from Ganoderma lucidum. Cell Mol. Biol. 2019, 65, 37–42. [Google Scholar] [CrossRef]
- Belardinelli, L.; Giles, W.R.; West, A. Ionic mechanisms of adenosine actions in pacemaker cells from rabbit heart. J Physiol 1988, 405, 615–633. [Google Scholar] [CrossRef]
- Irisawa, H.; Brown, H.F.; Giles, W. Cardiac pacemaking in the sinoatrial node. Physiol. Rev. 1993, 73, 197–227. [Google Scholar] [CrossRef]
- DiFrancesco, D.; Borer, J.S. The funny current: Cellular basis for the control of heart rate. Drugs 2007, 67, 15–24. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wang, Y.J.; Wu, P.Y.; Wu, S.N. Tramadol-induced block of hyperpolarization-activated cation current in rat pituitary lactotrophs. Naunyn. Schmiedebergs Arch. Pharmacol. 2009, 379, 127–135. [Google Scholar] [CrossRef]
- He, C.; Chen, F.; Li, B.; Hu, Z. Neurophysiology of HCN channels: From cellular functions to multiple regulations. Prog. Neurobiol. 2014, 112, 1–23. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endocr. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef] [PubMed]
- Datunashvili, M.; Chaudhary, R.; Zobeiri, M.; Luttjohann, A.; Mergia, E.; Baumann, A.; Balfanz, S.; Budde, B.; van Luijtelaar, G.; Pape, H.C.; et al. Modulation of Hyperpolarization-Activated Inward Current and Thalamic Activity Modes by Different Cyclic Nucleotides. Front. Cell Neurosci. 2018, 12, 369. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, V.; Sartiani, L.; Mugelli, A.; Romanelli, M.N.; Cerbai, E. Hyperpolarization-activated cyclic-nucleotide-gated channels: Pathophysiological, developmental, and pharmacological insights into their function in cellular excitability. Can. J. Physiol. Pharmacol. 2018, 96, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Byczkowicz, N.; Eshra, A.; Montanaro, J.; Trevisiol, A.; Hirrlinger, J.; Kole, M.H.; Shigemoto, R.; Hallermann, S. HCN channel-mediated neuromodulation can control action potential velocity and fidelity in central axons. Elife 2019, 8. [Google Scholar] [CrossRef]
- Hao, X.M.; Xu, R.; Chen, A.Q.; Sun, F.J.; Wang, Y.; Liu, H.X.; Chen, H.; Xue, Y.; Chen, L. Endogenous HCN Channels Modulate the Firing Activity of Globus Pallidus Neurons in Parkinsonian Animals. Front. Aging Neurosci. 2019, 11, 190. [Google Scholar] [CrossRef]
- Romanelli, M.N.; Cerbai, E.; Dei, S.; Guandalini, L.; Martelli, C.; Martini, E.; Scapecchi, S.; Teodori, E.; Mugelli, A. Design, synthesis and preliminary biological evaluation of zatebradine analogues as potential blockers of the hyperpolarization-activated current. Bioorg. Med. Chem. 2005, 13, 1211–1220. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, J.; Meng, Z.; Zhou, S.; Peng, J.; Chen, S.; Wang, Q.; Sun, K. Pharmacology of ivabradine and the effect on chronic heart failure. Curr. Top Med. Chem. 2019. [Google Scholar] [CrossRef]
- Hsiao, H.T.; Liu, Y.C.; Liu, P.Y.; Wu, S.N. Concerted suppression of Ih and activation of IK(M) by ivabradine, an HCN-channel inhibitor, in pituitary cells and hippocampal neurons. Brain Res. Bull. 2019, 149, 11–20. [Google Scholar] [CrossRef]
- Wu, S.N.; Liu, S.I.; Hwang, T.L. Activation of muscarinic K+ channels by extracellular ATP and UTP in rat atrial myocytes. J. Cardiovasc. Pharmaco. 1998, 31, 203–211. [Google Scholar] [CrossRef]
- Resta, F.; Micheli, L.; Laurino, A.; Spinelli, V.; Mello, T.; Mello, T.; Sartiani, L.; Di Cesare Mannelli, L.; Cerbai, E.; Ghelardini, C.; et al. Selective HCN1 block as a strategy to control oxaliplatin-induced neuropathy. Neuropharmacology 2018, 131, 403–413. [Google Scholar] [CrossRef]
- Simasko, S.M.; Sankaranarayanan, S. Characterization of a hyperpolarization-activated cation current in rat pituitary cells. Am. J. Physiol. 1997, 272, E405–E414. [Google Scholar] [CrossRef] [PubMed]
- Van Bogaert, P.P.; Pittoors, F. Use-dependent blockade of cardiac pacemaker current (If) by cilobradine and zatebradine. Eur. J. Pharmacol. 2003, 478, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Albertson, A.J.; Bohannon, A.S.; Hablitz, J.J. HCN Channel Modulation of Synaptic Integration in GABAergic Interneurons in Malformed Rat Neocortex. Front. Cell Neurosci. 2017, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Claycomb, W.C.; Lanson, N.A.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J., Jr. HL-1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Wu, S.N. Activation of voltage-gated sodium current and inhibition of erg-mediated potassium current caused by telmisartan, an antagonist of angiotensin II type-1 receptor, in HL-1 atrial cardiomyocytes. Clin. Exp. Pharmacol. Physio. 2018, 45, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Dongmo, A.B.; Azebaze, A.G.; Donfack, F.M.; Dimo, T.; Nkeng-Efouet, P.A.; Devkota, K.P.; Sontia, B.; Wagner, H.; Sewald, N.; Vierling, W. Pentacyclic triterpenoids and ceramide mediate the vasorelaxant activity of Vitex cienkowskii via involvement of NO/cGMP pathway in isolated rat aortic rings. J. Ethnopharmacol. 2011, 133, 204–212. [Google Scholar] [CrossRef]
- Wei, J.C.; Wang, A.H.; Wei, Y.L.; Huo, X.K.; Tian, X.G.; Feng, L.; Ma, X.C.; Wang, C.; Huang, S.S.; Jia, J.M. Chemical characteristics of the fungus Ganoderma lucidum and their inhibitory effects on acetylcholinesterase. J. Asian Nat. Prod. Res. 2018, 20, 992–1001. [Google Scholar] [CrossRef]
- Cheung, H.Y.; Ng, C.W.; Hood, D.J. Identification and quantification of base and nucleoside markers in extracts of Ganoderma lucidum, Ganoderma japonicum and Ganoderma capsules by micellar electrokinetic chromatography. J. Chromatogr. A 2001, 911, 119–126. [Google Scholar] [CrossRef]
- Gao, J.L.; Leung, K.S.; Wang, Y.T.; Lai, C.M.; Li, S.P.; Hu, L.F.; Lu, G.H.; Jiang, Z.H.; Yu, Z.L. Qualitative and quantitative analyses of nucleosides and nucleobases in Ganoderma spp. by HPLC-DAD-MS. J. Pharm. Biomed. Anal. 2007, 44, 807–811. [Google Scholar] [CrossRef]
- Khan, M.S.; Parveen, R.; Mishra, K.; Tulsawani, R.; Ahmad, S. Determination of nucleosides in Cordyceps sinensis and Ganoderma lucidum by high performance liquid chromatography method. J. Pharm. Bioallied. Sci. 2015, 7, 264–266. [Google Scholar]
- Zaza, A.; Rocchetti, M.; DiFrancesco, D. Modulation of the hyperpolarization-activated current (If) by adenosine in rabbit sinoatrial myocytes. Circulation 1996, 94, 734–741. [Google Scholar] [CrossRef] [PubMed]
- You, B.J.; Tien, N.; Lee, M.H.; Bao, B.Y.; Wu, Y.S.; Hu, T.C.; Lee, H.Z. Induction of apoptosis and ganoderic acid biosynthesis by cAMP signaling in Ganoderma lucidum. Sci. Rep. 2017, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Collado Mateo, D.; Pazzi, F.; Dominguez Munoz, F.J.; Martin Martinez, J.P.; Olivares, P.R.; Gusi, N.; Adsuar, J.C. Ganoderma Lucidum Improves Physical Fitness in Women with Fibromyalgia. Nutr. Hosp. 2015, 32, 2126–2135. [Google Scholar] [PubMed]
- Norberg, E.; Karlsson, M.; Korenovska, O.; Szydlowski, S.; Silberberg, G.; Uhlén, P.; Orrenius, S.; Zhivotovsky, B. Critical role for hyperpolarization-activated cyclic nucleotide-gated channel 2 in the AIF-mediated apoptosis. EMBO J. 2010, 29, 3869–3878. [Google Scholar] [CrossRef]
- Hurtado, R.; Smith, C.S. Hyperpolarization-activated cation and T-type calcium ion channel expression in porcine and human renal pacemaker tissues. J. Anat. 2016, 228, 812–825. [Google Scholar] [CrossRef]
- He, F.; Yang, Z.; Dong, X.; Fang, Z.; Liu, Q.; Hu, X.; Yu, S.; Li, L. The role of HCN channels in peristaltic dysfunction in human ureteral tuberculosis. Int. Urol. Nephrol. 2018, 50, 639–645. [Google Scholar] [CrossRef]
- Mader, F.; Muller, S.; Krause, L.; Springer, A.; Kernig, K.; Protzel, C.; Porath, K.; Rackow, S.; Wittstock, T.; Frank, M.; et al. Hyperpolarization-Activated Cyclic Nucleotide-Gated Non-selective (HCN) Ion Channels Regulate Human and Murine Urinary Bladder Contractility. Front. Physiol. 2018, 9, 753. [Google Scholar] [CrossRef]
- Furst, O.; D’Avanzo, N. Isoform dependent regulation of human HCN channels by cholesterol. Sci. Rep. 2015, 5, 14270. [Google Scholar] [CrossRef]
- He, Y.M.; Zhang, Q.; Zheng, M.; Fan, Z.H.; Li, Y.H.; Zhang, D.; Zhang, Z.; Yuan, S.S.; Wang, Y.Y.; Zhou, P.; et al. Protective effects of a G. lucidum proteoglycan on INS-1 cells against IAPP-induced apoptosis via attenuating endoplasmic reticulum stress and modulating CHOP/JNK pathways. Int. J. Biol. Macromol. 2018, 106, 893–900. [Google Scholar] [CrossRef]
- Kim, H.B.; Hong, Y.J.; Park, H.J.; Ahn, Y.; Jeong, M.H. Effects of Ivabradine on Left Ventricular Systolic Function and Cardiac Fibrosis in Rat Myocardial Ischemia-Reperfusion Model. Chonnam. Med. J. 2018, 54, 167–172. [Google Scholar] [CrossRef]
- Huang, M.H.; Liu, P.Y.; Wu, S.N. Characterization of Perturbing Actions by Verteporfin, a Benzoporphyrin Photosensitizer, on Membrane Ionic Currents. Front. Chem. 2019, 7, 566. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors upon requests. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-T.; Gao, Z.-H.; Lo, Y.-C.; Wu, S.-N. Evidence for Effective Inhibitory Actions on Hyperpolarization-Activated Cation Current Caused by Ganoderma Triterpenoids, the Main Active Constitutents of Ganoderma Spores. Molecules 2019, 24, 4256. https://doi.org/10.3390/molecules24234256
Chang W-T, Gao Z-H, Lo Y-C, Wu S-N. Evidence for Effective Inhibitory Actions on Hyperpolarization-Activated Cation Current Caused by Ganoderma Triterpenoids, the Main Active Constitutents of Ganoderma Spores. Molecules. 2019; 24(23):4256. https://doi.org/10.3390/molecules24234256
Chicago/Turabian StyleChang, Wei-Ting, Zi-Han Gao, Yi-Ching Lo, and Sheng-Nan Wu. 2019. "Evidence for Effective Inhibitory Actions on Hyperpolarization-Activated Cation Current Caused by Ganoderma Triterpenoids, the Main Active Constitutents of Ganoderma Spores" Molecules 24, no. 23: 4256. https://doi.org/10.3390/molecules24234256
APA StyleChang, W.-T., Gao, Z.-H., Lo, Y.-C., & Wu, S.-N. (2019). Evidence for Effective Inhibitory Actions on Hyperpolarization-Activated Cation Current Caused by Ganoderma Triterpenoids, the Main Active Constitutents of Ganoderma Spores. Molecules, 24(23), 4256. https://doi.org/10.3390/molecules24234256