
Cordycepin Isolated from Cordyceps militaris: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
2.1. Inhibitory Effects of Crude Extracts of C. militaris Fruiting body
2.2. Effects of Fractions from the EtOAc Extract of C. militaris Fruiting Body
2.3. Physiological Responses to the CM4 fraction, Cordycepin and Benzoic Acid
2.3.1. Chlorophyll and Carotenoid Contents
2.3.2. Electrolyte Leakage
2.3.3. Lipid Peroxidation
2.4. Biochemical Responses to the CM4 Fraction, Cordycepin and Benzoic acid
2.4.1. Total Phenolic Contents
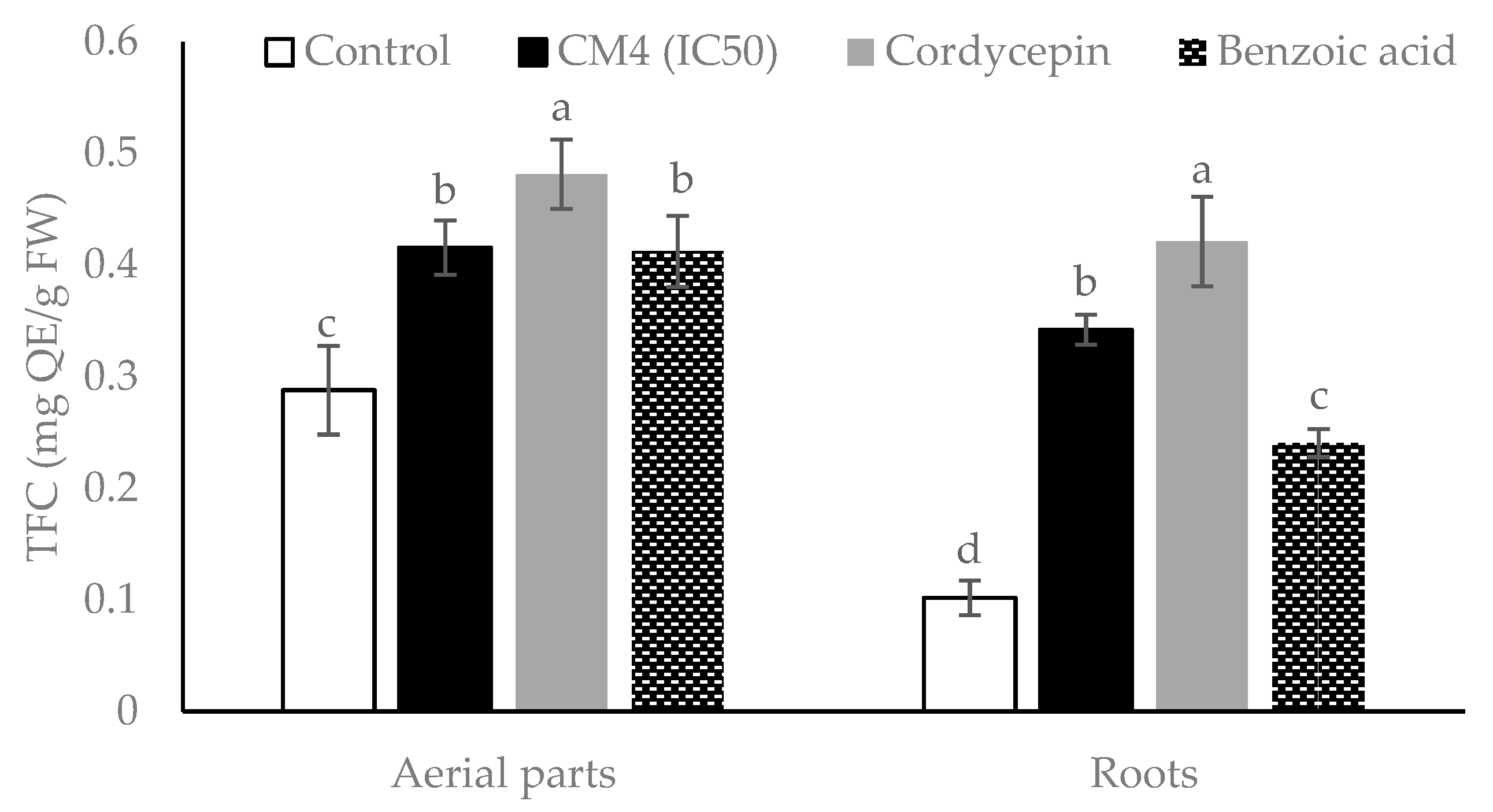
2.4.2. Total Flavonoid Contents
2.4.3. Proline Content
2.5. Compound Identification by GC-MS
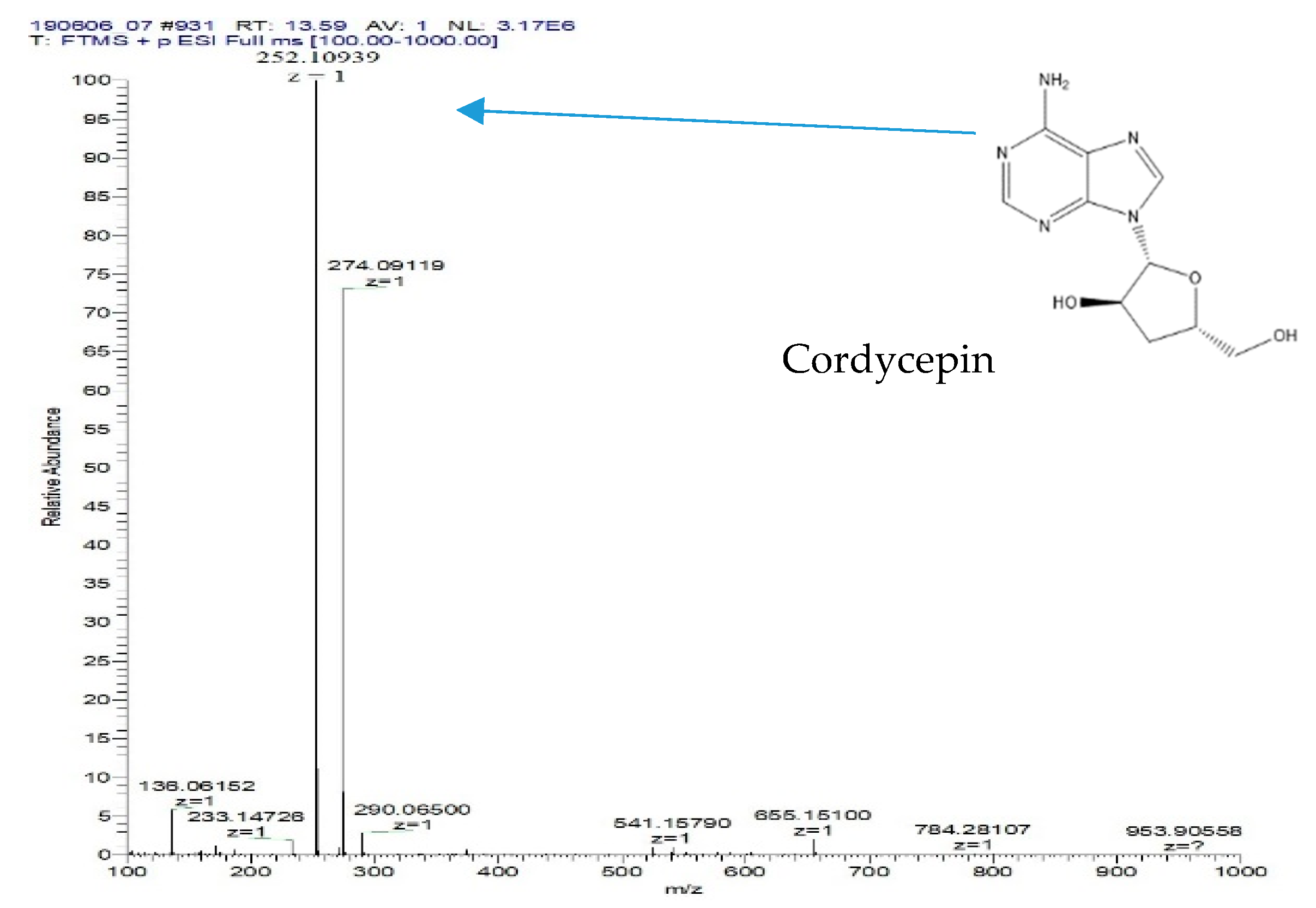
2.6. Cordycepin Detection and Quantification by High-Performance Liquid Chromatography (HPLC) and Liquid Chromatography-Electrospray Ionization-Mass Spectrometry (LC-ESI-MS) Analyses
2.7. Comparison of Cordycepin Yields in Different Extractions
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. C. militaris Materials
4.3. Preparation of C. militaris Extracts
4.4. Fractionation of Ethyl Acetate Extract
4.5. Germination and Growth Bioassays
4.6. Physiological Responses
4.6.1. Chlorophyll and Carotenoid Contents
4.6.2. Electrolyte Leakage
4.6.3. Lipid Peroxidation
4.7. Biochemical Responses
4.7.1. Quantification of Total Phenolic Contents
4.7.2. Quantification of Total Flavonoid Contents
4.7.3. Proline Content
4.8. Identification of Phytochemical Constituents by GC-MS and LC-ESI-MS
4.9. Quantification of Cordycepin by HPLC
4.10. Cordycepin Content in Different Extractions
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Xuan, T.D.; Tawata, S.; Khanh, T.D.; Chung, I.M. Decomposition of Allelopathic Plants in Soil. J. Agron. Crop. Sci. 2005, 191, 162–171. [Google Scholar] [CrossRef]
- Trezzi, M.M.; Vidal, R.A.; Junior, A.A.B.; Bittencourt, H.V.H.; Filho, A.P.D.S.S. Allelopathy: Driving mechanisms governing its activity in agriculture. J. Plant Interact. 2016, 11, 53–60. [Google Scholar] [CrossRef]
- Ladhari, A.; Omezzine, F.; Haouala, R. The impact of Tunisian Capparidaceae species on cytological, physiological and biochemical mechanisms in lettuce. S. Afr. J. Bot. 2014, 93, 222–230. [Google Scholar] [CrossRef]
- Omezzine, F.; Ladhari, A.; Haouala, R. Physiological and biochemical mechanisms of allelochemicals in aqueous extracts of diploid and mixoploid Trigonella foenum-graecum L. S. Afr. J. Bot. 2014, 93, 167–178. [Google Scholar] [CrossRef]
- Osivand, A.; Araya, H.; Appiah, K.S.; Mardani, H.; Ishizaki, T.; Fujii, Y. Allelopathy of Wild Mushrooms—An Important Factor for Assessing Forest Ecosystems in Japan. Forests 2018, 9, 773. [Google Scholar] [CrossRef]
- Idrees, H.; Javaid, A. Screening of some pathogenic fungi for their herbicidal potential against parthenium weed. Pak. J. Phytopathol. 2008, 20, 150–155. [Google Scholar]
- Vikrant, P.; Verma, K.K.; Rajak, R.C.; Pandey, A.K. Characterization of a Phytotoxin from Phoma herbarum for Management of Parthenium hysterophorus L. J. Phytopathol. 2006, 154, 461–468. [Google Scholar] [CrossRef]
- Brown, R.T. Influence of Naturally Occurring Compounds on Germination and Growth of Jack Pine. Ecology 1967, 48, 542–546. [Google Scholar] [CrossRef]
- Xuan, T.D.; Shinkichi, T.; Khanh, T.D.; Chung, I.M. Biological control of weeds and plant pathogens in paddy rice by exploiting plant allelopathy: An overview. Crop. Prot. 2005, 24, 197–206. [Google Scholar] [CrossRef]
- Khanh, T.D.; Cong, L.C.; Xuan, T.D.; Lee, S.J.; Kong, D.S.; Chung, I.M. Weed-Suppressing Potential of Dodder (Cuscuta hygrophilae) and its Phytotoxic Constituents. Weed Sci. 2008, 56, 119–127. [Google Scholar] [CrossRef]
- Dong, J.Z.; Wang, S.H.; Ai, X.R.; Yao, L.; Sun, Z.W.; Lei, C.; Wang, Y.; Wang, Q. Composition and characterization of cordyxanthins from Cordyceps militaris fruit bodies. J. Funct. Foods 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Koh, J.H.; Kim, K.M.; Kim, J.M.; Song, J.C.; Suh, H.J. Antifatigue and Antistress Effect of the Hot-Water Fraction from Mycelia of Cordyceps sinensis. Boil. Pharm. Bull. 2003, 26, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Feng, C.P.; Li, X.; Chang, M.C.; Meng, J.L.; Xu, L.J. Immunomodulatory and antioxidative activity of Cordyceps militaris polysaccharides in mice. Int. J. Boil. Macromol. 2016, 86, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Kang, I.C. The inhibitory effect of Cordycepin on the proliferation of cisplatin-resistant A549 lung cancer cells. Biochem. Biophys. Res. Commun. 2018, 498, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Quy, T.N.; Xuan, T.D. Xanthine Oxidase Inhibitory Potential, Antioxidant and Antibacterial Activities of Cordyceps militaris (L.) Link Fruiting Body. Medicines 2019, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Pan, M.C.; Chang, C.K.; Chang, S.W.; Hsieh, C.W. Optimization of Ultrasonic-Assisted Extraction of Cordycepin from Cordyceps militaris Using Orthogonal Experimental Design. Molecules 2014, 19, 20808–20820. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Burger, P.; Vogel, M.; Friese, K.; Bruning, A. The nucleoside antagonist cordycepin causes DNA double strand breaks in breast cancer cells. Investig. New Drugs 2012, 30, 1917–1925. [Google Scholar] [CrossRef]
- Wu, W.C.; Hsiao, J.R.; Lian, Y.Y.; Lin, C.Y.; Huang, B.M. The apoptoic effect of cordycepin on human OEC-M1 oral cancer line. Cancer Chemother. Pharmacol. 2007, 60, 103–111. [Google Scholar] [CrossRef]
- Sunaina; Singh, N. Alleviation of allelopathic stress of benzoic acid by indole acetic acid in Solanum lycopersicum. Sci. Hortic. 2015, 192, 211–217. [Google Scholar] [CrossRef]
- Bhowmik, P.C. Inderjit Challenges and opportunities in implementing allelopathy for natural weed management. Crop Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Yu, J.Q.; Ye, S.F.; Zhang, M.F.; Hu, W.H. Effects of root exudates and aqueous root extracts of cucumber (Cucumis sativus) and allelochemicals, on photosynthesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 2003, 31, 129–139. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, W.; Wang, X.; Yang, L.; Han, S.; Chen, S.; Strasser, R.J.; Valverde, B.E.; Qiang, S. Comparative phytotoxicity of usnic acid, salicylic acid, cinnamic acid and benzoic acid on photosynthetic apparatus of Chlamydomonas reinhardtii. Plant Physiol. Biochem. 2018, 128, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Kaushik, S. Cellular evidence of allelopathic interference of benzoic acid to mustard (Brassica juncea L.) seedling growth. Plant Physiol. Biochem. 2005, 43, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Andriana, Y.; Xuan, T.D.; Quan, N.V.; Quy, T.N. Allelopathic potential of Tridax procumbens L. on radish and identification of allelochemicals. Allelopath. J. 2018, 43, 223–238. [Google Scholar] [CrossRef]
- Tzvetkova, P.; Lyubenova, M.; Boteva, S.; Todorovska, E.; Tsonev, S.; Kalcheva, H. Effect of Herbicides Paraquat and Glyphosate on the Early Development of Two Tested Plants. IOP Conf. Ser. Earth Environ. Sci. 2019, 221, 1. [Google Scholar] [CrossRef]
- National Pesticide Information Center. Glyphosate, Technical Fact Sheet. Available online: http://npic.orst.edu/factsheets/archive/glyphotech.html (accessed on 9 July 2019).
- Xuan, T.D.; Tsuzuki, E.; Uematsu, H.; Terao, H. Weed control with alfalfa pellets in transplanting rice. Weed Boil. Manag. 2001, 1, 231–235. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research Progress on the use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Front. Plant Sci. 2015, 6, 347. [Google Scholar] [CrossRef] [PubMed]
- Van, T.M.; Xuan, T.D.; Minh, T.N.; Van Quan, N. Isolation and Purification of Potent Growth Inhibitors from Piper methysticum Root. Molecules 2018, 23, 1907. [Google Scholar] [CrossRef]
- Sodaeizadeh, H.; Rafieiolhossaini, M.; Van Damme, P. Herbicidal activity of a medicinal plant, Peganum harmala L., and decomposition dynamics of its phytotoxins in the soil. Ind. Crop. Prod. 2010, 31, 385–394. [Google Scholar] [CrossRef]
- Zhang, Z.; Tudi, T.; Liu, Y.; Zhou, S.; Feng, N.; Yang, Y.; Tang, C.; Tang, Q.; Zhang, J. Preparative isolation of cordycepin, N6-(2-hydroxyethyl)-adenosine and adenosine from Cordyceps militaris by macroporous resin and purification by recycling high-speed counter-current chromatography. J. Chromatogr. B 2016, 1033, 218–225. [Google Scholar] [CrossRef]
- Mokrani, A.; Madani, K. Effect of solvent, time and temperature on the extraction of phenolic compounds and antioxidant capacity of peach (Prunus persica L.) fruit. Sep. Purif. Technol. 2016, 162, 68–76. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A.L. Alleviation of salt stress-induced adverse effects on maize plants by exogenous application of indoleacetic acid (IAA) and inorganic nutrients—A field trial. Aust. J. Crop Sci. 2013, 7, 249–254. [Google Scholar]
- Bano, C.; Amist, N.; Sunaina; Singh, N. UV-B radiation escalate allelopathic effect of benzoic acid on Solanum lycopersicum L. Sci. Hortic. 2017, 220, 199–205. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Rana, N.; Kohli, R.K. Assessment of allelopathic interference of Chenopodium album through its leachates, debris extracts, rhizosphere and amended soil. Arch. Agron. Soil Sci. 2006, 52, 705–715. [Google Scholar] [CrossRef]
- Ben Jaballah, S.; Zribi, I.; Haouala, R. Physiological and biochemical responses of two lentil varieties to chickpea (Cicer arietinum L.) aqueous extracts. Sci. Hortic. 2017, 225, 74–80. [Google Scholar] [CrossRef]
- Chen, C.T.; Li, C.C.; Kao, C.H. Senescence of rice leaves XXXI. Changes of chlorophyll, protein, and polyamine contents and ethylene production during senescence of a chlorophyll-deficient mutant. J. Plant Growth Regul. 1991, 10, 201–205. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar]
- Ullah, F.; Ibrahim, M.; Ahmed, N.; Shinwari, Z.K.; Bano, A. Comparative impact of genetically modified and non modified maize (Zea mays L.) on succeeding crop and associated weed. Toxic. Ind. Health 2013, 32, 614–624. [Google Scholar]
- Amist, N.; Singh, N.B. Comparative Effects of Benzoic Acid and Water Stress on Wheat Seedlings. Russ. J. Plant Physiol. 2018, 65, 709–716. [Google Scholar] [CrossRef]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Belbachir, O.A.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2017, 218, 26–34. [Google Scholar] [CrossRef]
- Grzesiuk, A.; Dębski, H.; Oknińska, K.; Koczkodaj, D.; Szwed, M.; Horbowicz, M. Effect of root-zone glyphosate exposure on growth and anthocyanins content of radish seedlings. Acta Sci. Pol. Hortorum Cultus 2018, 17, 3–10. [Google Scholar] [CrossRef]
- Silva, F.B.; Costa, A.C.; Alves, R.R.P.; Megguer, C.A. Chlorophyll Fluorescence as an Indicator of Cellular Damage by Glyphosate Herbicide in Raphanus sativus L. Plants. Am. J. Plant Sci. 2014, 5, 2509–2519. [Google Scholar] [CrossRef]
- Sergiev, I.G.; Alexieva, V.S.; Ivanov, S.V.; Moskova, I.I.; Karanov, E.N. The phenylurea cytokinin 4PU-30 protects maize plants against glyphosate action. Pestic. Biochem. Physiol. 2006, 85, 139–146. [Google Scholar] [CrossRef]
- Huang, J.; Silva, E.N.; Shen, Z.; Jiang, B.; Lu, H. Effects of glyphosate on photosynthesis, chlorophyll fluorescence and physicochemical properties of cogongrass (Imperata cylindrical L.). Plant Omics 2012, 5, 177–183. [Google Scholar]
- Kang, N.; Lee, H.H.; Park, I.; Seo, Y.S. Development of High Cordycepin-Producing Cordyceps militaris Strains. Mycobiology 2017, 45, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Aramwit, P.; Porasuphatana, S.; Srichana, T.; Nakpheng, T. Toxicity evaluation of cordycepin and its delivery system for sustained in vitro anti-lung cancer activity. Nanoscale Res. Lett. 2015, 10, 138. [Google Scholar] [CrossRef]
- Huang, S.; Liu, H.; Sun, Y.; Chen, J.; Li, X.; Xu, J.; Hu, Y.; Li, Y.; Deng, Z.; Zhong, S. An effective and convenient synthesis of cordycepin from adenosine. Chem. Pap. 2017, 72, 149–160. [Google Scholar] [CrossRef]
- Farooq, M.; Ali, S.; Hameed, A.; Bharwana, S.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium stress in cotton seedlings: Physiological, photosynthesis and oxidative damages alleviated by glycinebetaine. S. Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Van Quan, N.; Xuan, T.D.; Tran, H.-D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Andriana, Y.; Tuyen, P.T. Antioxidant, α-Amylase and α-Glucosidase Inhibitory Activities and Potential Constituents of Canarium tramdenum Bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef]
- Rayee, R.; Tran, H.-D.; Xuan, T.D.; Khanh, T.D. Imposed Water Deficit after Anthesis for the Improvement of Macronutrients, Quality, Phytochemicals, and Antioxidants in Rice Grain. Sustainability 2018, 10, 4843. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Minh, T.N.; Xuan, T.D.; Tran, H.D.; Van, T.M.; Andriana, Y.; Khanh, T.D.; Van Quan, N.; Ahmad, A. Isolation and Purification of Bioactive Compounds from the Stem Bark of Jatropha podagrica. Molecules 2019, 24, 889. [Google Scholar] [CrossRef]
- Zhou, X.; Cai, G.; He, Y.I.; Tong, G. Separation of cordycepin from Cordyceps militaris fermentation supernatant using preparative HPLC and evaluation of its antibacterial activity as an NAD+-dependent DNA ligase inhibitor. Exp. Ther. Med. 2016, 12, 1812–1816. [Google Scholar] [CrossRef]
Sample Availability: Samples of C. militaris and purified cordycepin are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracting Solvents | IC50 (mg/mL) | ||
|---|---|---|---|
| Germination | Root | Shoot | |
| Hexane | 1.388 ± 0.170 a | 1.022 ± 0.132 a | 0.794 ± 0.043 a |
| Chloroform | 1.250 ± 0.126 a | 0.795 ± 0.033 b | 0.641 ± 0.436 b |
| EtOAc | 0.235 ± 0.030 c | 0.127 ± 0.008 d | 0.096 ± 0.006 d |
| Aqueous residue | 0.849 ± 0.082 b | 0.410 ± 0.042 c | 0.556 ± 0.029 c |
| Fractions | IC50 (mg/mL) | ||
|---|---|---|---|
| Germination | Root | Shoot | |
| CM1 | 2.865 ± 0.247 ab | 2.137 ± 0.100 a | 1.291 ± 0.183 b |
| CM2 | 2.438 ± 0.108 bc | 0.561 ± 0.213 b | 1.780 ± 0.350 a |
| CM3 | 2.090 ± 0.300 c | 0.467 ± 0.156 b | 1.682 ± 0.204 a |
| CM4 | 0.078 ± 0.013 e | 0.053 ± 0.004 c | 0.052 ± 0.015 e |
| CM5 | 1.402 ± 0.121 d | 0.382 ± 0.030 b | 0.611 ± 0.019 cd |
| CM6 | 1.442 ± 0.126 d | 0.368 ± 0.205 b | 0.862 ± 0.113 c |
| CM7 | 2.982 ± 0.449 a | 0.466 ± 0.175 b | 0.521 ± 0.092 d |
| CM8 | 2.463 ± 0.449 bc | 0.580 ± 0.076 b | 0.523 ± 0.165 d |
| Treatments | IC50 (mg/mL) | |||
|---|---|---|---|---|
| Germination | Root | Shoot | Reference | |
| CM4 | 0.078 ± 0.013 b | 0.053 ± 0.004 b | 0.052 ± 0.015 b | present study |
| Benzoic acid | 0.357 ± 0.052 a | 0.183 ± 0.017 a | 0.180 ± 0.004 a | present study |
| Cordycepin | 0.061 ± 0.001 b | 0.041 ± 0.003 b | 0.047 ± 0.004 b | present study |
| Glyphosate * | 0.226 | 0.137 | 0.161 | [25] |
| Compounds | Rt (min) | PA (%) | Chemical Formula | MW (g/mol) | Chemical Class | Quantity (g) |
|---|---|---|---|---|---|---|
| 1,6-Anhydro-beta-d-glucopyranose | 11.76 | 0.54 | C6H10O5 | 162,14 | Anhydrohexose | — |
| Pentadecanal | 14.52 | 19.79 | C15H30O | 226.40 | Fatty aldehyde | — |
| Cordycepin | 21.98 | 55.38 | C10H13N5O3 | 251,24 | Nucleosides | 0.226 |
| Code | Fungus Part | Extraction Methods | Cordycepin (mg/g DW) | |||
|---|---|---|---|---|---|---|
| Solvents | Extraction Times | Temperatures | Ultrasonic | |||
| A | Fruiting body | Methanol | 2 weeks | Ambient conditions | — | 6.166 ± 0.021a |
| B | Fruiting body | Water | 30 min | 100 °C | — | 3.548 ± 0.012c |
| C | Fruiting body | Water | 30 min | 70 °C | 40 KHz, 30 min | 4.248 ± 0.027b |
| Code | Fungus Part | Extraction Methods | |||
|---|---|---|---|---|---|
| Solvents | Extraction Time | Temperature | Ultrasonic | ||
| A | Fruiting body | Methanol | 2 weeks | Ambient condition | — |
| B | Fruiting body | Water | 30 min | 100 °C | — |
| C | Fruiting body | Water | 30 min | 70 °C | 40 Hz, 30 min |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.-D.; Khanh, T.D.; Teschke, R. Cordycepin Isolated from Cordyceps militaris: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate. Molecules 2019, 24, 2901. https://doi.org/10.3390/molecules24162901
Quy TN, Xuan TD, Andriana Y, Tran H-D, Khanh TD, Teschke R. Cordycepin Isolated from Cordyceps militaris: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate. Molecules. 2019; 24(16):2901. https://doi.org/10.3390/molecules24162901
Chicago/Turabian StyleQuy, Tran Ngoc, Tran Dang Xuan, Yusuf Andriana, Hoang-Dung Tran, Tran Dang Khanh, and Rolf Teschke. 2019. "Cordycepin Isolated from Cordyceps militaris: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate" Molecules 24, no. 16: 2901. https://doi.org/10.3390/molecules24162901
APA StyleQuy, T. N., Xuan, T. D., Andriana, Y., Tran, H.-D., Khanh, T. D., & Teschke, R. (2019). Cordycepin Isolated from Cordyceps militaris: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate. Molecules, 24(16), 2901. https://doi.org/10.3390/molecules24162901