Abstract
Polyphenols are categorized as plant secondary metabolites, and they have attracted much attention in relation to human health and the prevention of chronic diseases. In recent years, a considerable number of studies have been published concerning their physiological function in the digestive tract, such as their prebiotic properties and their modification of intestinal microbiota. It has also been suggested that several hydrolyzed and/or fission products, derived from the catabolism of polyphenols by intestinal bacteria, exert their physiological functions in target sites after transportation into the body. Thus, this review article focuses on the role of intestinal microbiota in the bioavailability and physiological function of dietary polyphenols. Monomeric polyphenols, such as flavonoids and oligomeric polyphenols, such as proanthocyanidins, are usually catabolized to chain fission products by intestinal bacteria in the colon. Gallic acid and ellagic acid derived from the hydrolysis of gallotannin, and ellagitannin are also subjected to intestinal catabolism. These catabolites may play a large role in the physiological functions of dietary polyphenols. They may also affect the microbiome, resulting in health promotion by the activation of short chain fatty acids (SCFA) excretion and intestinal immune function. The intestinal microbiota is a key factor in mediating the physiological functions of dietary polyphenols.
1. Introduction
1.1. Classification
Polyphenols are plant secondary metabolites ubiquitously present in many parts of the plant, including flowers, leaves, pulp, stems, and roots. They are not directly responsible for the growth and development of plants, but are necessary for the plant to survive in its environment. They are produced from primary metabolites and intermediates through unique biosynthetic pathways. In recent years, much attention has been paid to their potential role as functional food ingredients. A variety of polyphenols and their derivatives are currently expected to be used as dietary factors to prevent chronic diseases, such as diabetes, cancer, and stroke [1,2,3,4,5]. In 2013, Del Rio et al. [6] reviewed more than 500 publications focusing on the bioavailability of dietary polyphenols and their protective effects against chronic diseases.
Approximately 8000 compounds of polyphenols are found in the plant kingdom. These can be separated into high molecular weight tannins and low molecular weight polyphenols. Tannins consist of hydrolysable tannins (gallotannins and ellagitannins, Figure 1) and non-hydrolysable, condensed tannins (proanthocyanidins, Figure 2) [7]. Low molecular weight polyphenols can be categorized into several subgroups: phenolic acid derivatives, flavonoids, lignans, stilbenes, and curcumins. For many years, large numbers of studies have been undertaken to discover the physiological function of low molecular weight polyphenols, in particular, flavonoids; carboxylic acids, such as caffeic acid and ferulic acid; and resveratrol [8]. Flavonoids are typical low molecular weight polyphenols present in plant foods, and they are characterized by a coplanar diphenylpropane structure (Figure 3). Among them, catechins (flavan-3-ols) are also present as molecular units consisting of proanthocyanidins.
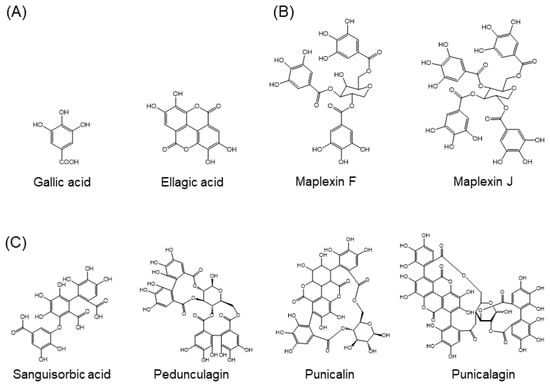
Figure 1.
Structures of hydrolysable tannins. (A) Molecules making up tannins, (B) examples of gallotannins, (C) examples of ellagitannins.
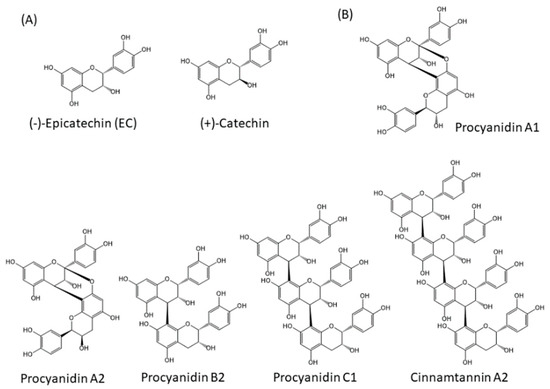
Figure 2.
Structures of non-hydrolysable condensed tannins (proanthocyanidins). (A) Molecules making up proanthocyanidins, (B) Examples of proanthocyanidins.
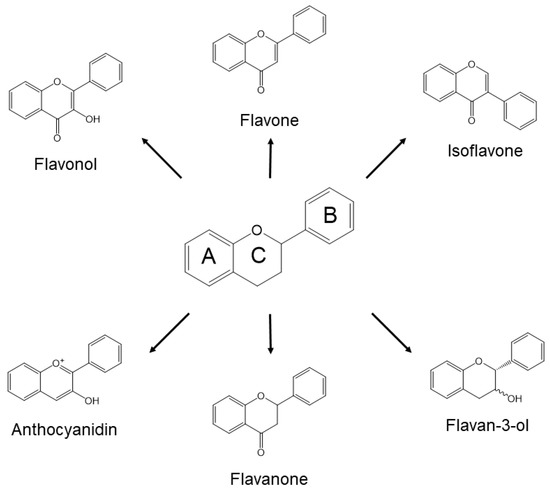
Figure 3.
Generic structures of major flavonoids.
1.2. Background of Polyphenol Research
In 1936, Bentsáth et al. [9] first reported that citrus flavonoids (hesperidin and rutin) reduced capillary fragility and permeability in human blood vessels. Thereafter, the vascular effects of plant-derived flavonoids were gradually explored [10]. In 1993, an epidemiological study in a Dutch population first revealed that the high intake of flavonoids (mainly flavonol-type flavonoids) correlates with a decrease in death due to atherosclerotic vascular disease [11]. This study encouraged researchers investigating dietary flavonoids and triggered human intervention studies of their bioavailability and bio-efficacy [12].
Polyphenols, including flavonoids, can act as antioxidants because they possess electron-donating phenolic groups in their structures. Therefore, several studies have investigated their antioxidant function in the prevention of oxidative stress-related cellular and extracellular damage. Antioxidant function has long been suggested to have a major role in promoting vascular health for the prevention of atherosclerosis [13]. Nevertheless, the bioavailability of flavonoids is significantly lower than the bioavailability of antioxidant vitamins and pro-vitamins (vitamin E, vitamin C, and carotenoids). Hollman et al. [14] suggested that antioxidant function may not explain the major vascular effects of dietary flavonoids. The cellular targets of flavonoids may be kinases involved in signal transduction pathways (Raf, Fyn, MEK, and PI3K) [15]. The resulting modulation of these pathways seems to be a major component of the physiological role of flavonoids after their absorption into the body [16]. They are able to act as modulators of cellular signaling pathways, provided that they are efficiently absorbed into the body and accumulate in target cells or target fluids [10,17]. The bioavailability of dietary flavonoids is, in general, very low. Therefore, they are mostly excreted into feces after transportation into the gut. Furthermore, high molecular weight tannins are scarcely absorbed in their original form because of their relatively large molecular size. Investigations into their physiological function has long been neglected or limited to their role in the digestive tract. The antioxidant effects of dietary polyphenols may only be exerted in the digestive tract, because of their high concentration in the gut. However, recent studies on the relationship between intestinal microbiota and dietary polyphenols have provoked new ideas on the physiological function of non-absorbable polyphenols, such as non-hydrolysable and hydrolysable tannins [18].
1.3. Interest in the Actions of Intestinal Microbiota
The gut microbiota is strongly associated with the occurrence of obesity by increasing the capacity for energy harvest [19]. It is, therefore, likely that an alteration of the intestinal microbiota may contribute to the improvement of human health. In fact, recent studies have demonstrated that non-absorbable polyphenols act in the intestine to alter the gut microbial community, resulting in lower systemic inflammation and improved metabolic outcomes [20,21,22]. Polyphenols may also exert beneficial effects on microbiota by acting as prebiotics [23].
In addition, a large new area of study involves investigating the effect of the gut microbiota on the brain and behavioral traits [24,25]. However, the association between behavior and the action of dietary polyphenols in the digestive tract is not well understood. On the other hand, the bioconversion of dietary polyphenols by gut microbiota has recently been shown to affect the nutritional phenotype of humans [26]. Enterobacteria-dependent catabolites of polyphenols may exert their effects in the body by transferring into the blood circulation. For example, urolithin or its glucuronide conjugate, an ellagitannin catabolized by intestinal bacteria, has been reported to increase muscle function in rodents [27], inhibit metastasis of cancer cell lines [28], and protect the impairment of cardiomyocytes [29]. Intestinal polyphenol catabolites may also exert their effects within the intestinal tract and intestinal walls. These may include inhibitory effects on colorectal cancer and suppressive effects on inflammatory bowel diseases [30,31,32,33].
The purpose of this review is to focus on the physiological effects of non-absorbable monomeric and polymeric polyphenols on the intestinal microbiota. Colonic bacteria-dependent catabolic pathways, bioavailability, and the function of their catabolites in the digestive tract and circulatory system is also discussed. Finally, we will discuss the mechanism by which plant polyphenols contribute to human health through the intestinal microbiota. In this paper, we will review flavonoids, tannins, curcumins, and resveratrol, but not isoflavones.
2. Hydrolysis and Absorption of Polyphenols in the Stomach and Small Intestine
2.1. Stability of Polyphenols in the Stomach
In general, low molecular weight polyphenols are partially absorbed into the body directly or after phase II enzyme-dependent metabolic conversion in small intestinal cells, as described above [34]. High molecular weight tannins and even low molecular weight polyphenols are transported into the large intestine in their original form. They are then excreted with feces without intestinal absorption, or they are catabolized by enterobacteria [35,36]. Therefore, the stomach is the minor location of absorption of polyphenols. In fact, low molecular weight polyphenols are stable, and glucose-bound polyphenols are only partially hydrolyzed, despite the low pH in the stomach [37]. It should be noted that anthocyanins (anthocyanidin glucosides) can be absorbed from the stomach via an active transport pathway involving bilitranslocase, an organic anion membrane carrier [38]. The stability of tannins in the stomach is similar to that of low molecular weight polyphenols [39]. However, the data for proanthocyanidins are more conflicting. An in vitro study suggests that procyanidins are hydrolyzed to bioavailable flavan-3-ol monomers [40], but an in vivo study claims a lack of depolymerization [41].
2.2. Intestinal Absorption of Sugar-Bound Polyphenols
In the small intestine, glucose-bound polyphenols are deglucosylated to aglycone by lactase-phlorizin hydrolase (LPH) or cytosolic β-glucosidase (CBG) of intestinal epithelial cells and then converted into O-glucuronide/O-sulfate conjugates by phase II enzymes in intestinal cells [34]. These conjugated metabolites are transported into the liver, where they are further metabolized, after which they either enter into the blood circulation or return to the digestive tract through the enterohepatic circulation (Figure 4). The total concentration of conjugated metabolites may reach micromolar levels within a few hours and then almost disappear after 24 h [42].
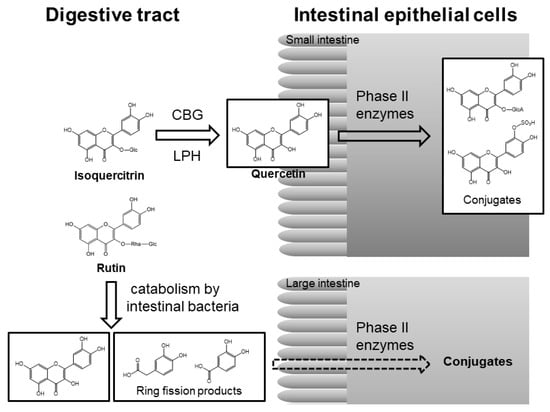
Figure 4.
Pathway of absorption and metabolism of isoquercitrin (quercetin 3-O-glucoside) and rutin (quercetin 3-O-β-rutinoside) in the digestive system [34,46,48]. CBG, cytosolic β-glucosidase; LPH, lactose phlorizin hydrolase; Glc, glucose; Rha, rhamnose; GlcA, glucuronic acid.
Other glycosides that are bound to sugars other than glucose are not substrates for these hydrolytic enzymes that release aglycone in the small intestine [34]. The time to reach Cmax (the compound’s maximum concentration in the blood) is significantly longer after rutin (quercetin-3-O-β-rutinoside) supplementation than after quercetin aglycone supplementation [43]. In contrast to quercetin aglycone and its glucosides, rutin is barely absorbed in the small intestine [44,45]. Most dietary rutin enters into the large intestine, where the colonic microbiota liberates quercetin aglycone from rutin. Quercetin aglycone can then be absorbed or further degraded to produce various ring-fission products by the action of enterobacteria (Figure 4) [46,47].
2.3. Intestinal Absorption of Monomeric Epicatechin and Its Related Monomeric Flavan-3-ols
In the case of flavan-3-ols, monomeric epicatechin is partly absorbed in the small intestine in its original form and is then subjected to glucuronidation, sulfation and/or O-methylation [49]. Natsume et al. [48] identified (−)-epicatechin-3′-O-glucuronide, 4′-O-methyl-(−)-epicatechin-3′-O-glucuronide, and 4′-O-methyl-(−)-epicatechin-5 or 7-O-glucuronide as metabolites of (−)-epicatechin in human urine. Ottaviani et al. [50] demonstrated that 82 ± 5% of ingested (−)-epicatechin is absorbed, and more than 20 different metabolites are present in human plasma after ingestion of [2-14C](−)-epicatechin. They also suggested that the gut microbiota is a key driver of (−)-epicatechin metabolism, because the concentration of its metabolites in plasma shows a biphasic pattern as a function of time. In the case of green tea flavan-3-ols, overall flavan-3-ol metabolite excretion is estimated to be 8.1% of intake [51]. Interestingly, un-metabolized (−)-epigallocatechin-3-gallate and (−)-epicatechin-3-gallate are also detected in the plasma, together with O-methylated, O-sulfated, and O-glucuronide conjugates of (−)-epicatechin and (−)-epigallocatechin [52].
2.4. Intestinal Absorption of Oligomeric Procyanidins
It is generally accepted that procyanidin polymers and oligomers with a degree of polymerization (DP) > 4 are not directly absorbed from the small intestine, although dimers and trimers can be detected in the plasma [53]. A-type procyanidin dimers, trimers, and tetramers can be transported across human intestinal epithelial Caco-2 cells [54]. Shoji et al. [55] found that apple procyanidins of each group, from dimers to pentamers, are present in rat plasma. Donovan et al. [39] claimed that neither monomers nor oligomers existed at detectable concentrations in vivo when grapeseed procyanidins were administered to rats. However, Serra et al. [56] reported free forms of dimers and trimers in rat plasma after oral intake of a grapeseed extract. The degree of polymerization has a major impact on the fate of procyanidins in the body, with greater degrees of polymerization showing poorer absorption through the gut barrier [57]. Baba et al. [58] found that the procyanidin dimer, B2 [epicatechin-(4-8)-epicatechin], is absorbed and a portion of B2 is degraded to monomeric epicatechin after B2 administration to rats. Interestingly, another study in rats showed that the procyanidin dimer, A1 [epicatechin-(2-O-7,4-8)-catechin] and the procyanidin dimer, A2 [epicatechin-(2-O-7, 4-8)-epicatechin] are better absorbed than the procyanidin dimer, B2 [epicatechin-(4-8)-epicatechin], although absorption of the A-type dimers was only 5–10% of monomeric epicatechin absorption [59].
2.5. Hydrolysis of Tannins in the Digestive Tract
A human pilot trial using gallotannin-rich mango pulp (Mangifera indica L. cv. Keitt) strongly suggested that gallotannins release free gallic acid in the gastrointestinal tract, indicating that gallotannins may serve as a pool of pro-gallic acid compounds that can be absorbed or can undergo microbial metabolism [60]. In humans, ellagic acid can be released by the hydrolysis of ellagitannins after the consumption of pomegranate juice. This is facilitated by physiological pH and/or gut microbiota actions [61]. A study of ileostomists consuming raspberries indicated that the hydrolysis of ellagitannins occurs in the stomach or small intestine [62]. Therefore, hydrolysable tannins can be hydrolyzed in the digestive tract before absorption or microbial catabolism.
3. Decomposition and Metabolism by Intestinal Bacteria
The gut microbiota influences polyphenol bioavailability by modifying the structure of aglycones, glycosides, and conjugates, such as O-glucuronides and O-sulfates. A large proportion of O-glycosides are converted into their corresponding aglycones by the acidic conditions of the stomach, by mucosal enzymes in the small intestine, or by enzymes in the gut microbiota. Aglycones are subjected to the activity of phase II enzymes during the absorption process and after their delivery to the liver, resulting in O-glucuronides and/or O-sulfates. These polyphenol conjugates are then excreted in the bile and re-enter the intestinal tract. In addition to enzymatic activity to deconjugate O-glycosides and O-glucuronides, gut microbes have catabolic capacity to perform carbon-carbon cleavage of heterocyclic and aromatic rings, dehydroxylation, decarboxylation, and hydrogenation of alkene moieties. These catabolic transformations follow a procedure of reactions that conform to the basic principles of chemistry. In this section, we outline the mechanism of the gut microbial transformation of major dietary polyphenols.
3.1. Flavonoid Quercetin
Quercetin, one of the major flavonols, is ubiquitously distributed in plants. It is mainly present as O-glycosides in various fruits and vegetables. The bacterial catabolites of rutin have been identified as 3,4-dihydroxyphenylacetic acid, 3-hydroxyphenylacetic acid, and 3-(3-hydroxyphenyl)propionic acid [44,47,63]. Several studies on the catabolism of quercetin or its glycosides by fecal microbiota have demonstrated an alternative conversion that generates 3,4-dihydroxybenzoic acid (i.e., protocatechuic acid) and 2,4,6-trihydroxybenzoic acid as the major catabolites (Figure 5) [64,65,66,67]. These catabolites are then further transformed into their O-methyl derivatives [68]. The enzymatic cleavage of quercetin to 3,4-dihydroxybenzoic acid is mediated by fungal and bacterial quercetin dioxygenases, known as quercetinases [69,70]. These enzymes contain divalent metal ions, such as iron, copper, or manganese, in their catalytic domain [71]. A reaction involving the chelation of oxygen atoms at positions 3 and 4 and the generation of superoxide or peroxyl radical species is proposed as the mechanism for producing 3,4-dihydroxybenzoic acid [69,72]. That is, quercetin degradation by quercetinase is initiated by chelation of the 4-keto/2,3-enol moiety with a divalent cation, resulting in the 3,4-diketo tautomer. The metal ion-bound flavonol complex places the oxygen atoms at position 2 via semi-quinone radicals, derived from phenoxy radicals. Arrangement of the endo-peroxide, based on the conversion of the 3-keto group into an acylium group, results in an ester, 2-protocatechuoyl-phloroglucinol carboxylic acid, linking the two aromatic rings and the release of carbon monoxide. While quercetin dioxygenases yield 2-protocatechuoyl-phloroglucinol carboxylic acid after incubation of quercetin with the microbiota, no catabolite of quercetin formed by hydrolytic cleavage of the ester, that is, 3,4-dihydroxybenzoic acid or 2,4,6-trihydroxybenzoic acid, is found in either animals or humans. On the other hand, there are no reports of 2,4,6-trihydroxybenzoic acid as a catabolite of quercetin. This may be because a large portion of 2,4,6-trihydroxybenzoic acid may be modified to phloroglucinol, as the known gut microbial catabolite of quercetin [73].
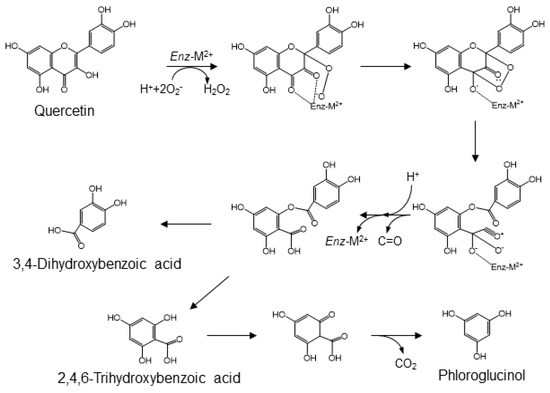
Figure 5.
A mechanism for quercetinase-mediated conversion of quercetin into protocatechuic acid, 2,4,6-trihydroxybenzoic acid and phloroglucinol [64,65,66,67].
Quercetinase- and other dioxygenase-mediated catabolic conversions depend on oxygen molecules. However, these microbial reactions can occur in the anoxic environment of the colon. The gastrointestinal mucosa separates the highly vascularized and oxygen-rich epithelium from the anoxic gut lumen, and the colon, which is relatively anoxic compared with the small intestine, is helpful for intestinal bacteria to form the more complex community [74,75]. Blood vessels in the sub-epithelial tissue of the intestine accelerate efficient nutrient uptake and supply oxygen to the epithelium and then, indirectly, to the gut lumen by diffusion. The resulting steep oxygen gradient across the epithelial cell layer affords stress on the epithelium and on the absolute anaerobic micro-organisms in the gut lumen. Facultative anaerobic micro-organisms may survive and thrive at this oxic/anoxic interface without interference from absolute anaerobes. Bacillus subtilis has been documented as an aerobic soil microbe that can be a facultative anaerobe [76]. Moreover, its isolation and behavior in the human intestinal tract has led to its recognition as a normal gut commensal bacterium in humans [77]. Quercetinase from B. subtilis has been overexpressed in E. coli, isolated, and biochemically characterized [70,78]. Based on these studies, B. subtilis is proposed to be a quercetinase-dependent producer of protocatechuic acid from dietary quercetin in the human gut. As a major gut microbial metabolite of quercetin, protocatechuic acid contributes to the health effects of fruits and vegetables rich in these flavonoids.
3.2. Proanthocyanidin
Dietary proanthocyanidins, which are formed by the condensation of single or multi-component of flavan-3-ols, are found in apples, chocolate, and grapes [79]. While monomeric, dimeric, and trimeric catechins are absorbed, to some extent, from the small intestine, larger oligomers have very poor bioavailability [80]. It has been demonstrated that proanthocyanidins undergo partial acid-catalyzed cleavage into monomeric flavan-3-ol units in the gastric environment [40]. The microbial catabolites of monomeric and oligomeric catechins in the large intestine appear to be 3-hydroxyphenylacetic acid, 3,4-dihydroxyphenylacetic acid, 3-(3-hydroxyphenyl)propionic acid, and 5-(3′-hydroxyphenyl)-γ-valerolactone [81]. The latter catabolite is the result of A-ring cleavages by two consecutive reverse Claisen reactions (Figure 6). Cleavage of the C-ring of a methylated quinone intermediate prepares the molecule for retro-Claisen cleavage. There are two possible mechanisms for the catabolism of catechin. One possible mechanism is a reduction of the quinone carbonyl, yielding the corresponding p-hydroxy catabolite, which is then subjected to dehydroxylation. The other one is that the carbonyl quinone hydride attacks at the methylated carbon, yielding 3,4-dihydroxyphenyl catabolites, which then eventually give rise to 5-(3′,4′-dihydroxyphenyl)-γ-valerolactone. The latter compound has been reported to be a product of gut microbial catabolism [82,83,84,85,86,87].
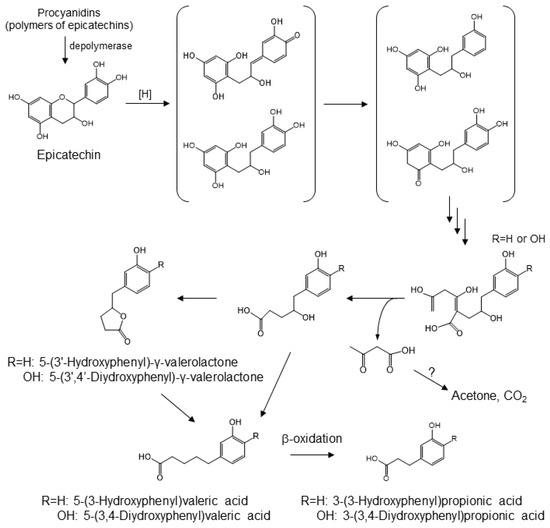
Figure 6.
Gut microbial conversion of procyanidin [81].
3.3. Anthocyanidins
Anthocyanidins, a subclass of flavonoid plant pigments, are responsible for red, purple, and blue colors of flower petals, vegetables, and fruits. Anthocyanins are the glycosylated forms of their corresponding aglycones. While the glycosides are bioavailable, they are susceptible to hydrolytic conversion into their corresponding anthocyanidins [88,89,90,91,92]. The glycosides and aglycones are both found as glucuronides in the urine of animals and humans [93,94]. The anthocyanidins are frequently metabolized into protocatechuic acid [95]. Using cyanidin as the prototypic anthocyanidin, its metabolism is initiated by cleavage of the heterocyclic flavylium ring at neutral or slightly basic conditions of the small intestine. Subsequent attack of the flavylium carbon at position 2 produces an unstable hemi-ketal that rapidly forms a ketone. Through keto-enol tautomerism of the neighboring enol functionality, the resulting α-diketo group is cleaved by gut microbiota to form protocatechuic acid and 2,4,6-trihydroxyphenylacetic acid (Figure 7). Although the cleavage reaction mechanism remains to be elucidated, it seems reasonable to propose that the C-C cleavage involves the attack of either carbonyl by a peroxyl anion species, similar to the initial step in the dioxygenase-mediated conversion of α-ketoglutarate into succinate [96]. Insertion of the resulting alkoxy oxygen between the original carbonyl carbons yields an anhydride, which forms two phenolic acids upon hydrolysis (Figure 7). This proposed mechanism is also similar to that of oxidative conversion of benzoins into benzoic acids [97]. After absorption from the intestinal tract and hepatic phase II metabolism, these phenolic acids are frequently detected in the urine, either in their original form or as their O-glucuronide or O-methyl derivatives [98,99,100].
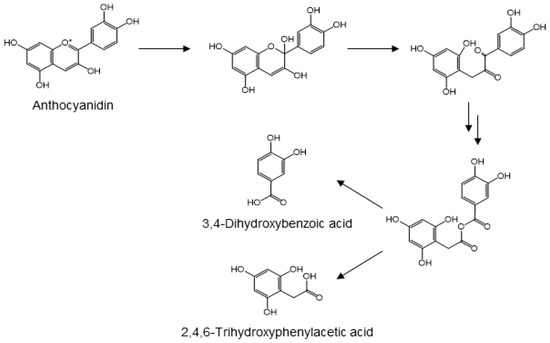
Figure 7.
Proposed mechanism for the microbial conversion of anthocyanidin into protocatechuic acid and 2,4,6-trihydroxyphenylacetic acid [96].
3.4. Curcumin
In animals and humans, curcumin, a yellow pigment from turmeric, is catabolized to its hydrogenated (dihydro-, tetrahydro-, hexahydro-, and octahydro-), desmethyl, O-glucuronide, and O-sulfate metabolites [101,102]. E. coli NADPH-dependent reductase, the enzyme responsible for the stepwise reduction of curcumin, has been isolated from human feces (Figure 8A) [103]. While the enzyme, NADPH-dependent curcumin/dihydrocurcumin reductase (CurA), catalyzes curcumin selectively, it does not reduce tetrahydrocurcumin to yield corresponding secondary alcohols. CurA has been identified as a member of the medium-chain dehydrogenase/reductase superfamily [103].
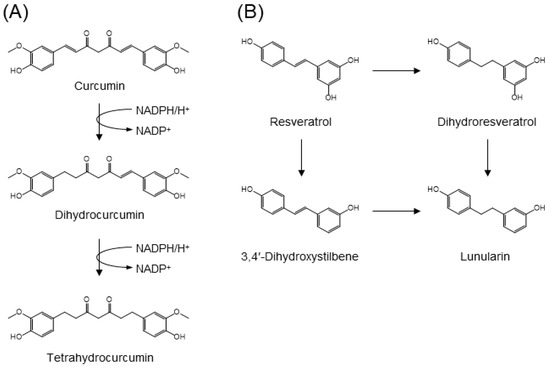
Figure 8.
Gut microbial metabolism of (A) curcumin by curcumin reductase CurA and (B) resveratrol into lunularin [101,102,104].
3.5. Resveratrol
Resveratrol is a stilbene-type polyphenol characteristically present in red wine and grapes. The main gut microbial catabolites of resveratrol are dihydroresveratrol and the m-deoxy metabolites of both resveratrol and dihydroresveratrol (Figure 8B) [104]. Using a panel of gut microbes, Slackia equolifaciens and Adlercreutzia equolifaciens were identified as dihydroresveratrol producers. Upon incubation of resveratrol with fecal samples, both dihydroresveratrol and 3,4′-dihydroxystilbene were observed as intermediates in the formation of lunularin [104].
3.6. Ellagitannin
Ellagitannins, including ellagic acid, punicalin and punicalagin, are known to be present in pomegranates, raspberries, strawberries, and walnuts [105,106]. Acid hydrolysis of ellagitannins releases free ellagic acid [107]. The process of gut microbial conversion of ellagic acid into urolithins can be explained by the carboxyl group-driven dehydroxylation of polyphenols. Strawberries, pomegranate juice, and walnuts are good sources of dietary ellagic acid, which is formed by C-C coupling of two molecules of gallic acid, followed by intramolecular condensation to form a di-lactone [108,109,110,111]. Urolithins are, arguably, the main urinary biomarkers of nut consumption [112]. In the catabolic pathway from ellagic acid to urolithins, one of the two lactone moieties undergo hydrolysis and the methylated quinone tautomer of the resulting carboxylic acid is then reduced in a manner similar to the ferredoxin-mediated reduction of 4-hydroxybenzoic acid (Figure 9). This results in a semi-hydroquinone, from which the p-hydroxy group leaves as a water molecule following decarboxylation. Subsequent dehydroxylation can occur via reduction of quinone-methide tautomers to form urolithins A, B, and C. In the microbial pathway from ellagic acid to urolithins, only the first dehydroxylation is driven by decarboxylation. Subsequent dehydroxylations involve a step-by-step reduction of a quinone, in which keto-enol tautomerism first produces a secondary alcohol and a subsequent hydride attack of the quinone leads to the dehydration step. Analogous dehydration is found in the dehydroxylation of catechin (Figure 6), which leads to the formation of 5-(3′-hydroxyphenyl)-γ-valerolactone. In addition, the pathway described for the dehydroxylation of 4-hydroxybenzoic acid may explain the formation of 3-hydroxybenzoic acid from protocatechuic acid after C-ring cleavage of cyanidin or quercetin (Figure 5).
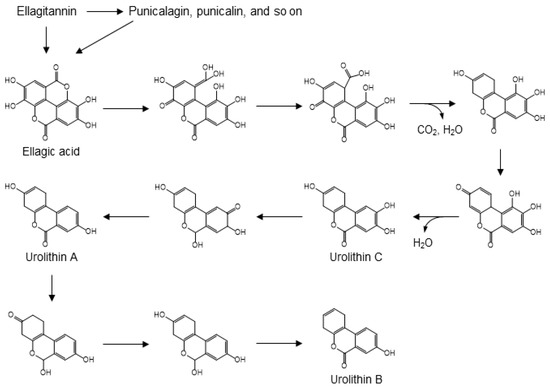
Figure 9.
Gut microbial conversion of ellagitannin into urolithins A, B, and C [108,109,110,111].
4. Physiological Functions Mediated by Intestinal Microbiota
4.1. Bioavailability and Action of Polyphenol Catabolites
The total intake of dietary polyphenol at the small intestine is estimated at around 10% [113,114,115]. Hence, a large proportion of ingested polyphenols are transported to the large intestine, where they are catabolized to phenolic acids by the intestinal bacteria. Recently, much attention has been paid to the bioavailability and physiological actions of their catabolites and to the functional interaction between polyphenols and intestinal bacteria (Figure 10).
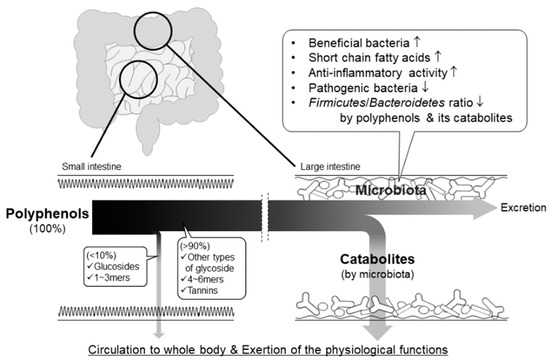
Figure 10.
Pathways of absorption, bioconversion, and physiological action of non-absorbable polyphenols in the intestinal tract.
Cyanidin glucosides are mainly converted into 3,4-dihydroxybenzoic acid in the human large intestine and are then absorbed into the bloodstream [116]. In the plasma, it is estimated that 44% and 0.02% of ingested cyanidin glucoside is in the form of 3,4-dihydroxybenzoic acid and cyanidin-3-glucoside, respectively [116], suggesting that 3,4-dihydroxybenzoic acid is a key compound for the vascular effects of cyanidin glucosides (Table 1). The intake of a cranberry juice cocktail containing mainly peonidin and cyanidin glycosides, hyperoside, and quercein, increases the plasma levels of these flavonoids and phenolic acids (Table 1) [117]. This study also showed a higher concentration of 3,4-dihydroxybenzoic acid than quercetin in human plasma. Plasma antioxidant capacity, estimated by the Oxygen Radical Absorption Capacity (ORAC) assay, correlates with the plasma levels of quercetin, epicatechin, 3,4-dihydroxybenzoic acid, 4-hydroxy-3-methoxy-benzoic acid, and 3,4-dihydroxyphenylacetic acid. 3,4-Dihydroxybenzoic acid exhibits the strongest correlation with plasma ORAC activity (r = 0.440) and also correlates with plasma TAP activity (r = 0.233) and protective activity against LDL oxidation (r = 0.503). Epicatechin conjugates, including O-sulfate and O-glucuronide, in human plasma after oral intake of 207 μmol of epicatechin, showed higher Cmax and faster Tmax than those of their conjugated catabolites (Table 1) [50]. However, the AUC for 0–24 h of the conjugated catabolites was higher than that of the epicatechin conjugates due to a longer elimination half-life. These pharmacokinetic data show that the increase in catabolites and their conjugates in plasma occurs around 7 h after detection of the parent flavonoids, due to colonic microbiota-dependent catabolism. Additionally, the bimodal peak of the original flavonoids suggests that they may be absorbed from the large intestine. A large proportion of ingested flavonoids seem to be transported into the large intestine, where they are degraded into their catabolites, including phenolic acids. The absorption level of the resulting catabolites is, consequently, high enough to exert greater physiological actions than the parent flavonoids. The combination of quercetin-3-glucoside or α-glucosyl rutin with fructooligosaccharide (FOS) increases the concentration of quercetin and O-methylated quercetin conjugates in rat plasma compared with the plasma concentrations in rats treated with quercetin-3-glucoside or α-glucosyl rutin alone [118,119]. This suggests that FOS may decrease the degradation of quercetin via the modification of the intestinal microbiota and therefore, increase the bioavailability of the parent compound, quercetin. Dietary fiber shows promise in modifying the bioavailability and physiological functions of polyphenols by changing components of the microbiota.

Table 1.
Pharmacokinetic properties of flavonoid catabolites in human trials.
The hydrolysable tannin, ellagitannin, is an oligomer composed of gallic acid, ellagic acid, and glucose. Ellagic acid is detected in human plasma 40 min after pomegranate intake [107,120]. However, plasma ellagic acid is undetectable within 5 h after the intake, suggesting it is rapidly cleared. Urolithins, which are well-known active catabolites of ellagic acid, and their conjugates can be detected in plasma at 0.5 and 6 h (0.04 and 0.11 μM of urolithin A respectively), with continuous excretion into the urine for 48 h after pomegranate consumption in some, but not all, volunteers [107,120]. Therefore, ellagic acid, produced by pH-dependent hydrolysis of ellagitannins [121], may be absorbed into the bloodstream at the upper digestive tract and may also undergo sustainable absorption as its bacterial catabolites, urolithins, at the colon. Since urolithin A and its conjugates are detected in some mouse tissues, including colon, prostate, liver, and kidney, at 1 to 6 h after oral administration of urolithin A [32], the continuous circulation of urolithins may allow their accumulation in tissues, thus facilitating their physiological functions.
The physiological functions of polyphenol catabolites, such as anti-oxidative, anti-inflammatory, and anti-proliferation activities, may differ from the functions of their parent polyphenols. Quercetin catabolites, such as 3,4-dihydroxybenzoic acid, 3-methyoxy-4-hydroxybenzoic acid, 3,4-dihydoxyphenylacetic acid, and 3-(3,4-dihydroxyphenyl)propionic acid, show one-half to one-eighth lower anti-oxidative activity than that of quercetin [122].
In an in vitro proximal colon model, green tea, black tea, and citrus flavonoids containing flavan-3-ols, theaflavin, quercetin, rutin, and hesperidin were catabolized to 4-hydroxyphenylacetic acid, 3-methoxy-4-hydroxyphenylacetic acid, 3,4-dihydroxyphenylacetic acid, and other catabolites. The anti-proliferation effect of these phenolic acids on colon cancer cell lines was one-fifth of the effect of the corresponding parent flavonoids [123]. A mixture of phenolic acids containing 3-(4-hydroxy-3-methoxyphenyl)propionic acid, 4-hydroxy-3-methoxybenzoic acid, 3,4-dihydroxybenzoic acid, and 3-(3-hydroxyphenyl)propanoic acid, which are detected in human plasma after blueberry juice consumption, have no effect on Nrf2 activation in human umbilical vein endothelial cells [124]. Furthermore, the inhibitory effect of phenolic acids, including 3-hydroxyphenylacetic acid, 3,4-dihydroxyphenylacetic acid, 3-phenylpropionic acid, 3-(3,4-dihydroxyphenyl)propionic acid, resorcinol, and phloroglucinol, on nitric oxide production in lipopolysaccharide-activated macrophages is minimal compared to the effects of their parent compound [125]. However, procyanidin A2 and its microbial catabolite, 3-(4-hydroxyphenyl)propionic acid, inhibit oxidized LDL-induced foam cell formation, inflammatory responses, oxidative stress, and the expression of cholesterol efflux/influx-related genes to the same degree, when applied at the same concentration [126]. Interestingly, urolithins show higher anti-oxidative, anti-inflammatory, and anti-proliferation activities than ellagitannin and ellagic acid [32,127,128]. Taken together with the pharmacokinetic data, bacterial catabolites can exert their specific functions in the large intestine and bloodstream, although the physiological function of polyphenols may be weakened or altered by the catabolism of parent compounds.
4.2. Action of Polyphenols on Gut Microbiota
Polyphenols are known to exhibit growth-promoting effects, namely prebiotic actions, on intestinal bacteria. This subject has recently been summarized in other review papers [129,130,131,132]. Although the molecular mechanisms of these prebiotic effects have not been fully elucidated, they may be associated, in part, with their selective antimicrobial activity against pathogenic bacteria. Phenolic extracts obtained from eight berries containing anthocyanins, flavan-3-ols, B-type proanthocyanidins, and ellagitannins inhibit the growth of food-poisoning bacteria, such as Salmonella Typhimurium, Listeria monocytogenes, and Staphylococcus aureus, but do not affect the growth of the probiotic bacterium, Lactobacillus rhamnosus [133]. Pomegranate ellagitannins strongly suppress the growth of Clostridium bacteria and S. aureus [134]. Lactic acid bacteria are also slightly reduced by treatment with ellagitannin, while the growth of bifidobacteria is markedly enhanced. However, in batch culture of human feces, pomegranate ellagitannin, but not punicalagin, increases the number of Bifidobacteria spp. and Lactobacillus-Enterococcus group bacteria after 5 and 10 h of inoculation, respectively, with no effect on Eubacterium rectale-Clostridium coccoides group and Clostridium histolyticum group bacteria [135]. Tannic acid from grape seeds and pomace show potent growth-promoting effects on Lactobacillus acidophilus [136]. Mango peels containing gallotannins also show prebiotic effects on bifidobacteria and lactic acid bacteria [18,137]. However, phenolic acids derived from the colonic bacteria-dependent catabolism of flavonoids have low selectivity against probiotic and pathogenic bacteria [138]. An intervention trial of 10 healthy adult men (45–50 years) investigated changes in the components of gut microbiota [139]. Drinking red wine containing anthocyanins, flavan-3-ols, B-type proanthocyanidins, flavonols, stilbenes, and phenolic acids for 20 days was shown to increase the proportions of the phyla Proteobacteria, Actinobacteria, Fusobacteria, Firmicutes, and Bacteroidetes in feces compared with baseline measurements. Meanwhile, de-alcoholized red wine with the same polyphenol composition only increased the proportions of Fusobacteria and Bacteroidetes and therefore, reduced the Firmicutes/Bacteroidetes ratio compared with subjects drinking red wine. Subjects drinking gin for the same period, as a non-polyphenol control group, showed a reduction in the proportions of these phyla. Curcumin and resveratrol also decreased the Firmicutes/Bacteroidetes ratio and have anti-inflammation and anti-carcinogenesis effects by modifying colonic microbial ecology in animal experiments [140,141,142,143,144]. Furthermore, resveratrol is thought to inhibit the production of trimethylamine and hence, trimethylamine oxide, from choline via the remodeling of the gut microbe composition [144]. Human microbiota-associated (HMA) animals (also known as human flora-associated, HFA) are germ-free rodents inoculated with a human fecal microbiota in order to study the human gut microbiome. Green tea polyphenols reduced blood glucose level, lipid metabolism biomarkers, and the Firmicutes/Bacteroidetes ratio in HFA mice fed a high-fat diet [145,146].
Akkermansia muciniphila is a gram-negative bacterium, isolated as a novel mucin-degrading bacterium from human feces [147]. Both live and pasteurized A. muciniphila are able to improve metabolic function in mice fed a high-fat diet [148]. This improvement is partly explained by the fact that an outer membrane protein, Amuc_1100, enhances glucose metabolism and gut barrier function by activating insulin and toll-like receptor 2 signaling [148]. The microbiome of mice on a high-fat diet show that an abundance of A. muciniphila is positively correlated with fatty acid oxidation and the browning of white adipocytes, but is negatively correlated with inflammation and metabolic syndrome makers [149]. This finding strongly suggests that A. muciniphila is a promising beneficial bacterium to prevent the development of metabolic syndrome. Interestingly, Colombian type 2 diabetes patients taking metformin have a higher abundance of A. muciniphila and SCFA-producing bacteria, such as Butyrivibri, Bifidobacteria, and Megasphaera, compared with those who do not take this drug [150]. Concord grape polyphenols, which appear to be composed of anthocyanidins, B-type proanthocyanidins, and flavan-3-ols, suppress inflammation in the intestinal tract and metabolic abnormalities and increase gut barrier integrity [20]. Cranberry extracts containing flavonols, anthocyanins, and proanthocyanidins also improve glucose and lipid homeostasis in mice fed high-fat and high-sucrose diets [22]. Oligomeric (monomer to tetramer) and polymeric (>pentamer) proanthocyanidins from apples suppress the abnormalities in both glucose and lipid homeostasis by modifying the expression of genes associated with inflammation, lipid metabolism, and gut epithelial tight junctions. However, there are some differences in efficiency between the oligomers and polymers [21]. More interestingly, only polymeric proanthocyanidins tend to decrease the Firmicutes/Bacteroidetes ratio and increase the relative abundance of A. muciniphila and Verrucomicrobia in the cecum of mice on a high-fat/high-sucrose diet. Although the role of A. muciniphila and Verrucomicrobia in the amelioration of metabolic abnormalities by these polyphenols needs to be fully investigated, the oligomeric and polymeric proanthocyanidins may exert these effects through a variety of molecular mechanisms.
Adhesion to the intestinal epithelia and subsequent colonization are thought to be important criteria for probiotic bacteria to exert their physiological effects on human health [151]. Epigallocatechin and procyanidin B1 and B2 increase the adhesion of lactic acid bacteria to intestinal epithelial cells in vitro, while some types of flavan-3-ols inhibit this adhesion [152]. Apple extracts containing procyanidin B2 and chlorogenic acid increase adhesion to intestinal epithelial cells in vitro. Quercetin and its glycosides show the most potent activity to enhance bacterial adhesion [153].
SCFA including acetate, propionate, and butyrate, are important metabolites from the action of colonic bacteria on indigestible carbohydrates. They play a role as physiological mediators by providing an energy source and acting as immunomodulators and metabolic regulators [154]. Rutin and some phenolic acids increase total SCFA levels and the ratio of acetate among SCFA in a multi-reactor gastrointestinal model [155]. The intake of a grapefruit extract, containing hesperidin and naringin, increases the wet weight of cecum digesta, and hence total SCFA content, in rats, while the combination of the extract with inulin weakened this effect [156]. Apple polyphenols containing epicatechin, procyanidins, and chlorogenic acid also increase rat cecum size and SCFA concentration in the cecum, and the combination of apple polyphenols with apple pectin shows an additive effect [157]. Inoculation with pomegranate ellagitannin increases the concentration of SCFAs in human fecal batch culture, while punicalagin has no such effect [135]. However, ellagitannin shows contradictory results for its combinational effect with FOS. Dietary FOS shows beneficial effects on pH, SCFA production, and bacterial enzymatic activity in the cecum. Different studies have shown that ellagitannin either interfered with or enhanced these positive effects of FOS [158,159]. Anthocyanins and phenolic acids may enhance SCFA production in the cecum of animals after FOS ingestion [118,159]. Flavonoids and phenolic acids, which promote the growth of SCFA-producing bacteria, including Bifidobacterium spp. and A. muciniphilla as mentioned above, may enhance the production of SCFAs. In addition, epigallocatechin-3-gallate, but not quercetin, stimulates the in vitro production of acetate and lactate by Bifidobacterium adolescentis, without growth-promoting effects [160], suggesting that flavonoids may be able to induce the production of SCFAs by colonic bacteria via activation of their metabolic function.
Culture supernatants of bifidobacteria and lactic acid bacteria are known to have anti-inflammatory activity [161,162,163], but both the active compounds and the mechanism of this action has not been fully demonstrated. When probiotic bacteria and polyphenols were anaerobically incubated for 3 h, a culture supernatant showed increased anti-inflammatory activity against lipopolysaccharide-stimulated macrophages [125]. Among 60 phytochemicals, quercetin, epigallocatechin-3-gallate, and some flavonoids increased the anti-inflammatory effects of B. adolescentis culture supernatant on nitric oxide production in activated macrophages in vitro [125,160]. On the other hand, these polyphenols were not able to potentiate the anti-inflammatory activity of lactic acid bacteria. Phenolic acids, which are assumed to be catabolites of these polyphenols, and the bifidobacteria metabolites, acetate and lactate, have no such effect [125,164], suggesting that these polyphenols may make B. adolescentis produce unknown active compounds. Interestingly, stearic acid has been tentatively identified as the anti-inflammatory compound obtained from the culture supernatant of quercetin-treated bifidobacteria [164]. Taken together, these findings suggest that polyphenols reaching the large intestine may not only be catabolized to small phenolic acids, but also elicit potentially beneficial effects of intestinal probiotic bacteria.
5. Future Directions
In recent years, metagenomic analysis of intestinal microbiota have provided a breakthrough in our understanding of the etiology of digestive diseases. Moreover, it is now recognized that changes in the intestinal microbiota affect the onset of not only digestive diseases, but also diabetes, atherosclerosis, neuropsychiatric diseases, and other systemic diseases [165]. However, the gut is an immune organ, in which more than half of all immune cells within the body are concentrated. The immune system in the gut affects the progression of obesity, diabetes, food allergies, and inflammatory bowel disease [166,167]. It is, therefore, of much interest to understand the relationship between the intestinal immune system and the intestinal microbiota. In this context, it is essential to focus research on elucidating the physiological function of polyphenols in the gut and their effects on the intestinal microbiota. In other words, microbiota should be taken into consideration when discussing the health effects of dietary polyphenols. It is, therefore, necessary to fully clarify the process of microbiota-dependent catabolism of dietary polyphenols in the large intestine. The effect of polyphenols on microbial behavior should also be evaluated in detail. As a whole, the relationship between dietary polyphenols and the intestinal microbiota is still obscure and unexplored. The intestinal microbiota is quite diverse among individuals within and between different ethnic groups. Therefore, the relationship between dietary polyphenols and microbiota is also diverse and profoundly complicated. To clarify the function of polyphenols in the absence of this confounding diversity, innovative in vitro experimental systems need to be established to replace rodent models, which have different microbiota to humans.
Finally, it should be noted that the role of microbiota in the brain–gut interaction is a future trend of polyphenol research. The intestinal microbiota is known to contribute to the development of reactions of the hypothalamic-pituitary-adrenal axis, which comprises major emotional reactions [25,168]. Microbiota is apparently recognized as a key factor in determining the stress responses and behavioral characteristics of the host. Successive intake of polyphenols may affect mental stress and behavior through the modification of intestinal microbiota. This is a subject on which future research should focus.
6. Conclusions
Polyphenols are composed of various subgroups whose basic structures differ for individual compounds. They can be divided into monomeric polyphenols and oligomeric tannins. After oral ingestion, some of the monomeric polyphenols and oligomeric tannins are absorbed, with or without deglucosylation, and enter into the blood circulation by crossing the epithelial cells in the small intestine. However, most polyphenols are transported into the large intestine, where intestinal bacteria catabolize them to chain fission products and/or monomeric catechins. Hydrolysable tannins, such as gallotannins and ellagitannins, generate gallic acid, ellagic acid, and their catabolites in the gut. Thereafter, these catabolites are excreted into the feces, but some of may be absorbed into the body through epithelial cells in the bowel. Polyphenols can be converted to various catabolites by the action of hydrolase and dioxygenase present in the microbiota. These catabolites may be non-negligible contributors to the health effects of dietary polyphenols. Furthermore, it is apparent that polyphenols, especially proanthocyanidins, have suppressive effects on the progress of life-style related diseases, such as diabetes, by modifying the patterns of the intestinal microbiota.
The intake of polyphenols improves the health effects of the intestinal microbiota by activating SCFA excretion, intestinal immune function, and other physiological processes. The microbiota-dependent effects of polyphenols may to be applied practically to the health food or supplement industries. For this purpose, further research is necessary to analyze the catabolic reactions of polyphenols and their reaction products, and to determine the mechanisms of action of these compounds on the intestinal microbiota.
Author Contributions
Conceptualization, J.T.; Writing—original draft preparation, K.K., Y.Y., and J.T.; writing—review and editing, K.K., Y.Y., and J.T.; Funding acquisition, J.T.
Funding
This work is supported by JSPS Kakenhi (Grant No. 16K00815 for J.T).
Conflicts of Interest
No potential conflicts were disclosed.
References
- Alkhalidy, H.; Wang, Y.; Liu, D. Dietary flavonoids in the prevention of T2D: An overview. Nutrients 2018, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Knaze, V.; Zamora-Ros, R. Polyphenols: Dietary assessment and role in the prevention of cancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 512–521. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Sheen, J.M.; Hu, W.L.; Hung, Y.C. Polyphenols and Oxidative Stress in Atherosclerosis-Related Ischemic Heart Disease and Stroke. Oxid. Med. Cell. Longev. 2017, 2017, 8526438. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Cheung, F.; Hong, M.; Feng, Y. The Potential and Action Mechanism of Polyphenols in the Treatment of Liver Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 8394818. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Phenols, polyphenols and tannins: An overview. In Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Crozier, A., Clifford, M.N., Ashihara, H., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 1–24. [Google Scholar] [CrossRef]
- Terao, J. Flavonoids: Metabolism bioavailability, and health impacts. In Phenolics and Human Health: Biochemistry, Nutrition, and Pharmacology; Fraga, C.G., Ed.; John Wiley & Sons: Hoboken NJ, USA, 2009; pp. 185–196. [Google Scholar] [CrossRef]
- Bentsáth, A.; Rusznyák, S.; Szent-Györgyi, A. Vitamin nature of flavones [1]. Nature 1936, 138, 798. [Google Scholar] [CrossRef]
- Almeida Rezende, B.; Pereira, A.C.; Cortes, S.F.; Lemos, V.S. Vascular effects of flavonoids. Curr. Med. Chem. 2016, 23, 87–102. [Google Scholar] [CrossRef]
- Hertog, M.G.L.; Feskens, E.J.M.; Kromhout, D.; Hertog, M.G.L.; Hollman, P.C.H.; Hertog, M.G.L.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Williamson, G.; Manach, C. Bioavailablity and bioefficacy of poly-phenols in humans, II: Review of 93 intervention studies. Am.J.Clin.Nutr. 2005, 81, 2435–2555. [Google Scholar] [CrossRef]
- Terao, J. Dietary flavonoids as antioxidants. Forum Nutr. 2009, 61, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.H.; Cassidy, A.; Comte, B.; Heinonen, M.; Richelle, M.; Richling, E.; Serafini, M.; Scalbert, A.; Sies, H.; Vidry, S. The Biological Relevance of Direct Antioxidant Effects of Polyphenols for Cardiovascular Health in Humans Is Not Established. J. Nutr. 2011, 141, 989S–1009S. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohnishi, K. Target molecules of food phytochemicals: Food science bound for the next dimension. Food Funct. 2012, 3, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Terao, J. Factors modulating bioavailability of quercetin-related flavonoids and the consequences of their vascular function. Biochem. Pharmacol. 2017, 139, 15–23. [Google Scholar] [CrossRef]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Roopchand, D.E.; Carmody, R.N.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.J.; Raskin, I. Dietary polyphenols promote growth of the gut bacterium akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, S.; Terao, A.; Yamamoto, Y.; Mukai, T.; Miura, T.; Shoji, T. Non-absorbable apple procyanidins prevent obesity associated with gut microbial and metabolomic changes. Sci. Rep. 2016, 6, 31208. [Google Scholar] [CrossRef]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Van Duynhoven, J.; Vaughan, E.E.; Jacobs, D.M.; Kemperman, R.A.; van Velzen, E.J.J.; Gross, G.; Roger, L.C.; Possemiers, S.; Smilde, A.K.; Dore, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-Dit-Félix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Shi, F.; Guo, Z.; Zhao, J.; Song, X.; Yang, H. Metabolite of ellagitannins, urolithin A induces autophagy and inhibits metastasis in human sw620 colorectal cancer cells. Mol. Carcinog. 2018, 57, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Savi, M.; Bocchi, L.; Bresciani, L.; Falco, A.; Quaini, F.; Mena, P.; Brighenti, F.; Crozier, A.; Stilli, D.; Del Rio, D. Trimethylamine-N-oxide (TMAO)-induced impairment of cardiomyocyte function and the protective role of urolithin B-glucuronide. Molecules 2018, 23, 549. [Google Scholar] [CrossRef]
- Kasimsetty, S.G.; Bialonska, D.; Reddy, M.K.; Thornton, C.; Willett, K.L.; Ferreira, D. Effects of pomegranate chemical constituents/intestinal microbial metabolites on CYP1B1 in 22Rv1 prostate cancer cells. J. Agric. Food Chem. 2009, 57, 10636–10644. [Google Scholar] [CrossRef]
- Miene, C.; Weise, A.; Glei, M. Impact of polyphenol metabolites produced by colonic microbiota on expression of COX-2 and GSTT2 in human colon cells (LT97). Nutr. Cancer 2011, 63, 653–662. [Google Scholar] [CrossRef]
- Surapaneni, K.M.; Priya, V.V.; Mallika, J. Pioglitazone, quercetin and hydroxy citric acid effect on cytochrome P450 2E1 (CYP2E1) enzyme levels in experimentally induced non alcoholic steatohepatitis (NASH). Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2736–2741. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases - Promises, Perspectives, and Pitfalls. Oxid. Med. Cell. Longev. 2016, 9346470. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Cassidy, A.; Minihane, A.M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Van Der Hooft, J.J.J.; Crozier, A. Human studies on the absorption, distribution, metabolism, and excretion of tea polyphenols1-3. Am. J. Clin. Nutr. 2013, 98, 1619S–1630S. [Google Scholar] [CrossRef] [PubMed]
- Pforte, H.; Hempel, J.; Jacobasch, G. Distribution pattern of a flavonoid extract in the gastrointestinal lumen and wall of rats. Nahrung 1999, 43, 205–208. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Mattivi, F. The interaction of anthocyanins with bilitranslocase. Biochem. Biophys. Res. Commun. 2002, 296, 631–636. [Google Scholar] [CrossRef]
- Donovan, J.L.; Lee, A.; Manach, C.; Rios, L.; Morand, C.; Scalbert, A.; Rémésy, C. Procyanidins are not bioavailable in rats fed a single meal containing a grapeseed extract or the procyanidin dimer B3. Br. J. Nutr. 2002, 87, 299–306. [Google Scholar] [CrossRef]
- Spencer, J.P.E.; Chaudry, F.; Pannala, A.S.; Srai, S.K.; Debnam, E.; Rice-Evans, C. Decomposition of cocoa procyanidins in the gastric milieu. Biochem. Biophys. Res. Commun. 2000, 272, 236–241. [Google Scholar] [CrossRef]
- Rios, L.Y.; Bennett, R.N.; Lazarus, S.A.; Rémésy, C.; Scalbert, A.; Williamson, G. Cocoa procyanidins are stable during gastric transit in humans. Am. J. Clin. Nutr. 2002, 76, 1106–1110. [Google Scholar] [CrossRef]
- Piskula, M.K.; Murota, K.; Terao, J. Bioavailability of flavonols and flavones. In Flavonoids and Related Compounds- Bioavailability and Function; Spencer, J.P., Crozier, A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 93–108. [Google Scholar]
- Erlund, I.; Kosonen, T.; Alfthan, G.; Mäenpää, J.; Perttunen, K.; Kenraali, J.; Parantainen, J.; Aro, A. Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers. Eur. J. Clin. Pharmacol. 2000, 56, 545–553. [Google Scholar] [CrossRef]
- Aura, A.M.; O’Leary, K.A.; Williamson, G.; Ojala, M.; Bailey, M.; Puupponen-Pimiä, R.; Nuutila, A.M.; Oksman-Caldentey, K.M.; Poutanen, K. Quercetin derivatives are deconjugated and converted to hydroxyphenylacetic acids but not methylated by human fecal flora in vitro. J. Agric. Food Chem. 2002, 50, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.C.P.; van Amelsvoort, J.M.M.; Katan, M.B. Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Jaganath, I.B.; Mullen, W.; Edwards, C.A.; Crozier, A. The relative contribution of the small and large intestine to the absorption and metabolism of rutin in man. Free Radic. Res. 2006, 40, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Jaganath, I.B.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. In vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites. Free Radic. Biol. Med. 2009, 47, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Natsume, M.; Osakabe, N.; Oyama, M.; Sasaki, M.; Baba, S.; Nakamura, Y.; Osawa, T.; Terao, J. Structures of (-)-epicatechin glucuronide identified from plasma and urine after oral ingestion of (-)-epicatechin: Differences between human and rat. Free Radic. Biol. Med. 2003, 34, 840–849. [Google Scholar] [CrossRef]
- Baba, S.; Osakabe, N.; Yasuda, A.; Natsume, M.; Takizawa, T.; Nakamura, T.; Terao, J. Bioavailability of (-)-epicatechin upon intake of chocolate and cocoa in human volunteers. Free Radic. Res. 2000, 33, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Borges, G.; Momma, T.Y.; Spencer, J.P.E.; Keen, C.L.; Crozier, A.; Schroeter, H. The metabolome of [2-14C](-)-epicatechin in humans: Implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives. Sci. Rep. 2016, 6, 29034. [Google Scholar] [CrossRef]
- Stalmach, A.; Mullen, W.; Steiling, H.; Williamson, G.; Lean, M.E.J.; Crozier, A. Absorption, metabolism, and excretion of green tea flavan-3-ols in humans with an ileostomy. Mol. Nutr. Food Res. 2010, 54, 323–334. [Google Scholar] [CrossRef]
- Stalmach, A.; Troufflard, S.; Serafini, M.; Crozier, A. Absorption, metabolism and excretion of Choladi green tea flavan-3-ols by humans. Mol. Nutr. Food Res. 2009, 53, 44–53. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Li, D.; Ho, C.T.; Li, J.; Wan, X. The absorption, distribution, metabolism and excretion of procyanidins. Food Funct. 2016, 7, 1273–1281. [Google Scholar] [CrossRef]
- Ou, K.; Percival, S.S.; Zou, T.; Khoo, C.; Gu, L. Transport of cranberry A-type procyanidin dimers, trimers, and tetramers across monolayers of human intestinal epithelial caco-2 cells. J. Agric. Food Chem. 2012, 60, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Masumoto, S.; Moriichi, N.; Akiyama, H.; Kanda, T.; Ohtake, Y.; Goda, Y. Apple procyanidin oligomers absorption in rats after oral administration: Analysis of procyanidins in plasma using the porter method and high-performance liquid chromatography/tandem mass spectrometry. J. Agric. Food Chem. 2006, 54, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; MacI, A.; Romero, M.P.; Valls, J.; Bladé, C.; Arola, L.; Motilva, M.J. Bioavailability of procyanidin dimers and trimers and matrix food effects in in vitro and in vivo models. Br. J. Nutr. 2010, 103, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, M.P.; Donovan, J.L.; Texier, O.; Felgines, C.; Remesy, C.; Scalbert, A. Metabolism of dietary procyanidins in rats. Free Radic. Biol. Med. 2003, 35, 837–844. [Google Scholar] [CrossRef]
- Baba, S.; Osakabe, N.; Natsume, M.; Terao, J. Absorption and urinary excretion of procyanidin B2 [epicatechin-(4β-8)-epicatechin] in rats. Free Radic. Biol. Med. 2002, 33, 142–148. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Vincken, J.-P.; Gruppen, H.; Hollman, P.C.H. Procyanidin Dimers A1, A2, and B2 Are Absorbed without Conjugation or Methylation from the Small Intestine of Rats. J. Nutr. 2009, 139, 1469–1473. [Google Scholar] [CrossRef]
- Barnes, R.C.; Krenek, K.A.; Meibohm, B.; Mertens-Talcott, S.U.; Talcott, S.T. Urinary metabolites from mango (Mangifera indica L. cv. Keitt) galloyl derivatives and in vitro hydrolysis of gallotannins in physiological conditions. Mol. Nutr. Food Res. 2016, 60, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Lee, R.; Heber, D. Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice. Clin. Chim. Acta 2004, 348, 63–68. [Google Scholar] [CrossRef]
- González-Barrio, R.; Borges, G.; Mullen, W.; Crozier, A. Bioavailability of anthocyanins and ellagitannins following consumption of raspberries by healthy humans and subjects with an ileostomy. J. Agric. Food Chem. 2010, 58, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic metabolism of dietary polyphenols: Influence of structure on microbial fermentation products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Serra, A.; MacIà, A.; Romero, M.P.; Reguant, J.; Ortega, N.; Motilva, M.J. Metabolic pathways of the colonic metabolism of flavonoids (flavonols, flavones and flavanones) and phenolic acids. Food Chem. 2012, 130, 383–393. [Google Scholar] [CrossRef]
- Hein, E.M.; Rose, K.; Van’t Slot, G.; Friedrich, A.W.; Humpf, H.U. Deconjugation and degradation of flavonol glycosides by pig cecal microbiota characterized by fluorescence in situ hybridization (FISH). J. Agric. Food Chem. 2008, 56, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhang, Z.; Zhang, N.; Liu, L.; Li, S.; Wei, H. In vitro catabolism of quercetin by human fecal bacteria and the antioxidant capacity of its catabolites. Food Nutr. Res. 2014, 58, 23406. [Google Scholar] [CrossRef]
- Schneider, H.; Simmering, R.; Hartmann, L.; Pforte, H.; Blaut, M. Degradation of quercetin-3-glucoside in gnotobiotic rats associated with human intestinal bacteria. J. Appl. Microbiol. 2000, 89, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, R.W.; George, S.E.; Claxton, L.D. Role of the gastrointestinal mucosa and microflora in the bioactivation of dietary and environmental mutagens or carcinogens. Drug Metab. Rev. 1992, 24, 425–492. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, K.; Fujita, Y. Excess production of Bacillus subtilis quercetin 2,3-dioxygenase affects cell viability in the presence of quercetin. Biosci. Biotechnol. Biochem. 2010, 74, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Schaab, M.R.; Barney, B.M.; Francisco, W.A. Kinetic and spectroscopic studies on the quercetin 2,3-dioxygenase from Bacillus subtilis. Biochemistry 2006, 45, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.J.; Huang, Q.Q.; Li, P.; Zhang, J.J. Catalytic dioxygenation of flavonol by MII-complexes (M = Mn, Fe, Co, Ni, Cu and Zn)-mimicking the MII-substituted quercetin 2,3-dioxygenase. Dalt. Trans. 2015, 44, 13926–13938. [Google Scholar] [CrossRef] [PubMed]
- Fetzner, S. Ring-Cleaving Dioxygenases with a Cupin Fold. Appl. Environ. Microbiol. 2012, 78, 2505–2514. [Google Scholar] [CrossRef]
- Ulbrich, K.; Reichardt, N.; Braune, A.; Krohd, L.W.; Blaut, M.; Rohn, S. The microbial degradation of onion flavonol glucosides and their roasting products by the human gut bacteria Eubacterium ramulus and Flavonifractor plautii. Food Res. Int. 2015, 67, 349–355. [Google Scholar] [CrossRef]
- Heinken, A.; Thiele, I. Anoxic Conditions Promote Species-Specific Mutualism between Gut Microbes In Silico. Appl Env. Microbiol 2015, 81, 4049–4061. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.T.; Colgan, S.P. Hypoxia and gastrointestinal disease. J. Mol. Med. 2007, 85, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Troup, B.; Szabo, A.; Hungerer, C.; Jahn, D. The anaerobic life of Bacillus subtilis: Cloning of the genes encoding the respiratory nitrate reductase system. FEMS Microbiol. Lett. 1995, 131, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.A.; Khaneja, R.; Tam, N.M.; Cazzato, A.; Tan, S.; Urdaci, M.; Brisson, A.; Gasbarrini, A.; Barnes, I.; Cutting, S.M. Bacillus subtilis isolated from the human gastrointestinal tract. Res. Microbiol. 2009, 160, 134–143. [Google Scholar] [CrossRef]
- Bowater, L.; Fairhurst, S.A.; Just, V.J.; Bornemann, S. Bacillus subtilis YxaG is a novel Fe-containing quercetin 2,3-dioxygenase. FEBS Lett. 2004, 557, 45–48. [Google Scholar] [CrossRef]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Gebhardt, S.; Prior, R.L. Concentrations of Proanthocyanidins in Common Foods and Estimations of Normal Consumption. J. Nutr. 2004, 134, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.E.; Frederiksen, H.; Krogholm, K.S.; Poulsen, L. Dietary proanthocyanidins: Occurrence, dietary intake, bioavailability, and protection against cardiovascular disease. Mol. Nutr. Food Res. 2005, 49, 159–174. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Vincken, J.P.; Aura, A.M.; Hollman, P.C.H.; Gruppen, H. Procyanidin dimers are metabolized by human microbiota with 2-(3,4-dihydroxyphenyl)acetic acid and 5-(3,4-dihydroxyphenyl)-γ-valerolactone as the major metabolites. J. Agric. Food Chem. 2009, 57, 1084–1092. [Google Scholar] [CrossRef]
- Barroso, E.; Sánchez-Patán, F.; Martín-Alvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V.; Peláez, C.; Requena, T.; Van De Wiele, T.; Martínez-Cuesta, M.C. Lactobacillus plantarum IFPL935 favors the initial metabolism of red wine polyphenols when added to a colonic microbiota. J. Agric. Food Chem. 2013, 61, 10163–10172. [Google Scholar] [CrossRef]
- Goodrich, K.M.; Neilson, A.P. Simultaneous UPLC-MS/MS analysis of native catechins and procyanidins and their microbial metabolites in intestinal contents and tissues of male Wistar Furth inbred rats. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 958, 63–74. [Google Scholar] [CrossRef]
- Jiménez-Girón, A.; Ibáñez, C.; Cifuentes, A.; Simó, C.; Muñoz-González, I.; Martín-Álvarez, P.J.; Bartolomé, B.; Victoria Moreno-Arribas, V. Faecal metabolomic fingerprint after moderate consumption of red wine by healthy subjects. J. Proteome Res. 2015, 14, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Margalef, M.; Pons, Z.; Muguerza, B.; Arola-Arnal, A. A rapid method to determine colonic microbial metabolites derived from grape flavanols in rat plasma by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2014, 62, 7698–7706. [Google Scholar] [CrossRef] [PubMed]
- Mülek, M.; Högger, P. Highly sensitive analysis of polyphenols and their metabolites in human blood cells using dispersive SPE extraction and LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 1885–1899. [Google Scholar] [CrossRef]
- Takagaki, A.; Nanjo, F. Catabolism of (+)-catechin and (-)-epicatechin by rat intestinal microbiota. J. Agric. Food Chem. 2013, 61, 4927–4935. [Google Scholar] [CrossRef] [PubMed]
- Bitsch, I.; Janssen, M.; Netzel, M.; Strass, G.; Frank, T. Bioavailability of anthocyanidin-3-glycosides following consumption of elderberry extract and blackcurrant juice. Int. J. Clin. Pharmacol. Ther. 2004, 42, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Bub, A.; Watzl, B.; Heeb, D.; Rechkemmer, G.; Briviba, K. Malvidin-3-glucoside bioavailability in humans after ingestion of red wine, dealcoholized red wine and red grape juice. Eur. J. Nutr. 2001, 40, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A13C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Felgines, C.; Verine, S.; Ra, T.; Gonthier, M.-P.; Texier, O.; Scalbert, A.; Lamaison, J.-L.; Ré Mé Sy, C. Strawberry anthocyanins are recovered in urine as glucuro- and sulfoconjugates in humans. J. Nutr. 2003, 133, 1296–1301. [Google Scholar] [CrossRef]
- Mertens-Talcott, S.U.; Rios, J.; Jilma-Stohlawetz, P.; Pacheco-Palencia, L.A.; Meibohm, B.; Talcott, S.T.; Derendorf, H. Pharmacokinetics of anthocyanins and antioxidant effects after the consumption of anthocyanin-rich açai juice and pulp (Euterpe oleracea Mart.) in human healthy volunteers. J. Agric. Food Chem. 2008, 56, 7796–7802. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, T.; Shida, Y.; Rahman, M.M.; Sekiya, M.; Hatano, Y.; Matsumoto, H.; Hirayama, M.; Konishi, T.; Ikeshiro, Y. Effect on both aglycone and sugar moiety towards Phase II metabolism of anthocyanins. Food Chem. 2008, 110, 493–500. [Google Scholar] [CrossRef]
- Kay, C.D.; Mazza, G.; Holub, B.J.; Wang, J. Anthocyanin metabolites in human urine and serum. Br. J. Nutr. 2004, 91, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.M.; Martin-Lopez, P.; O’Leary, K.A.; Williamson, G.; Oksman-Caldentey, K.M.; Poutanen, K.; Santos-Buelga, C. In vitro metabolism of anthocyanins by human gut microflora. Eur. J. Nutr. 2005, 44, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.B. Organic Chemistry of Enzyme-Catalyzed Reactions; Revised ed.; Academic Press: London, UK, 2002; pp. 227–250. ISBN 9780080513362. [Google Scholar]
- Kang, S.; Joo, C.; Kim, S.M.; Han, H.; Yang, J.W. Oxidation of benzoins to benzoic acids using sodium hydride under oxygen atmosphere. Tetrahedron Lett. 2011, 52, 502–504. [Google Scholar] [CrossRef]
- Amin, H.P.; Czank, C.; Raheem, S.; Zhang, Q.; Botting, N.P.; Cassidy, A.; Kay, C.D. Anthocyanins and their physiologically relevant metabolites alter the expression of IL-6 and VCAM-1 in CD40L and oxidized LDL challenged vascular endothelial cells. Mol. Nutr. Food Res. 2015, 59, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xia, M.; Yan, X.; Li, D.; Wang, L.; Xu, Y.; Jin, T.; Ling, W. Gut microbiota metabolism of anthocyanin promotes reverse cholesterol transport in mice via repressing miRNA-10b. Circ. Res. 2012, 111, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Woodward, G.M.; Needs, P.W.; Kay, C.D. Anthocyanin-derived phenolic acids form glucuronides following simulated gastrointestinal digestion and microsomal glucuronidation. Mol. Nutr. Food Res. 2011, 55, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Holder, G.M.; Plummer, J.L.; Ryan, A.J. The metabolism and excretion of curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) in the rat. Xenobiotica 1978, 8, 761–768. [Google Scholar] [CrossRef]
- Ireson, C.R.; Jones, D.J.L.; Orr, S.; Coughtrie, M.W.H.; Boocock, D.J.; Williams, M.L.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Metabolism of the Cancer Chemopreventive Agent Curcumin in Human and Rat Intestine. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 105–111. [Google Scholar] [PubMed]
- Hassaninasab, A.; Hashimoto, Y.; Tomita-Yokotani, K.; Kobayashi, M. Discovery of the curcumin metabolic pathway involving a unique enzyme in an intestinal microorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 6615–6620. [Google Scholar] [CrossRef]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.A.P.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef]
- Cerdá, B.; Tomás-Barberán, F.A.; Espín, J.C. Metabolism of antioxidant and chemopreventive ellagitannins from strawberries, raspberries, walnuts, and oak-aged wine in humans: Identification of biomarkers and individual variability. J. Agric. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef]
- González, C.A.; Pera, G.; Agudo, A.; Bueno-De-Mesquita, H.B.; Ceroti, M.; Boeing, H.; Schulz, M.; Del Giudice, G.; Plebani, M.; Carneiro, F.; et al. Fruit and vegetable intake and the risk of stomach and oesophagus adenocarcinoma in the European Prospective Investigation into Cancer and Nutrition (EPIC-EURGAST). Int. J. Cancer 2006, 118, 2559–2566. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Henning, S.M.; Zhang, Y.; Suchard, M.; Li, Z.; Heber, D. Pomegranate Juice Ellagitannin Metabolites Are Present in Human Plasma and Some Persist in Urine for Up to 48 Hours. J. Nutr. 2006, 136, 2481–2485. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Giménez-Bastida, J.A.; García-Conesa, M.T.; Gómez-Sánchez, M.B.; García-Talavera, N.V.; Gil-Izquierdo, A.; Sánchez-Álvarez, C.; Fontana-Compiano, L.O.; Morga-Egea, J.P.; Pastor-Quirante, F.A.; et al. Occurrence of urolithins, gut microbiota ellagic acid metabolites and proliferation markers expression response in the human prostate gland upon consumption of walnuts and pomegranate juice. Mol. Nutr. Food Res. 2010, 54, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Sánchez, M.A.; García-Villalba, R.; Monedero-Saiz, T.; García-Talavera, N.V.; Gómez-Sánchez, M.B.; Sánchez-Álvarez, C.; García-Albert, A.M.; Rodríguez-Gil, F.J.; Ruiz-Marín, M.; Pastor-Quirante, F.A.; et al. Targeted metabolic profiling of pomegranate polyphenols and urolithins in plasma, urine and colon tissues from colorectal cancer patients. Mol. Nutr. Food Res. 2014, 58, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; García-Villalba, R.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagic acid metabolism by human gut microbiota: Consistent observation of three urolithin phenotypes in intervention trials, independent of food source, age, and health status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Larrosa, M.; García-Conesa, M.T.; Cerdá, B.; Vidal-Guevara, M.L.; Tomás-Barberán, F.A.; Espín, J.C. Strawberry processing does not affect the production and urinary excretion of urolithins, ellagic acid metabolites, in humans. J. Agric. Food Chem. 2012, 60, 5749–5754. [Google Scholar] [CrossRef] [PubMed]
- Tulipani, S.; Urpi-Sarda, M.; García-Villalba, R.; Rabassa, M.; López-Uriarte, P.; Bulló, M.; Jáuregui, O.; Tomás-Barberán, F.; Salas-Salvadó, J.; Espín, J.C.; et al. Urolithins are the main urinary microbial-derived phenolic metabolites discriminating a moderate consumption of nuts in free-living subjects with diagnosed metabolic syndrome. J. Agric. Food Chem. 2012, 60, 8930–8940. [Google Scholar] [CrossRef]
- Hollman, P.C.H. Absorption, Bioavailability, and Metabolism of Flavonoids. Pharm. Biol. 2009, 42, 74–83. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Clifford, M.N. Diet-derived phenols in plasma and tissues and their implications for health. Planta Med. 2004, 70, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic Acid Is the Major Human Metabolite of Cyanidin-Glucosides. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.L.; Chen, C.Y.O.; Zampariello, C.A.; Blumberg, J.B. Flavonoids and phenolic acids from cranberry juice are bioavailable and bioactive in healthy older adults. Food Chem. 2015, 168, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Betalleluz-Pallardel, I.; Chirinos, R.; Aguilar-Galvez, A.; Noratto, G.; Pedreschi, R. Prebiotic effects of yacon (Smallanthus sonchifolius Poepp. & Endl), a source of fructooligosaccharides and phenolic compounds with antioxidant activity. Food Chem. 2012, 135, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, N.; Matsumoto, M.; Chiji, H.; Hara, H. Oligosaccharide promotes bioavailability of a water-soluble flavonoid glycoside, αG-rutin, in rats. J. Agric. Food Chem. 2009, 57, 1498–1505. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Zhang, Y.; McKeever, R.; Henning, S.M.; Lee, R.; Suchard, M.A.; Li, Z.; Chen, S.; Thames, G.; Zerlin, A.; et al. Pomegranate Juice and Extracts Provide Similar Levels of Plasma and Urinary Ellagitannin Metabolites in Human Subjects. J. Med. Food 2008, 11, 390–394. [Google Scholar] [CrossRef]
- Daniel, E.M.; Ratnayake, S.; Kinstle, T.; Stoner, G.D. The effects of pH and rat intestinal contents on the liberation of ellagic acid from purified and crude ellagitannins. J. Nat. Prod. 1991, 54, 946–952. [Google Scholar] [CrossRef]
- Dueñas, M.; Surco-Laos, F.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C. Antioxidant properties of major metabolites of quercetin. Eur. Food Res. Technol. 2011, 232, 103–111. [Google Scholar] [CrossRef]
- Has, A.; Activity, A.; Gao, K.; Xu, A.; Krul, C.; Venema, K.; Liu, Y.; Niu, Y.; Lu, J.; Bensoussan, L.; et al. Of the Major Phenolic Acids Formed during Human Microbial Fermentation of Tea, Citrus, and Soy Flavonoid Supplements, Only. Joournal Nutr. 2006, 136, 52–57. [Google Scholar] [CrossRef]
- Tang, J.S.; Vissers, M.C.M.; Anderson, R.F.; Sreebhavan, S.; Bozonet, S.M.; Scheepens, A.; Melton, L.D. Bioavailable Blueberry-Derived Phenolic Acids at Physiological Concentrations Enhance Nrf2-Regulated Antioxidant Responses in Human Vascular Endothelial Cells. Mol. Nutr. Food Res. 2018, 62, 1700647. [Google Scholar] [CrossRef]
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substance(s) by Bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. BioFactors 2013, 39, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Li, X.L.; Li, T.Y.; Li, M.Y.; Huang, R.M.; Li, W.; Yang, R.L. 3-(4-Hydroxyphenyl)propionic acid, a major microbial metabolite of procyanidin A2, shows similar suppression of macrophage foam cell formation as its parent molecule. RSC Adv. 2018, 8, 6242–6250. [Google Scholar] [CrossRef]
- Dobroslawa, B.; Kasimsetty, S.G.; Khan, S.I.; Daneel, F. Urolithins, intestinal microbial metabolites of pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Larrosa, M.; Toms-Barberán, F.A.; Dolara, P.; Espín, J.C. NF-κB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. Br. J. Nutr. 2010, 104, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of polyphenols and polyphenol-rich dietary sources on gut microbiota composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. Biomed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kähkönen, M.; Heinonen, M.; Määttä-Riihinen, K.; Oksman-Caldentey, K.M. Berry phenolics selectively inhibit the growth of intestinal pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Kasimsetty, S.G.; Schrader, K.K.; Ferreira, D. The effect of pomegranate (Punica granatum L.) byproducts and ellagitannins on the growth of human gut bacteria. J. Agric. Food Chem. 2009, 57, 8344–8349. [Google Scholar] [CrossRef]
- Bialonska, D.; Ramnani, P.; Kasimsetty, S.G.; Muntha, K.R.; Gibson, G.R.; Ferreira, D. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int. J. Food Microbiol. 2010, 140, 175–182. [Google Scholar] [CrossRef]
- Hervert-Hernández, D.; Pintado, C.; Rotger, R.; Goñi, I. Stimulatory role of grape pomace polyphenols on Lactobacillus acidophilus growth. Int. J. Food Microbiol. 2009, 136, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Sáyago-Ayerdi, S.G.; Zamora-Gasga, V.M.; Venema, K. Prebiotic effect of predigested mango peel on gut microbiota assessed in a dynamic in vitro model of the human colon (TIM-2). Food Res. Int. 2017, in press. [Google Scholar] [CrossRef]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Alvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Rivas, C.L.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Cerón, J.J.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Mcfadden, R.M.T.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The role of curcumin in modulating colonic microbiota during colitis and colon cancer prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Xiang, L.; Wang, Z.; Xiao, G.G.; Hu, J. Effect of curcumin on the diversity of gut microbiota in ovariectomized rats. Nutrients 2017, 9, 1146. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef]
- Chen, M.L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.D.; Zhang, Q.Y.; Mi, M.T. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. MBio 2016, 7, e02210-15. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, M.; Ho, C.T.; Guo, X.; Wu, Z.; Weng, P.; Yan, M.; Cao, J. Metagenomics analysis of gut microbiota modulatory effect of green tea polyphenols by high fat diet-induced obesity mice model. J. Funct. Foods 2018, 46, 268–277. [Google Scholar] [CrossRef]
- Wang, L.; Zeng, B.; Liu, Z.; Liao, Z.; Zhong, Q.; Gu, L.; Wei, H.; Fang, X. Green Tea Polyphenols Modulate Colonic Microbiota Diversity and Lipid Metabolism in High-Fat Diet Treated HFA Mice. J. Food Sci. 2018, 83, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia municiphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Everard, A.; Gómez-Valadés, A.G.; Matamoros, S.; Ramírez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed]
- De La Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin is associated with higher relative abundance of mucin-degrading Akkermansia muciniphila and several short-chain fatty acid-producing microbiota in the gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kailasapathy, K.; Chin, J. Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp. Immunol. Cell Biol. 2000, 78, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Bustos, I.; García-Cayuela, T.; Hernández-Ledesma, B.; Peláez, C.; Requena, T.; Martínez-Cuesta, M.C. Effect of flavan-3-ols on the adhesion of potential probiotic lactobacilli to intestinal cells. J. Agric. Food Chem. 2012, 60, 9082–9088. [Google Scholar] [CrossRef]
- Volstatova, T.; Marsik, P.; Rada, V.; Geigerova, M.; Havlik, J. Effect of apple extracts and selective polyphenols on the adhesion of potential probiotic strains of Lactobacillus gasseri R and Lactobacillus casei FMP. J. Funct. Foods 2017, 35, 391–397. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Sadeghi Ekbatan, S.; Sleno, L.; Sabally, K.; Khairallah, J.; Azadi, B.; Rodes, L.; Prakash, S.; Donnelly, D.J.; Kubow, S. Biotransformation of polyphenols in a dynamic multistage gastrointestinal model. Food Chem. 2016, 204, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Zduńczyk, Z.; Juśkiewicz, J.; Estrella, I. Cecal parameters of rats fed diets containing grapefruit polyphenols and inulin as single supplements or in a combination. Nutrition 2006, 22, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Aprikian, O.; Duclos, V.; Guyot, S.; Besson, C.; Manach, C.; Bernalier, A.; Morand, C.; Rémésy, C.; Demigné, C. Apple Pectin and a Polyphenol-Rich Apple Concentrate Are More Effective Together Than Separately on Cecal Fermentations and Plasma Lipids in Rats. J. Nutr. 2003, 133, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, B.; Milala, J.; Jurgoński, A.; Karlińska, E.; Zduńczyk, Z.; Juśkiewicz, J. Strawberry ellagitannins thwarted the positive effects of dietary fructooligosaccharides in rat cecum. J. Agric. Food Chem. 2014, 62, 5871–5880. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, B.; Juśkiewicz, J.; Jurgoński, A.; Kołodziejczyk, K.; Milala, J.; Kosmala, M.; Zduńczyk, Z. Anthocyanins in strawberry polyphenolic extract enhance the beneficial effects of diets with fructooligosaccharides in the rat cecal environment. PLoS ONE 2016, 11, e0149081. [Google Scholar] [CrossRef]
- Kawabata, K.; Kato, Y.; Sakano, T.; Baba, N.; Hagiwara, K.; Tamura, A.; Baba, S.; Natsume, M.; Ohigashi, H. Effects of phytochemicals on in vitro anti-inflammatory activity of Bifidobacterium adolescentis. Biosci. Biotechnol. Biochem. 2015, 79, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Candalh, C.; Bambou, J.C.; Terpend, K.; Cerf-Bensussan, N.; Heyman, M. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut 2004, 53, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Khokhlova, E.V.; Smeianov, V.V.; Efimov, B.A.; Kafarskaia, L.I.; Pavlova, S.I.; Shkoporov, A.N. Anti-inflammatory properties of intestinal Bifidobacterium strains isolated from healthy infants. Microbiol. Immunol. 2012, 56, 27–39. [Google Scholar] [CrossRef]
- Imaoka, A.; Shima, T.; Kato, K.; Mizuno, S.; Uehara, T.; Matsumoto, S.; Setoyana, H.; Hara, T.; Umesaki, Y. Anti-inflammatory activity of probiotic Bifidobacterium: Enhancement of IL-10 production in peripheral blood mononuclear cells from ulcerative colitis patients and inhibition of IL-8 secretion in HT-29 cells. World J. Gastroenterol. 2008, 14, 2511–2516. [Google Scholar] [CrossRef]
- Kawabata, K.; Baba, N.; Sakano, T.; Hamano, Y.; Taira, S.; Tamura, A.; Baba, S.; Natsume, M.; Ishii, T.; Murakami, S.; et al. Functional properties of anti-inflammatory substances from quercetin-treated Bifidobacterium adolescentis. Biosci. Biotechnol. Biochem. 2018, 82, 689–697. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Hirata, S.; Kunisawa, J. Gut microbiome, metabolome, and allergic diseases. Allergol. Int. 2017, 66, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Kunisawa, J.; Kiyono, H. Dietary and microbial metabolites in the regulation of host immunity. Front. Microbiol. 2017, 8, 2171. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).