
Role of L-Type Amino Acid Transporter 1 (LAT1) for the Selective Cytotoxicity of Sesamol in Human Melanoma Cells




Abstract
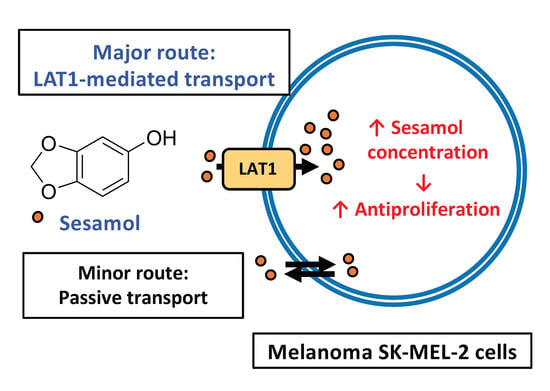
1. Introduction
2. Results
2.1. Sesamol Distribution into SK-MEL-2 and Vero Cells
2.2. Uptake Kinetics of Sesamol in SK-MEL-2 and Vero Cells
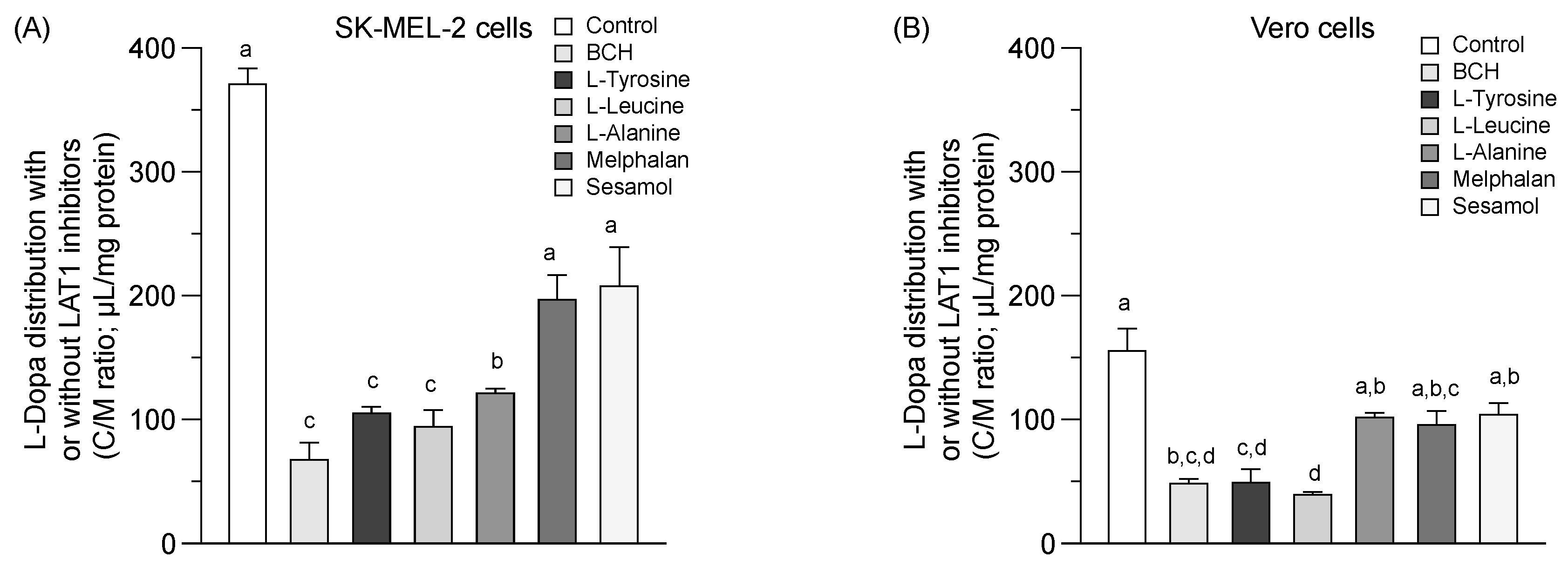
2.3. LAT1 Activity and Equilibrium Distribution of Sesamol
2.4. Inhibition of Sesamol Uptake Kinetics in SK-MEL-2 and Vero Cells
2.5. Anti-Proliferation in SK-MEL-2 and Vero Cells
2.6. mRNA Expression of LAT1 Protein in SK-MEL-2 and Vero Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Lines and Cell Culture
4.3. Cellular Uptake of Sesamol
4.3.1. Time-Course Study
4.3.2. Uptake Kinetics
4.3.3. LAT1 Function
4.3.4. Competitive Uptake between Sesamol and the other LAT1 Substrates
4.3.5. Uptake Kinetics of Sesamol via LAT1-Mediated Transport
4.4. HPLC Analysis
4.5. Antiproliferative Effect
4.6. mRNA Expression of LAT1 and LAT2
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krämer, S.D.; Aschmann, H.E.; Hatibovic, M.; Hermann, K.F.; Neuhaus, C.S.; Brunner, C.; Belli, S. When barriers ignore the “rule-of-five”. Adv. Drug Deliv. Rev. 2016, 101, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Artursson, P.; Avdeef, A.; Di, L.; Ecker, G.F.; Faller, B.; Houston, J.B.; Kansy, M.; Kerns, E.H.; Krämer, S.D.; et al. Passive lipoidal diffusion and carrier-mediated cell uptake are both important mechanisms of membrane permeation in drug disposition. Mol. Pharm. 2014, 11, 1727–1738. [Google Scholar] [CrossRef]
- Sugano, K.; Kansy, M.; Artursson, P.; Avdeef, A.; Bendels, S.; Di, L.; Ecker, G.F.; Faller, B.; Fischer, H.; Gerebtzoff, G. Coexistence of passive and carrier-mediated processes in drug transport. Nat. Rev. Drug Discov. 2010, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- The International Transporter Consortium. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.C.; Bode, B.P. Amino acid transporters ASCT2 and LAT1 in cancer: Partners in crime? Semin. Cancer Biol. 2005, 15, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, A.; Kaira, K.; Kato, M.; Yasuda, M.; Takahashi, A.; Tominaga, H.; Oriuchi, N.; Nagamori, S.; Kanai, Y.; Oyama, T.; et al. Prognostic significance of L-type amino acid transporter 1 (LAT1) expression in cutaneous melanoma. Melanoma Res. 2015, 25, 399–405. [Google Scholar] [CrossRef]
- Wang, Q.; Beaumont, K.A.; Otte, N.J.; Font, J.; Bailey, C.G.; van Geldermalsen, M.; Sharp, D.M.; Tiffen, J.C.; Ryan, R.M.; Jormakka, M.; et al. Targeting glutamine transport to suppress melanoma cell growth. Int. J. Cancer 2014, 135, 1060–1071. [Google Scholar] [CrossRef]
- Koch, A.; Lang, S.A.; Wild, P.J.; Gantner, S.; Mahli, A.; Spanier, G.; Berneburg, M.; Müller, M.; Bosserhoff, A.K.; Hellerbrand, C. Glucose transporter isoform 1 expression enhances metastasis of malignant melanoma cells. Oncotarget 2015, 6, 32748–32760. [Google Scholar] [CrossRef]
- Fukuda, Y.; Nagata, M.; Osawa, T.; Namiki, M. Chemical aspects of the antioxidative activity of roasted sesame seed oil, and the effect of using the oil for frying. Agric. Biol. Chem. 1986, 50, 857–862. [Google Scholar]
- Kato, M.J.; Chu, A.; Davin, L.B.; Lewis, N.G. Biosynthesis of antioxidant lignans in Sesamum indicum seeds. Phytochemistry 1998, 47, 583–591. [Google Scholar] [CrossRef]
- Baek, S.; Lee, S.-H. Sesamol decreases melanin biosynthesis in melanocyte cells and zebrafish: Possible involvement of MITF via the intracellular cAMP and p38/JNK signaling pathways. Exp. Dermatol. 2015, 24, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Mahendra Kumar, C.; Sathisha, U.V.; Dharmesh, S.; Rao, A.G.A.; Singh, S.A. Interaction of sesamol (3,4-methylenedioxyphenol) with tyrosinase and its effect on melanin synthesis. Biochimie 2011, 93, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Srisayam, M.; Weerapreeyakul, N.; Barusrux, S.; Tanthanuch, W.; Thumanu, K. Application of FTIR microspectroscopy for characterization of biomolecular changes in human melanoma cells treated by sesamol and kojic acid. J. Dermatol. Sci. 2014, 73, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Srisayam, M.; Weerapreeyakul, N.; Kanokmedhakul, K. Inhibition of two stages of melanin synthesis by sesamol, sesamin and sesamolin. Asian Pac. J. Trop. Biomed. 2017, 7, 886–895. [Google Scholar] [CrossRef]
- Wu, P.-Y.; You, Y.-J.; Liu, Y.-J.; Hou, C.-W.; Wu, C.-S.; Wen, K.-C.; Lin, C.-Y.; Chiang, H.-M. Sesamol inhibited melanogenesis by regulating melanin-related signal transduction in B16F10 cells. Int. J. Mol. Sci. 2018, 19, 1108. [Google Scholar] [CrossRef]
- Siriwarin, B.; Weerapreeyakul, N.; Tanthanuch, W.; Thumanu, K. Biomolecular changes and DNA targeting effect of sesamol in human lung adenocarcinoma (SK-LU-1) cells by FTIR microscopy. Asian Pac. J. Trop. Biomed. 2018, 8, 377. [Google Scholar]
- Siriwarin, B.; Weerapreeyakul, N. Sesamol induced apoptotic effect in lung adenocarcinoma cells through both intrinsic and extrinsic pathways. Chem. Biol. Interact. 2016, 254, 109–116. [Google Scholar] [CrossRef]
- Khamphio, M.; Barusrux, S.; Weerapreeyakul, N. Sesamol induces mitochondrial apoptosis pathway in HCT116 human colon cancer cells via pro-oxidant effect. Life Sci. 2016, 158, 46–56. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Sanyal, S.; Vaiphei, K.; Kakkar, V.; Kaur Deol, P.; Pal Kaur, I.; Kaur, T. Sesamol induces apoptosis by altering expression of Bcl-2 and Bax proteins and modifies skin tumor development in Balb/c mice. Anti Cancer Agents Med. Chem. 2017, 17, 726–733. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, B.; Wang, Y.; Zou, C.; Qiao, Q.; Diao, Z.; Mi, Y.; Zhu, D.; Liu, X. Sesamol induces human hepatocellular carcinoma cells apoptosis by impairing mitochondrial function and suppressing autophagy. Sci. Rep. 2017, 7, 45728. [Google Scholar] [CrossRef]
- Vivian, D.; Polli, J.E. Mechanistic interpretation of conventional Michaelis-Menten parameters in a transporter system. Eur. J. Pharm. Sci. 2014, 64, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 660. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational heterogeneity in cancer and the search for new cancer genes. Nature 2013, 499, 214. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sheikh, M.S. Melanoma: Molecular pathogenesis and therapeutic management. Mol. Cell. Pharmacol. 2014, 6, 228. [Google Scholar]
- Ribas, A.; Lawrence, D.; Atkinson, V.; Agarwal, S.; Miller, W.H.; Carlino, M.S.; Fisher, R.; Long, G.V.; Hodi, F.S.; Tsoi, J.; et al. Combined BRAF and MEK inhibition with PD-1 blockade immunotherapy in BRAF -mutant melanoma. Nat. Med. 2019, 25, 936. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Hanazono, K.; Fu, D.-R.; Endo, Y.; Kadosawa, T.; Iwano, H.; Uchide, T. A new treatment for human malignant melanoma targeting L-type amino acid transporter 1 (LAT1): A pilot study in a canine model. Biochem. Biophys. Res. Commun. 2013, 439, 103–108. [Google Scholar] [CrossRef]
- Yan, R.; Zhao, X.; Lei, J.; Zhou, Q. Structure of the human LAT1–4F2hc heteromeric amino acid transporter complex. Nature 2019, 568, 127. [Google Scholar] [CrossRef]
- Chien, H.-C.; Colas, C.; Finke, K.; Springer, S.; Stoner, L.; Zur, A.A.; Venteicher, B.; Campbell, J.; Hall, C.; Flint, A.; et al. Reevaluating the substrate specificity of the L-type amino acid transporter (LAT1). J. Med. Chem. 2018, 61, 7358–7373. [Google Scholar] [CrossRef]
- Zur, A.A.; Chien, H.-C.; Augustyn, E.; Flint, A.; Heeren, N.; Finke, K.; Hernandez, C.; Hansen, L.; Miller, S.; Lin, L.; et al. LAT1 activity of carboxylic acid bioisosteres: Evaluation of hydroxamic acids as substrates. Bioorg. Med. Chem. Lett. 2016, 26, 5000–5006. [Google Scholar] [CrossRef]
- Nagamori, S.; Wiriyasermkul, P.; Okuda, S.; Kojima, N.; Hari, Y.; Kiyonaka, S.; Mori, Y.; Tominaga, H.; Ohgaki, R.; Kanai, Y. Structure–activity relations of leucine derivatives reveal critical moieties for cellular uptake and activation of mTORC1-mediated signaling. Amino Acids 2016, 48, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Kärkkäinen, J.; Gynther, M.; Kokkola, T.; Petsalo, A.; Auriola, S.; Lahtela-Kakkonen, M.; Laine, K.; Rautio, J.; Huttunen, K.M. Structural properties for selective and efficient L-type amino acid transporter 1 (LAT1) mediated cellular uptake. Int. J. Pharm. 2018, 544, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Geier, E.G.; Schlessinger, A.; Fan, H.; Gable, J.E.; Irwin, J.J.; Sali, A.; Giacomini, K.M. Structure-based ligand discovery for the Large-neutral Amino Acid Transporter 1, LAT-1. Proc. Natl. Acad. Sci. USA 2013, 110, 5480–5485. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. In Chemical Biology: Method and Protocols, Methods in Molecular Biology; Hempel, J.E., Williams, C.H., Hong, C.C., Eds.; Springer New York: New York, NY, USA, 2015; Volume 1263, pp. 243–250. [Google Scholar]
- Del Amo, E.M.; Urtti, A.; Yliperttula, M. Pharmacokinetic role of L-type amino acid transporters LAT1 and LAT2. Eur. J. Pharm. Sci. 2008, 35, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Akanuma, S.; Yamakoshi, A.; Sugouchi, T.; Kubo, Y.; Hartz, A.M.S.; Bauer, B.; Hosoya, K. Role of L-type amino acid transporter 1 at the inner blood-retinal barrier in the blood-to-retina transport of gabapentin. Mol. Pharm. 2018, 15, 2327–2337. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, S.; Masuda, M.; Kim, S.-J.; Tanaka, Y.; Lee, K.-R.; Iwakado, S.; Nemoto, M.; Sasaki, S.; Shimono, K.; Tanaka, Y.; et al. The phenomenon of albumin-mediated hepatic uptake of organic anion transport polypeptide substrates: Prediction of the in vivo uptake clearance from the in vitro uptake by isolated hepatocytes using a facilitated-dissociation model. Drug Metab. Dispos. 2018, 46, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Soares-da-Silva, P.; Serrão, M.P. High- and low- affinity transport of L-leucine and L-dopa by the hetero amino acid exchangers LAT1 and LAT2 in LLC-PK 1 renal cells. Am. J. Physiol. Ren. Physiol. 2004, 287, F252–F261. [Google Scholar] [CrossRef]
- Srisongkram, T.; Weerapreeyakul, N. Validation of cell-based assay for quantification of sesamol uptake and its application for measuring target exposure. Molecules 2019, 24, 3522. [Google Scholar] [CrossRef]
Sample Availability: Not available. |
 ) and 37 °C (
) and 37 °C ( ) in (C) SK-MEL-2 and (D) Vero cells. Data points represent triplicates and are expressed as mean ± standard deviation. Different letters between bars indicate significant differences between samples (p < 0.05).
) and 37 °C () in (C) SK-MEL-2 and (D) Vero cells. Data points represent triplicates and are expressed as mean ± standard deviation. Different letters between bars indicate significant differences between samples (p < 0.05).
) in (C) SK-MEL-2 and (D) Vero cells. Data points represent triplicates and are expressed as mean ± standard deviation. Different letters between bars indicate significant differences between samples (p < 0.05).
) and 37 °C () in (C) SK-MEL-2 and (D) Vero cells. Data points represent triplicates and are expressed as mean ± standard deviation. Different letters between bars indicate significant differences between samples (p < 0.05).




 ) or without 1 mM BCH (
) or without 1 mM BCH ( ); and in Vero cells (---) with (▽) or without 1 mM BCH (△). (C) Percent cytotoxicity exhibited by BCH in both cells. Sigmoidal curved fitted to calculate the IC50. Data are expressed as mean ± standard deviation of three to six replicates.
) or without 1 mM BCH (); and in Vero cells (---) with (▽) or without 1 mM BCH (△). (C) Percent cytotoxicity exhibited by BCH in both cells. Sigmoidal curved fitted to calculate the IC50. Data are expressed as mean ± standard deviation of three to six replicates.
); and in Vero cells (---) with (▽) or without 1 mM BCH (△). (C) Percent cytotoxicity exhibited by BCH in both cells. Sigmoidal curved fitted to calculate the IC50. Data are expressed as mean ± standard deviation of three to six replicates.
) or without 1 mM BCH (); and in Vero cells (---) with (▽) or without 1 mM BCH (△). (C) Percent cytotoxicity exhibited by BCH in both cells. Sigmoidal curved fitted to calculate the IC50. Data are expressed as mean ± standard deviation of three to six replicates.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Sesamol Uptake Kinetic Parameters | |||||
|---|---|---|---|---|---|---|
| SK-MEL-2 | Vero | |||||
| Vmax (nmol/min/mg) | Km (µM) | Pd (µL/min/mg) | Vmax (nmol/min/mg) | Km (µM) | Pd (µL/min/mg) | |
| 37 °C | 4.2 ± 0.1a | 224.6 ± 19.3a | 2.3 ± 0.2b | 0.9 ± 0.0a | 159.9 ± 18.4a | 2.1 ± 0.1a |
| 4 °C | 0.3 ± 0.1b | 26.8 ± 9.8b | 2.7 ± 0.1a | 0.4 ± 0.1b | 66.1 ± 20.0b | 2.4 ± 0.3a |
| Compounds | Percent Inhibition of L-Dopa Distribution Volume (L-Dopa Distribution Volume; µL/mg Protein) | |
| SK-MEL-2 | Vero Cells | |
| Control | 0% (370.6 ± 13.0)a | 0% (155.6 ± 17.8)a |
| 1 mM BCH | 82% (67.3 ± 14.0)c | 69% (47.9 ± 3.7)b,c,d |
| 1 mM L-tyrosine | 72% (105.0 ± 5.2)c | 69% (48.7 ± 11.0)c,d |
| 2 mM L-leucine | 75% (94.1 ± 13.5)c | 75% (39.1 ± 2.3)d |
| 2 mM L-Alanine | 67% (121.2 ± 3.6)b | 35% (101.3 ± 4.0)a,b |
| 0.05 mM Melphalan | 47% (196.6 ± 20.3)a | 39% (95.5 ± 11.2)a,b,c |
| 1 mM Sesamol | 44% (207.6 ± 31.5)a | 33% (103.9 ± 9.2)a,b |
| Compounds | Percent Inhibition of Sesamol Distribution Volume (Sesamol Distribution Volume; µL/mg Protein) | |
| SK-MEL-2 | Vero Cells | |
| Control | 0% (23.2 ± 2.1)a | 0% (17.9 ± 1.7)a |
| 1 mM BCH | 55% (10.3 ± 0.9)c,d | 42% (10.4 ± 1.9)c |
| 1 mM L-tyrosine | 75% (5.8 ± 0.2)e | 41% (10.5 ± 0.7) c |
| 2 mM L-leucine | 50% (11.7 ± 0.7)c | 23% (13.8 ± 1.4)b |
| 2 mM L-Alanine | 44% (13.1 ± 0.3)b | 25% (13.3 ± 2.0)b,c |
| 0.05 mM Melphalan | 60% (9.4 ± 0.7)c,d | 35% (11.7 ± 0.7)b,c |
| 1 mM L-Dopa | 68% (7.4 ± 0.8)d | 40% (10.8 ± 2.0)b,c |
| Compounds | Sesamol Uptake Kinetic Parameters | |||||
|---|---|---|---|---|---|---|
| SK-MEL-2 | Vero | |||||
| Vmax (nmol/min/mg) | Km (µM) | Pd (µL/min/mg) | Vmax (nmol/min/mg) | Km (µM) | Pd (µL/min/mg) | |
| Sesamol | 4.2 ± 0.1a | 224.6 ± 19.3a | 2.3 ± 0.2b | 0.9 ± 0.0a | 159.9 ± 18.4a | 2.1 ± 0.1a |
| Sesamol + BCH | 0.2 ± 0.0d | 6.0 ± 1.1d | 0.5 ± 0.1d | 0.3 ± 0.1b | 38.2 ± 10.2b | 0.4 ± 0.2b |
| Sesamol + L-Leucine | 0.6 ± 0.2c | 62.1 ± 20.8b | 1.7 ± 0.2c | 0.4 ± 0.1b | 8.6 ± 2.4c | 2.5 ± 0.4a |
| Sesamol + L-Alanine | 1.4 ± 0.2b | 34.9 ± 3.5c | 2.9 ± 0.0a | 0.7 ± 0.0b | 5.6 ± 1.7c | 2.4 ± 0.0a |
| Compounds | IC50 (mM) | Selective Index | |
|---|---|---|---|
| SK-MEL-2 | Vero Cells | ||
| Sesamol | 2.0 ± 0.1b | 6.7 ± 0.2a | 3.4 |
| Sesamol + 1 mM BCH | 3.0 ± 0.0a | 4.8 ± 0.1b | 1.6 |
| Melphalan | 0.1 ± 0.0d | 0.6 ± 0.3b | 6.0 |
| Melphalan + 1 mM BCH | 0.2 ± 0.0c | 0.5 ± 0.1b | 2.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srisongkram, T.; Weerapreeyakul, N.; Kärkkäinen, J.; Rautio, J. Role of L-Type Amino Acid Transporter 1 (LAT1) for the Selective Cytotoxicity of Sesamol in Human Melanoma Cells. Molecules 2019, 24, 3869. https://doi.org/10.3390/molecules24213869
Srisongkram T, Weerapreeyakul N, Kärkkäinen J, Rautio J. Role of L-Type Amino Acid Transporter 1 (LAT1) for the Selective Cytotoxicity of Sesamol in Human Melanoma Cells. Molecules. 2019; 24(21):3869. https://doi.org/10.3390/molecules24213869
Chicago/Turabian StyleSrisongkram, Tarapong, Natthida Weerapreeyakul, Jussi Kärkkäinen, and Jarkko Rautio. 2019. "Role of L-Type Amino Acid Transporter 1 (LAT1) for the Selective Cytotoxicity of Sesamol in Human Melanoma Cells" Molecules 24, no. 21: 3869. https://doi.org/10.3390/molecules24213869
APA StyleSrisongkram, T., Weerapreeyakul, N., Kärkkäinen, J., & Rautio, J. (2019). Role of L-Type Amino Acid Transporter 1 (LAT1) for the Selective Cytotoxicity of Sesamol in Human Melanoma Cells. Molecules, 24(21), 3869. https://doi.org/10.3390/molecules24213869