
Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin


 ,
, 
Abstract

1. Introduction
2. Results
2.1. Palynological Analysis
2.2. Instrumental Color Measurement
2.3. Pollen Load Size Distribution
2.4. Protein Determination
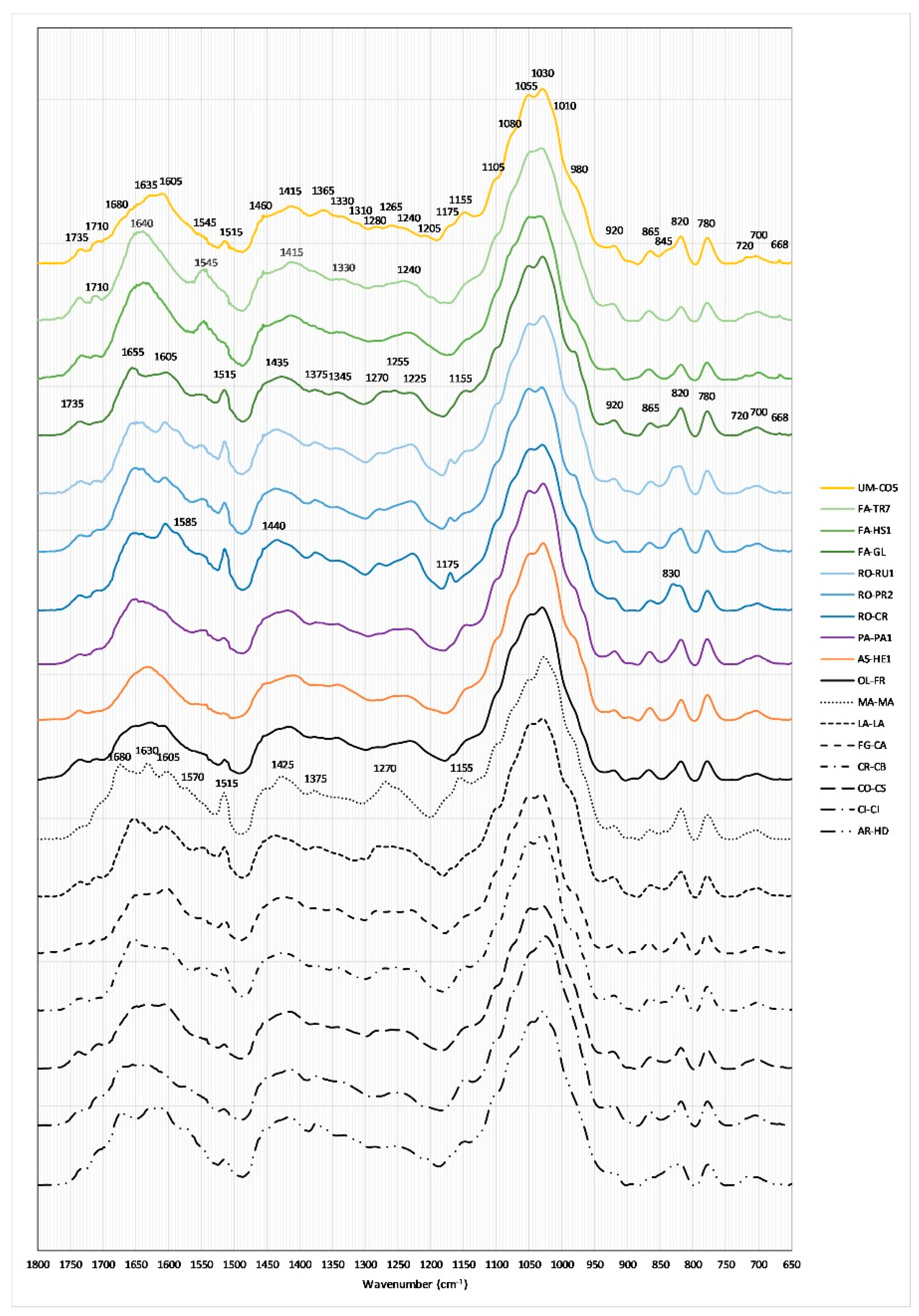
2.5. FTIR-ATR Spectral Characterization
3. Discussion
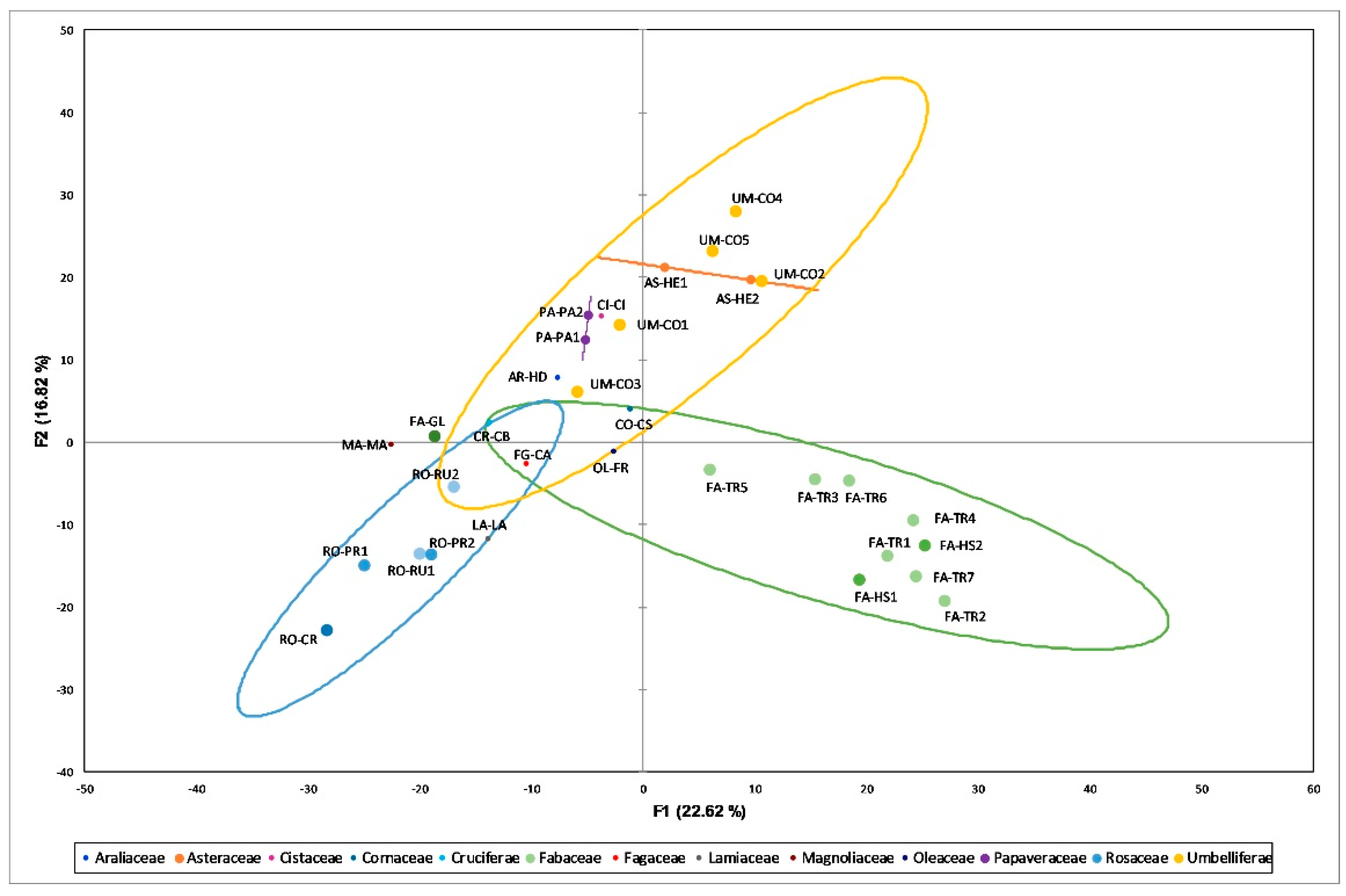
3.1. Cluster Analysis
3.2. Principal Component Analysis (PCA)
4. Material and Methods
4.1. Pollen Samples
4.2. Palynological Analysis
- group (gr.): when referring to a specific species (e.g., Trifolium repens gr.) and one is certain of the belonging of the unknown pollen to the same botanical genus or to a related genus;
- form (f.): when referring to a botanical genus (e.g., Eucalyptus f., or “M” form for the Labiatae) and one is certain of belonging to the same botanical family;
- type (t.): when the unknown pollen has a similarity with another known pollen but is not possible to state that it belongs to the same botanical family (we can refer to a genus or a family, e.g., Moraceae t.) [22].
4.3. Visual Characterization
4.4. Instrumental Color Measurement
4.5. Pollen Load Size Distribution
4.6. Moisture Determination
4.7. Total Protein Content (%)
4.8. FTIR-ATR Data Acquisition and Data Processing
4.9. Multivariate Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Di Marco, G.; Manfredini, A.; Leonardi, D.; Canuti, L.; Impei, S.; Gismondi, A.; Canini, A. Geographical, botanical and chemical profile of monofloral Italian honeys as food quality guarantee and territory brand. Plant Biosyst. 2017, 151, 450–463. [Google Scholar] [CrossRef]
- Dimou, M.; Thrasyvoulou, A.; Tsirakoglou, V. Efficient use of pollen traps to determine the pollen flora used by honey bees. J. Apic. Res. 2006, 45, 42–46. [Google Scholar] [CrossRef]
- Gabriele, M.; Parri, E.; Felicioli, A.; Sagona, S.; Pozzo, L.; Biondi, C.; Domenici, V.; Pucci, L. Phytochemical composition and antioxidant activity of Tuscan bee pollen of different botanic origins. Ital. J. Food Sci. 2015, 27, 248–259. [Google Scholar]
- Castiglioni, S.; Stefano, M.; Astolfi, P.; Carloni, P. Chemometric approach to the analysis of antioxidant properties and color of typical Italian monofloral honeys. Int. J. Food Sci. Tech. 2017, 52, 1138–1146. [Google Scholar] [CrossRef]
- Ciappini, M.C. Polyhenolic profile of floral honeys in correlation with their pollen spectrum. J. Apicult. Res. 2019, 58, 772–779. [Google Scholar] [CrossRef]
- Castiglioni, S.; Stefano, M.; Pisani, M.; Carloni, P. Geographical characterisation of multifloral honeys from the Marche region (Italy) according to their antioxidant activity and color using a chemometric approach. Int. J. Food Sci. Tech. 2018, 53, 571–581. [Google Scholar] [CrossRef]
- Conti, I.; Medrzycki, P.; Argenti, C.; Meloni, M.; Vecchione, V.; Boi, M.; Mariotti, M.G. Sugar and protein content in different monofloral pollens—Building a database. Bull. Insectology 2016, 69, 318–320. [Google Scholar]
- Rocchetti, G.; Castiglioni, S.; Maldarizzi, G.; Carloni, P.; Lucini, L. UHPLC-ESI-QTOF-MS phenolic profiling and antioxidant capacity of bee pollen from different botanical origin. Int. J. Food Sci. Tech. 2019, 54, 335–346. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and bee bread as new health-oriented products: A review. Trends Food Sci. Tech. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Salazar-González, C.Y.; Rodríguez-Pulido, F.J.; Terrab, A.; Díaz-Moreno, C.; Fuenmayor, C.A.; Heredia, F.J. Analysis of Multifloral Bee Pollen Pellets by Advanced Digital Imaging Applied to Functional Food Ingredients. Plant. Foods Hum. Nutr. 2018, 73, 328–335. [Google Scholar] [CrossRef]
- Liolios, V.; Tananaki, C.; Papaioannou, A.; Kanelis, D.; Rodopoulou, M.-A.; Argena, N. Mineral content in monofloral bee pollen: Investigation of the effect of the botanical and geographical origin. J. Food Meas. Charact. 2019, 13, 1674–1682. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Barać, M.B.; Stanojević, S.P.; Milojković-Opsenica, D.M.; Tešić, Ž.L.; Šikoparija, B.; Radišić, P.; Prentović, M.; Pešić, M.B. Physicochemical composition and techno-functional properties of bee pollen collected in Serbia. LWT Food Sci. Technol. 2015, 62, 301–309. [Google Scholar] [CrossRef]
- Yang, K.; Wu, D.; Ye, X.; Liu, D.; Chen, J.; Sun, P. Characterization of Chemical Composition of Bee Pollen in China. J. Agric. Food Chem. 2013, 61, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Feás, X.; Vázquez-Tato, M.P.; Estevinho, L.; Seijas, J.A.; Iglesias, A. Organic Bee Pollen: Botanical Origin, Nutritional Value, Bioactive Compounds, Antioxidant Activity and Microbiological Quality. Molecules 2012, 17, 8359–8377. [Google Scholar] [CrossRef] [PubMed]
- Liolios, V.; Tananaki, C.; Dimou, M.; Kanelis, D.; Goras, G.; Karazafiris, E.; Thrasyvoulou, A. Ranking pollen from bee plants according to their protein contribution to honey bees. J. Apic. Res. 2015, 54, 582–592. [Google Scholar] [CrossRef]
- Forcone, A.; Aloisi, P.V.; Ruppel, S.; Muñoz, M. Botanical composition and protein content of pollen collected by Apis mellifera L. in the north-west of Santa Cruz (Argentinean Patagonia). Grana 2011, 50, 30–39. [Google Scholar] [CrossRef]
- Bleha, R.; Shevtsova, T.; Sinica, A.; Kruzik, V.; Brindza, J. Morphology, physicochemical properties and antioxidant capacity of bee pollens. Czech. J. Food Sci. 2019, 37, 1–8. [Google Scholar] [CrossRef]
- Kasprzyk, I.; Depciuch, J.; Grabek-Lejko, D.; Parlinska-Wojtan, M. FTIR-ATR spectroscopy of pollen and honey as a tool for unifloral honey authentication. The case study of rape honey. Food Control. 2018, 84, 33–40. [Google Scholar] [CrossRef]
- Xu, X.; Zheng, Y.; Chen, X.; Zhu, F.; Miao, X. Identification of cattail pollen, pine pollen and bee pollen by Fourier transform infrared spectroscopy and two-dimensional correlation infrared spectroscopy. J. Mol. Struct. 2018, 1167, 78–81. [Google Scholar] [CrossRef]
- Svečnjak, L.; Bubalo, D.; Baranović, G.; Novosel, H. Optimization of FTIR-ATR spectroscopy for botanical authentication of unifloral honey types and melissopalynological data prediction. Eur. Food Res. Technol. 2015, 240, 1101–1115. [Google Scholar] [CrossRef]
- Cozzolino, D. Infrared Spectroscopy as a Versatile Analytical Tool for the Quantitative Determination of Antioxidants in Agricultural Products, Foods and Plants. Antioxidants 2015, 4, 482–497. [Google Scholar] [CrossRef] [PubMed]
- Von Der Ohe, W.; Persano Oddo, L.; Piana, M.L.; Morlot, M.; Martin, P. Harmonized methods of melissopalynology. Apidologie 2004, 35, S18–S25. [Google Scholar] [CrossRef]
- Castiglioni, S.; Damiani, E.; Astolfi, P.; Carloni, P. Influence of steeping conditions (time, temperature, and particle size) on antioxidant properties and sensory attributes of some white and green teas. Int. J. Food Sci. Nutr. 2015, 66, 491–497. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: The pollen samples are available from the authors for limited time. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Family | Genus | Determined Species and Predominance | Palynological Analysis |
|---|---|---|---|---|
| AR-HD | Araliaceae | Hedera | H. helix L. 100% | H. helix L. 100% |
| CI-CI | Cistaceae | Cistus | Cistus incanus/creticus 86%, Castanea sativa Miller 10% | |
| CO-CS | Cornaceae | Cornus | C. sanguinea L. 100% | C. sanguinea L. 100% |
| CR-CB | Cruciferae | Brassica | Cruciferae Brassica type 100% | |
| FG-CA | Fagaceae | Castanea | C. sativa Miller 100% | C. sativa Miller 100% |
| LA-LA | Lamiaceae | Lamium | Lamiaceae L form 100% | |
| MA-MA | Magnoliaceae | Magnolia | Magnolia 69%, Plantago 12%, Umbelliferae 7%, Asteraceae S Form (Carduus) 5% | |
| OL-FR | Oleaceae | Fraxinus | F. ornus L. 90% | F. ornus L. 90%, Quercus robur gr. 10% |
| AS-HE1 | Asteraceae | Helianthus | H. annuus L. 45% | Asteraceae T form (Taraxacum and Cichorium) 55%, H. annuus L. 45% |
| AS-HE2 | Asteraceae | Helianthus | H. annuus L. 77% | H. annuus L. 77%, Asteraceae T form (Taraxacum and Cichorium) 23% |
| PA-PA1 | Papaveraceae | Papaver | P. rhoeas L. 100% | P. rhoeas L. 100% |
| PA-PA2 | Papaveraceae | Papaver | P. rhoeas L. 100% | P. rhoeas L. 100% |
| RO-CR | Rosaceae | Crataegus | C. monogyna Jacq. 70% | C. monogyna Jacq. 70%, Salix 20%, Quercus robur gr. 10% |
| RO-PR1 | Rosaceae | Prunus | Prunus f. 100% | |
| RO-PR2 | Rosaceae | Prunus | Prunus f. 80%, Salix 18% | |
| RO-RU1 | Rosaceae | Rubus | R. ulmifolius Schott 100% | R. ulmifolius Schott 100% |
| RO-RU2 | Rosaceae | Rubus | R. ulmifolius Schott 100% | R. ulmifolius Schott 100% |
| FA-GL | Fabaceae | Gleditsia | G. triacanthos L. 100% | G. triacanthos L. 100% |
| FA-HS1 | Fabaceae | Hedysarum | H. coronarium L. 100% | H. coronarium L. 100% |
| FA-HS2 | Fabaceae | Hedysarum | H. coronarium L. 100% | H. coronarium L. 100% |
| FA-TR1 | Fabaceae | Trifolium | T. alexandrinum L. 98% | T. alexandrinum L. 98% |
| FA-TR2 | Fabaceae | Trifolium | T. alexandrinum L. 97% | T. alexandrinum L. 97% |
| FA-TR3 | Fabaceae | Trifolium | T. alexandrinum L. 81% | T. alexandrinum L. 81%, T. repens L. 17% |
| FA-TR4 | Fabaceae | Trifolium | T. alexandrinum L. 90% | T. alexandrinum L. 90%, T. repens L. 10% |
| FA-TR5 | Fabaceae | Trifolium | T. alexandrinum L. 98% | T. alexandrinum L. 98% |
| FA-TR6 | Fabaceae | Trifolium | T. alexandrinum L. 59% | T. alexandrinum L. 59%, T. repens L. 17%, Galega officinalis L. 6%, Papaver rhoeas L. 5%, Rubus ulmifolius Schott 5%, Cruciferae 4%, Lotus corniculatus L. 3% |
| FA-TR7 | Fabaceae | Trifolium | T. alexandrinum L. 100% | T. alexandrinum L. 100%, |
| UM-CO1 | Umbelliferae | Coriandrum | C. sativum L. 100% | Daucus and Coriandrum gr. 100% |
| UM-CO2 | Umbelliferae | Coriandrum | C. sativum L. 100% | Daucus and Coriandrum gr. 100% |
| UM-CO3 | Umbelliferae | Coriandrum | C. sativum L. 100% | Daucus and Coriandrum gr. 100% |
| UM-CO4 | Umbelliferae | Coriandrum | C. sativum L. 100% | Daucus and Coriandrum gr. 100% |
| UM-CO5 | Umbelliferae | Coriandrum | C. sativum L. 100% | Daucus and Coriandrum gr. 100% |
| Sample | Morphology | Color | Protein | Pollen Load Size 2 Distribution 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L* | a* | b* | Content 1 | >2800 | >2400 | >2000 | >1690 | >1400 | <1400 | ||
| AR-HD | Yellow Ocher | 67.6 | 7.9 | 47.8 | 26.7 | 3 | 22 | 58 | 14 | 3 | 0 |
| CI-CI | Yellow Orange | 56.5 | 12.3 | 48.5 | 18.3 | 24 | 48 | 26 | 1 | 0 | 0 |
| CO-CS | Ocher Big | 56.0 | 7.0 | 36.0 | 17.1 | 23 | 66 | 10 | 0 | 0 | 0 |
| CR-CB | Yellow | 60.4 | 5.8 | 46.9 | 23.9 | 2 | 30 | 61 | 6 | 1 | 0 |
| FG-CA | Yellow | 63.8 | 5.7 | 50.3 | 23.9 | 2 | 20 | 66 | 11 | 1 | 0 |
| LA-LA | Light Beige | 63.5 | 5.0 | 29.4 | 26.2 | 14 | 53 | 30 | 2 | 0 | 0 |
| MA-MA | Dirty White | 67.5 | 6.5 | 33.9 | 25.5 | 0 | 0 | 41 | 31 | 23 | 5 |
| OL-FR | Yellow | 63.7 | 6.4 | 50.1 | 23.0 | 2 | 15 | 68 | 12 | 2 | 1 |
| AS-HE1 | Vivid Orange | 55.6 | 21.6 | 65.4 | 14.5 | 1 | 10 | 49 | 30 | 9 | 1 |
| AS-HE2 | Vivid Orange | 56.8 | 22.3 | 69.4 | 13.6 | 1 | 10 | 58 | 23 | 8 | 1 |
| PA-PA1 | Green | 28.6 | −1.9 | 18.9 | 23.8 | 0 | 64 | 25 | 7 | 4 | 0 |
| PA-PA2 | Green | 29.8 | 2.0 | 18.2 | 24.7 | 0 | 29 | 56 | 8 | 7 | 0 |
| RO-CR | Light Yellow | 66.3 | 3.8 | 40.8 | 27.4 | 2 | 32 | 54 | 9 | 1 | 1 |
| RO-PR1 | Olive Green | 53.3 | 3.4 | 23.7 | 30.2 | 13 | 50 | 32 | 3 | 1 | 0 |
| RO-PR2 | Green Beige | 57.3 | 3.0 | 30.0 | 25.8 | 0 | 5 | 41 | 39 | 14 | 0 |
| RO-RU1 | Light Green | 56.0 | 3.7 | 18.6 | 25.5 | 0 | 27 | 50 | 13 | 8 | 0 |
| RO-RU2 | Two-Tone Green | 50.1 | 3.9 | 14.6 | 24.0 | 2 | 16 | 63 | 16 | 3 | 0 |
| FA-GL | Yellow Hearth | 63.9 | 5.5 | 39.0 | 28.8 | 45 | 49 | 6 | 0 | 0 | 0 |
| FA-HS1 | Two-Tone Red | 51.3 | 12.7 | 27.0 | 40.7 | 0 | 1 | 23 | 37 | 36 | 2 |
| FA-HS2 | Red Orange | 49.0 | 16.8 | 26.9 | 40.5 | 0 | 0 | 12 | 32 | 50 | 5 |
| FA-TR1 | Light Brown | 35.3 | 5.1 | 22.7 | 28.6 | 0 | 5 | 54 | 29 | 10 | 0 |
| FA-TR2 | Dark Brown | 30.8 | 7.1 | 21.8 | 23.5 | 0 | 1 | 54 | 34 | 11 | 0 |
| FA-TR3 | Ocher Light Brown | 43.3 | 6.9 | 29.9 | 30.8 | 0 | 2 | 48 | 35 | 15 | 0 |
| FA-TR4 | Ocher Brown | 38.0 | 8.9 | 31.3 | 25.4 | 0 | 0 | 33 | 46 | 20 | 0 |
| FA-TR5 | Ocher | 49.8 | 7.4 | 35.2 | 23.7 | 1 | 6 | 61 | 29 | 4 | 0 |
| FA-TR6 | Brick Red | 35.7 | 10.3 | 26.1 | 33.0 | 1 | 6 | 40 | 37 | 17 | 1 |
| FA-TR7 | Brown | 33.2 | 7.0 | 24.3 | 27.8 | 0 | 2 | 37 | 48 | 13 | 0 |
| UM-CO1 | Faded Light Pink | 52.9 | 13.6 | 19.3 | 19.8 | 0 | 24 | 55 | 16 | 4 | 0 |
| UM-CO2 | Pink | 46.9 | 14.0 | 12.1 | 18.5 | 1 | 26 | 63 | 9 | 1 | 0 |
| UM-CO3 | Rough Light Pink | 57.9 | 11.7 | 17.4 | 22.7 | 10 | 35 | 41 | 10 | 2 | 1 |
| UM-CO4 | Vivid Light Pink | 50.1 | 16.9 | 15.7 | 23.3 | 0 | 24 | 66 | 6 | 3 | 0 |
| UM-CO5 | Opaque Light Pink | 51.4 | 15.1 | 12.0 | 18.6 | 1 | 23 | 68 | 6 | 1 | 0 |
| Peak Position (cm−1) | Vibrational Mode | Biochemical Assignments |
|---|---|---|
| ~3260 | Stretching mode of OH | Water |
| ~2924, ~2855 | Symmetric and asymmetric stretching modes of CH2 moieties | Lipids alkyl chains, cellulose |
| ~1735 | Stretching mode of carbonyl moiety | Hemicellulose |
| ~1710 | Stretching mode of carboxylic moiety | Carboxylic (shoulder) |
| ~1660 | Stretching and bending modes of peptide linkage | Amide I of proteins |
| ~1545 | Bending mode of CH2 moieties | Amide II (shoulder) |
| ~1515 | C=C stretching vibrations | Phenolic acids |
| ~1440–1390 | C–H deformation vibration | Lipids and cellulose |
| ~1375–1345 | CH3 bending vibrations | Aliphatic groups |
| ~1340–1200 | In-phase combination of N–H deformation and C–N stretching vibrations | Amide III of proteins |
| ~1030 | C–O, C–N and C–C stretching vibrations | Sugar and proteins |
| ~920–700 | Vibrational modes of C–OH groups | Saccharides |
| ~708 | Bending mode of N–H bond | Amide V of proteins |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglioni, S.; Astolfi, P.; Conti, C.; Monaci, E.; Stefano, M.; Carloni, P. Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin. Molecules 2019, 24, 3974. https://doi.org/10.3390/molecules24213974
Castiglioni S, Astolfi P, Conti C, Monaci E, Stefano M, Carloni P. Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin. Molecules. 2019; 24(21):3974. https://doi.org/10.3390/molecules24213974
Chicago/Turabian StyleCastiglioni, Sara, Paola Astolfi, Carla Conti, Elga Monaci, Mariassunta Stefano, and Patricia Carloni. 2019. "Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin" Molecules 24, no. 21: 3974. https://doi.org/10.3390/molecules24213974
APA StyleCastiglioni, S., Astolfi, P., Conti, C., Monaci, E., Stefano, M., & Carloni, P. (2019). Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin. Molecules, 24(21), 3974. https://doi.org/10.3390/molecules24213974