
Gingers and Their Purified Components as Cancer Chemopreventative Agents


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. True Ginger
2.1. Anti-Inflammatory Studies
2.2. Animal and In Vitro Studies with Ginger Extract
2.3. True Ginger Extract and Helicobacter
2.4. Human Chemopreventive Efficacy Studies with True Ginger Extract
2.5. True Ginger Rhizome Bioactive Constituents
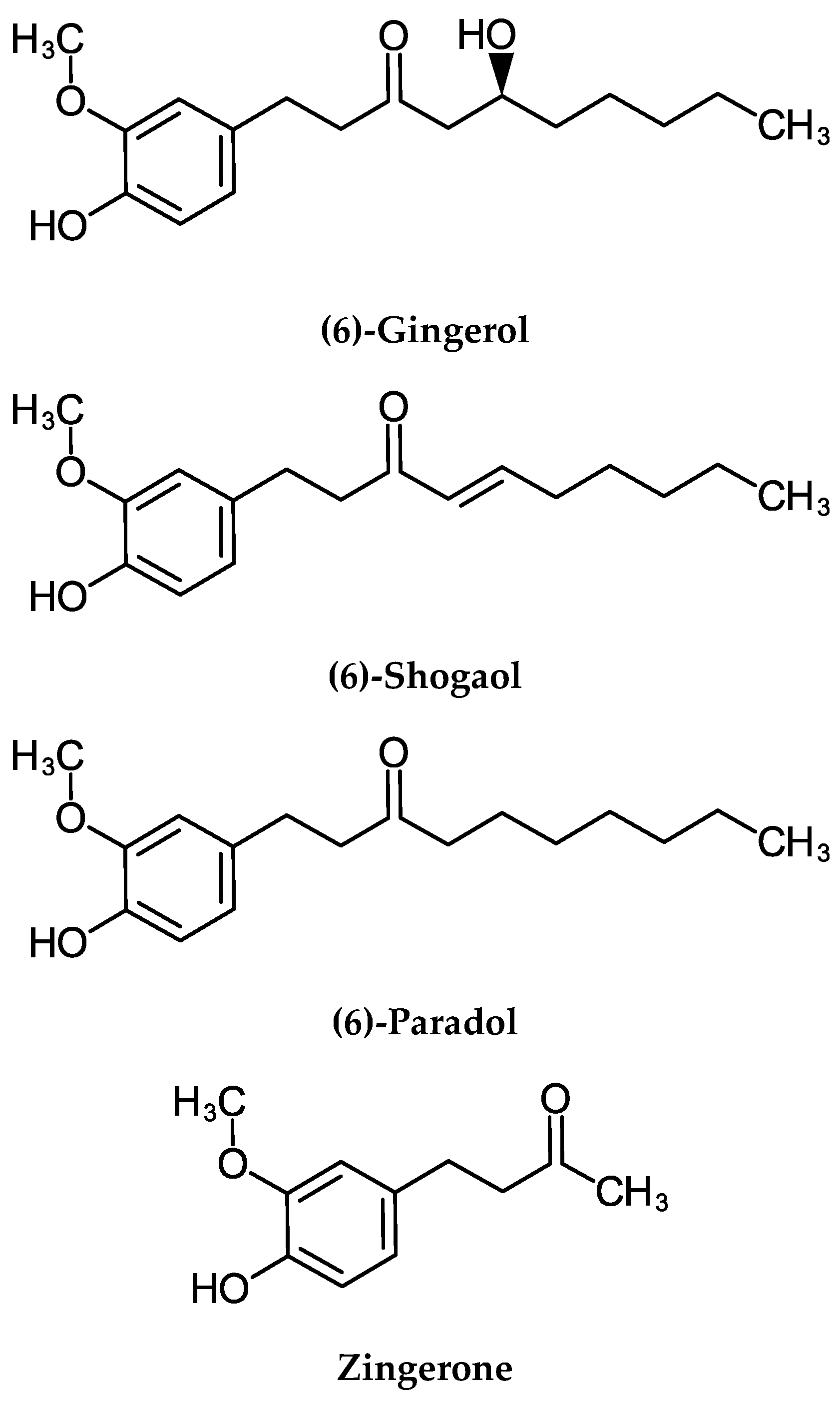
2.5.1. Gingerol
Metabolism of Gingerol
Molecular Activities of Gingerols Gleaned from Animal Chemoprevention Studies
Molecular Activities of Gingerols Gleaned from Cell Culture Models
Blood-Borne Cells
Digestive Cells
Breast Cells
Skin Cells
Other Cell Types
2.5.2. Cancer Chemopreventive Activities of Shogaols
Pharmacokinetics and Metabolism of Shogaols
Animal Initiation/Promotion Chemoprevention Studies
In Vitro Studies
Blood-Borne Cells
Digestive Cells
Breast Cells
Other Cell Types
2.6. Cellular and Molecular Activities of Paradols
2.7. Cellular and Molecular Activities of Zingerone
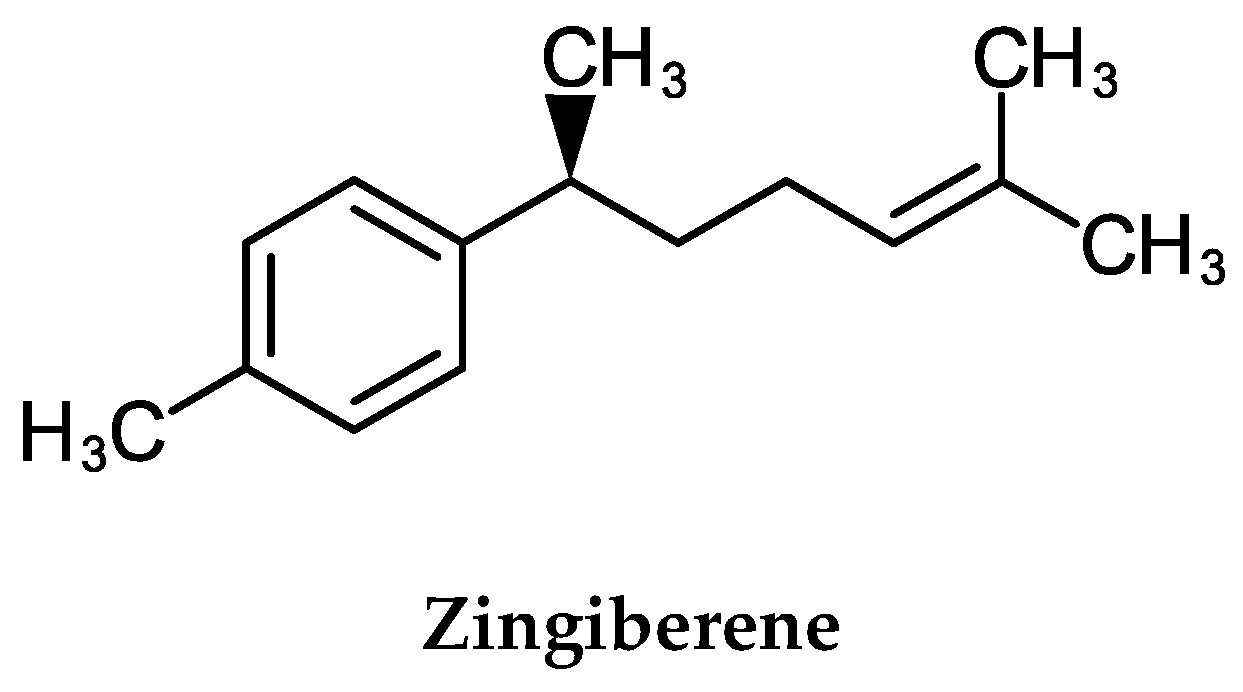
2.8. Cellular and Molecular Activities of α-Zingiberene
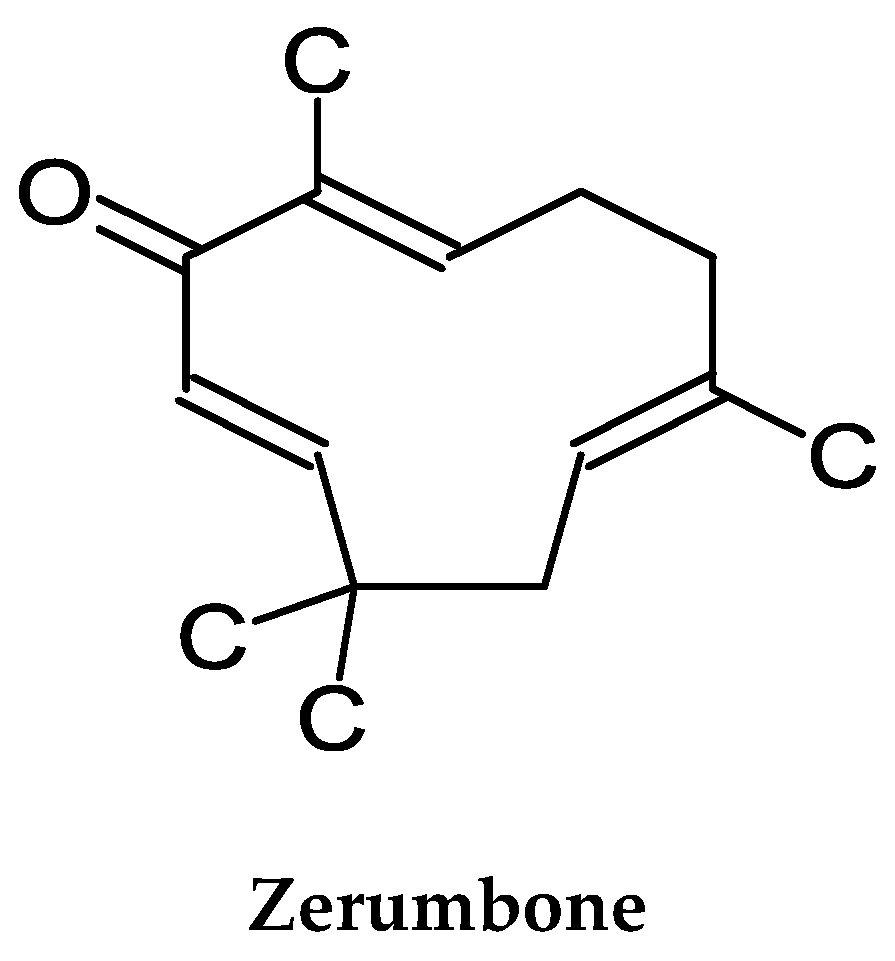
3. Bitter/Shampoo Ginger
3.1. Initiation/Promotion Studies
3.2. Blood-Borne Cells
3.3. Digestive Cells
3.4. Breast Cells
3.5. Skin Cells
3.6. Other Cells
4. Conclusions and Future Research
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAF | 2-Acetylaminofluorene |
| ACF | Aberrant crypt foci |
| αFP | α-Fetal Protein |
| AKR1B10 | Aldo-keto reductase family 1 member B10 |
| AKT | Protein kinase B |
| ALT | Alanine transaminase |
| AMPK | 5′ AMP-activated protein kinase |
| AOM | Azoxymethane |
| AP-1 | Activator protein 1 |
| ARE | AU-rich regulatory element |
| AST | Aspartate transaminase |
| BAK | Bcl-2 homologous antagonist/killer |
| BAX | Bcl-2-associated X protein |
| B[a]P | Benzo(a)pyrene |
| Bcl-2 | Bcl-2 (B-cell lymphoma 2) |
| Bcl-XL | B-cell lymphoma-extra large |
| Bid | BH3 interacting-domain death agonist |
| bFGF | Basic fibroblast growth factor |
| CD44 | CD44 antigen |
| cdc2 | Cyclin-dependent kinase 1 |
| cdc20 | Cell-division cycle protein 20 |
| cdc25A | Cell division cycle 25 homolog A |
| cdc25B | Cell division cycle 25 homolog B |
| cdc25C | Cell division cycle 25 homolog C |
| cdk1 | Cyclin-dependent kinase 1 |
| cdk4 | Cyclin-dependent kinase 4 |
| CEA | Carcinoembryonic antigen |
| c-FILP | CASP8 and FADD-like apoptosis regulator |
| c-myc | Member of a family of regulator genes |
| COX-2 | Prostaglandin-endoperoxide synthase 2 |
| c-Src | Proto-oncogene tyrosine-protein kinase Src |
| CXCL12 | Stromal cell-derived factor 1 |
| CXCR4 | C-X-C chemokine receptor type 4 |
| CypA | Cytochrome P450 A |
| DEN | Diethylnitrosamine |
| DMBA | 7,12-Dimethylbenz(a)anthracene |
| DMH | Dimethylhydrazine |
| DR | Death Receptor |
| E-CAD | Cadherin-1 |
| EGF | Epidermal growth factor |
| EGFR | Epidermal growth factor receptor |
| elF2α | Eukaryotic translation initiation factor 2A |
| ERK | Extracellular signal-egulated kinases |
| ESC | Esophageal Squamous Carcinoma |
| ETV-1 | ETS translocation variant 1 |
| FAK | Focal adhesion kinase, |
| FAS | Fas signaling cascade |
| FASL | FAS ligand |
| FGFR-1 | Fibroblast growth factor receptor 1 |
| FKHR | Forkhead box protein O1 |
| FTL | Ferritin light chain |
| GADD45α | Growth arrest and DNA-damage-inducible protein GADD45 alpha |
| GADD153 | Growth arrest and DNA-damage-inducible protein GADD153 |
| γ-GCLC | Glutamate-ysteine ligase catalytic subunit |
| GCS | Glutamate Cysteine Ligase |
| GGTLA4 | Gamma-glutamyltransferase 5 |
| Gli-1 | Zinc finger protein GLI1 |
| GPx1 | Glutathione peroxidase 1 |
| GRP-78 | Heat shock 70 kDa protein 5 |
| GSH | Glutathione |
| GSK-3β | Glycogen synthase kinase 3 beta |
| GSSG | Glutathione disulfide |
| GST | Glutathione S-transferase |
| GSTP1 | Glutathione S-transferase P |
| HDAC | Histone deacetylase |
| HER2 | Receptor tyrosine-protein kinase erbB-2 |
| HMOX1/HO-1 | heme oxygenase (decycling) 1 |
| HPV | Human papillomavirus |
| hTERT | Human telomerase reverse transcriptase |
| HUVEC | Human umbilical vein endothelial cells |
| IkB | Inhibitor of κB |
| IkB2 | Inhibitor of κB2 |
| IKK | IκB kinase |
| IKKB | Inhibitor of nuclear factor kappa-B kinase subunit beta |
| IL-1α | Interleukin 1 alpha |
| IL-1β | Interleukin 1 beta |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| INF-γ | Interferon gamma |
| iNOS | Inducible nitric oxide synthase |
| JAK | Janus kinase |
| JNK | c-Jun N-terminal kinase |
| Keap | Kelch-like ECH-associated protein 1 |
| KI 67 | Antigen KI-67 |
| LDH | Lactate dehydrogenase |
| LIMK1/2 | LIM domain kinase 1/2 |
| LPS | Lipopolysaccharide |
| Mad2 | Mitotic arrest deficient 2 |
| MAPK | Mitogen-activated protein kinase |
| Mcl-1 | Induced myeloid leukemia cell differentiation protein |
| MIB-1 | E3 ubiquitin-protein ligase |
| miR-27b | MiR-27b microRNA precursor |
| MMP-2 | Matrix metalloproteinase-2 |
| MMP-3 | Matrix metalloproteinase-3 |
| MMP-9 | Matrix metalloproteinase-9 |
| MnSOD | Superoxide dismutase 2, mitochondrial |
| MPM-2 | Cyclin dependent kinase 2 associated protein 2 pseudogene 2 |
| mTOR | Mechanistic target of rapamycin |
| MyD88 | Myeloid differentiation primary response 88 |
| NFκB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Notch2 | Neurogenic locus notch homolog protein 2 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| NSCLC | Non-small cell lung cancer |
| ODC | Ornithine decarboxylase |
| OSC | Oral squamous carcinoma |
| p21 | Cyclin-dependent kinase inhibitor 1 |
| p27 | Cyclin-dependent kinase inhibitor 1B |
| p53 | Tumor suppressor p53 |
| PAI-1 | Plasminogen activator inhibitor-1 |
| PARP | Poly ADP ribose polymerase |
| PCNA | Proliferating cell nuclear antigen |
| PGE2 | Prostaglandin E2 |
| PI3K | Phosphatidylinositol 3-kinase |
| PKC-α | Protein kinase C alpha |
| p38 MAPK | P38 mitogen-activated protein kinase |
| RCC | Renal cell carcinoma |
| RhoA | Ras homolog gene family, member A |
| RKIP | Raf kinase inhibitor protein |
| ROCK | Rho-associated protein kinase |
| ROS | Reactive oxygen species |
| SAA1 | Serum amyloid A1 |
| SHP-1 | Src homology region 2 domain-containing phosphatase-1 |
| Snail | Zinc finger protein SNAI1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TIMP-1 | TIMP metallopeptidase inhibitor 1 |
| TLR2 | Toll-like receptor 2 |
| TLR4 | Toll-like receptor 4 |
| TNF-α | TNF, tumor necrosis factor alpha |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TPA | 12-O-Tetradecanoylphorbol-13-acetate |
| uPA | Urokinase-type plasminogen activator |
| VEGF | Vascular endothelial growth factor |
| VEGFR-2 | Vascular endothelial growth factor receptor 2 |
| XIAP | X-linked inhibitor of apoptosis protein |
| ZO-1 | Zonula occludens-1 |
| 14-3-3 | 14-3-3 proteins |
References
- Truong, T.H.; Carroll, K.S. Redox Regulation of Protein Kinases. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 332–356. [Google Scholar] [CrossRef] [PubMed]
- Reczek, C.R.; Chandel, N.S. ROS-dependent Signal Transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Long. 2016, 18. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Z.; Min, W. Mitochondria, Oxidative Stress and Innate Immunity. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, Inflammation, and Cancer: How are they Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Kuo, C.T.; Huang, Y.W.; Stoner, G.D.; Lechner, J.F. Gene-Diet Interactions on Colorectal Cancer Risk. Curr. Nutr. Rep. 2012, 1, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Meitzler, J.; Antony, S.; Wu, Y.; Juhasz, A.; Liu, H.; Jiang, G.; Lu, J.; Roy, K.; Doroshow, J.H. NADPH Oxidases: A Perspective on Reactive Oxygen Species Production in Tumor Biology. Antioxidants Redox. Signal. 2014, 20, 2873–2889. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front. Immunol. 2018, 13. [Google Scholar] [CrossRef]
- Hofman, P.M. Pathobiology of the Neutrophil-intestinal Epithelial Cell Interaction: Role in Carcinogenesis. World J. Gastroenterol. 2010, 16, 5790–5800. [Google Scholar] [CrossRef]
- Hoffman, A.; Spetner, L.M.; Burke, M. Ramifications of a Redox Switch within a Normal Cell: Its Absence in a Cancer Cell. Free Radic. Biol. Med. 2008, 45, 265–268. [Google Scholar] [CrossRef]
- Wallace, D.R. Natural Products as a Source of Anti-Cancer Lead Compounds: Ginger and Breast Cancer. J. Pharm. Clin. Res. 2016, 1, 555564. [Google Scholar] [CrossRef]
- Mascolo, N.; Jain, R.; Jain, S.C.; Capasso, F. Ethnopharmacologic investigation of ginger (Zingiber officinale). J. Ethnopharmacol. 1989, 27, 129–140. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Kim, M.H.; Hong, J.; Kim, S.H.; Yang, W.M. Dried Ginger (Zingiber officinalis) Inhibits Inflammation in a Lipopolysaccharide-Induced Mouse Model. Evid. Based Complement. Alternat Med. 2013, 2013, 914563. [Google Scholar] [CrossRef] [PubMed]
- Manju, V.; Nalini, N. Chemopreventive Efficacy of Ginger, a Naturally Occurring Anticarcinogen during the Initiation, Post-initiation Stages of 1,2 Dimethylhydrazine-induced Colon Cancer. Clin. Chim. Acta. 2005, 358, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Karna, P.; Chagani, S.; Gundala, S.R.; Rida, P.C.; Asif, G.; Sharma, V.; Gupta, M.V.; Aneja, R. Benefits of Whole Ginger Extract in Prostate Cancer. Br. J. Nutr. 2012, 107, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Yoshimi, N.; Wang, A.; Morishita, Y.; Tanaka, T.; Sugie, S.; Kawai, K.; Yamahara, J.; Mori, H. Modifying Effects of Fungal and Herb Metabolites on Azoxymethane-induced Intestinal Carcinogenesis in Rats. Jpn. J. Cancer Res. 1992, 83, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Tuntiwechapikul, W.; Taka, T.; Songsomboon, C.; Kaewtunjai, N.; Imsumran, A.; Makonkawkeyoon, L.; Pompimon, W.; Lee, T.R. Ginger Extract Inhibits Human Telomerase Reverse Transcriptase and c-Myc Expression in A549 Lung Cancer Cells. J. Med. Food. 2010, 13, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Elkady, A.I.; Abuzinadah, O.A.; Baeshen, N.A.; Rahmy, T.R. Differential Control of Growth, Apoptotic Activity, and Gene Expression in Human Breast Cancer Cells by Extracts Derived from Medicinal Herbs Zingiber officinale. J. Biomed. Biotechnol. 2012, 2012, 614356. [Google Scholar] [CrossRef]
- Rhode, J.; Fogoros, S.; Zick, S.; Wahl, H.; Griffith, K.A.; Huang, J.; Liu, J.R. Ginger Inhibits Cell Growth and Modulates Angiogenic Factors in Ovarian Cancer Cells. BMC Complement. Altern Med. 2007, 7, 44. [Google Scholar] [CrossRef]
- Pashaei-Asl, R.; Pashaei-Asl, F.; Mostafa Gharabaghi, P.; Khodadadi, K.; Ebrahimi, M.; Ebrahimie, E.; Pashaiasl, M. The Inhibitory Effect of Ginger Extract on Ovarian Cancer Cell Line; Application of Systems Biology. Adv. Pharm. Bull. 2017, 7, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whelan, R.J.; Pattnaik, B.R.; Ludwig, K.; Subudhi, E.; Rowland, H.; Claussen, N.; Zucker, N.; Uppal, S.; Kushner, D.M.; et al. Terpenoids from Zingiber officinale (Ginger) Induce Apoptosis in Endometrial Cancer Cells Through the Activation of p53. PLoS ONE 2012, 7, e53178. [Google Scholar] [CrossRef] [PubMed]
- Vipin, A.V.; Raksha Rao, K.; Nawneet, K.K.; Anu Appaiah, K.A.; Vankateswaran, G. Protective Effects of Phenolics Rich Extract of Ginger Against Aflatoxin B1-induced Oxidative Stress and Hepatotoxicity. Biomed. Pharmacother. 2017, 91, 415–424. [Google Scholar]
- Akimoto, M.; Iizuka, M.; Kanematsu, R.; Yoshida, M.; Takenaga, K. Anticancer Effect of Ginger Extract against Pancreatic Cancer Cells Mainly through Reactive Oxygen Species-Mediated Autotic Cell Death. PLoS ONE 2015, 10, e0126605. [Google Scholar] [CrossRef] [PubMed]
- Mahady, G.B.; Pendland, S.L.; Yun, G.S.; Lu, Z.Z.; Stoia, A. Ginger (Zingiber officinale Roscoe) and the Gingerols Inhibit the Growth of Cag A+ Strains of Helicobacter pylori. Anticancer Res. 2003, 23, 3699–3702. [Google Scholar] [PubMed]
- Gaus, K.; Huang, Y.; Israel, D.A.; Pendland, S.L.; Adeniyi, B.A.; Mahady, G.B. Standardized Ginger (Zingiber officinale) Extract Reduces Bacterial Load and Suppresses Acute and Chronic Inflammation in Mongolian Gerbils Infected with CagA+ Helicobacter pylori. Pharm Biol. 2009, 47, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Zick, S.M.; Turgeon, D.K.; Vareed, S.K.; Ruffin, M.T.; Litzinger, A.J.; Wright, B.D.; Alrawi, S.; Normolle, D.P.; Djuric, Z.; Brenner, D.E. Phase II Study of the Effects of Ginger Root Extract on Eicosanoids in Colon Mucosa in People at Normal Risk for Colorectal Cancer. Cancer Prev. Res. 2011, 4, 1929–1937. [Google Scholar] [CrossRef] [PubMed]
- Citronberg, J.; Bostick, R.; Ahearn, T.; Turgeon, D.K.; Ruffin, M.T.; Djuric, Z.; Sen, A.; Brenner, D.E.; Zick, S.M. Effects of Ginger Supplementation on Cell-cycle Biomarkers in the Normal-appearing Colonic Mucosa of Patients at Increased Risk for Colorectal Cancer: Results from a Pilot, Randomized, and Controlled Trial. Cancer Prev. Res. 2013, 6, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.P.; Deol, P.K.; Kondepudi, K.K.; Bishnoi, M. Anticancer Potential of Ginger: Mechanistic and Pharmaceutical Aspects. Curr. Pharm. Des. 2016, 22, 4160–4172. [Google Scholar] [CrossRef] [PubMed]
- Almatroudi, A.; Alsahli, M.A.; Alrumaihi, F.; Allemailem, K.S.; Rahmani, A.H. Ginger: A Novel Strategy to Battle Cancer through Modulating Cell Signalling Pathways: A Review. Curr Pharm Biotechnol. 2019, 20, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Wang, N.S.; Mi, S.Q. Plasma Pharmacokinetics and Tissue Distribution of [6]-Gingerol in Rats. Biopharm. Drug Dispos. 2008, 29, 529–537. [Google Scholar] [CrossRef]
- Li, L.L.; Cui, Y.; Guo, X.H.; Ma, K.; Tian, P.; Feng, J.; Wang, J.M. Pharmacokinetics and Tissue Distribution of Gingerols and Shogaols from Ginger (Zingiber officinale Rosc.) in Rats by UPLC-Q-Exactive-HRMS. Molecules 2019, 24, 512. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Ohsawa, K. Metabolism of [6]-Gingerol in Rats. Life Sci. 2002, 70, 2165–2175. [Google Scholar] [CrossRef]
- Zick, S.M.; Djuric, Z.; Ruffin, M.T.; Litzinger, A.J.; Normolle, D.P.; Alrawi, S.; Feng, M.R.; Brenner, D.E. Pharmacokinetics of 6-Gingerol, 8-Gingerol, 10-Gingerol, and 6-Shogaol and Conjugate Metabolites in Healthy Human Subjects. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zick, S.; Li, X.; Zou, P.; Wright, B.; Sun, D. Examination of the Pharmacokinetics of Active Ingredients of Ginger in Humans. AAPS J. 2011, 13, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Mukkavilli, R.; Yang, C.; Singh Tanwar, R.; Ghareeb, A.; Luthra, L.; Aneja, R. Absorption, Metabolic Stability, and Pharmacokinetics of Ginger Phytochemicals. Molecules 2017, 22, 553. [Google Scholar] [CrossRef] [PubMed]
- Mukkavilli, R.; Yang, C.; Tanwar, R.S.; Saxena, R.; Gundala, S.; Zhang, Y.; Ghareeb, A.; Floyd, S.D.; Vangala, S.; Kuo, W.W.; et al. Pharmacokinetic-Pharmacodynamic Correlations in the Development of Ginger Extract as an Anticancer Agent. Sci. Rep. 2018, 8, 3056. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Agarwal, R.; Mukhtar, H. Inhibition of Tumor Promotion in SENCAR Mouse Skin by Ethanol Extract of Zingiber officinale Rhizome. Cancer Res. 1996, 56, 1023–1030. [Google Scholar]
- Park, K.K.; Chun, K.S.; Lee, J.M.; Lee, S.S.; Surh, Y.J. Inhibitory Effects of [6]-Gingerol, a Major Pungent Principle of Ginger, on Phorbol Ester-induced Inflammation, Epidermal Ornithine Decarboxylase Activity and Skin Tumor Promotion in ICR Mice. Cancer Lett. 1998, 129, 139–144. [Google Scholar] [CrossRef]
- Surh, Y.J.; Park, K.K.; Chun, K.S.; Lee, L.J.; Lee, E.; Lee, S.S. Anti-tumor-promoting Activities of Selected Pungent Phenolic Substances Present in Ginger. J. Environ. Pathol. Toxicol. Oncol. 1999, 18, 131–139. [Google Scholar]
- Nigam, N.; George, J.; Srivastava, S.; Roy, P.; Bhui, K.; Singh, M.; Shukla, Y. Induction of Apoptosis by [6]-Gingerol Associated with the Modulation of p53 and Involvement of Mitochondrial Signaling Pathway in B[a]P-induced Mouse Skin Tumorigenesis. Cancer Chemother. Pharmacol. 2010, 65, 687–696. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, Y.; Na, K.M.; Surh, Y.J.; Kim, T.Y. [6]-Gingerol Prevents UVB-induced ROS Production and COX-2 Expression in vitro and in vivo. Free Radic. Res. 2007, 41, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Shukla, Y.; Prasad, S.; Tripathi, C.; Singh, M.; George, J.; Kalra, N. In vitro and in vivo Modulation of Testosterone Mediated Alterations in Apoptosis Related Proteins by [6]-Gingerol. Mol. Nutr. Food Res. 2007, 51, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Ippoushi, K.; Azuma, K.; Ito, H.; Horie, H.; Higashio, H. [6]-Gingerol Inhibits Nitric Oxide Synthesis in Activated J774.1 Mouse Macrophages and Prevents Peroxynitrite-induced Oxidation and Nitration Reactions. Life Sci. 2003, 73, 3427–3437. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.Y.; Lee, K.C.; Chen, S.Y.; Chang, H.H. 6-Gingerol Inhibits ROS and iNOS Through the Suppression of PKC-alpha and NF-kappaB Pathways in Lipopolysaccharide-stimulated Mouse Macrophages. Biochem. Biophys. Res. Commun. 2009, 382, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, N.; Gara, R.K.; Trivedi, R.; Singh, A.; Dixit, P.; Maurya, R.; Duggal, S.; Bhatt, M.L.; Singh, S.; Mishra, D.P. (6)-Gingerol Induced Myeloid Leukemia Cell Death is Initiated by Reactive Oxygen Species and Activation of miR-27b Expression. Free Radic. Biol. Med. 2014, 68, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Park, S.H.; Lee, M.; Kim, H.J.; Ryu, S.Y.; Kim, N.D.; Hwang, B.Y.; Hong, J.T.; Han, S.B.; Kim, Y. 1-Dehydro-[10]-gingerdione from Ginger Inhibits IKKβ Activity for NF-κB Activation and Suppresses NF-κB-regulated Expression of Inflammatory Genes. Br. J. Pharmacol. 2012, 167, 128–140. [Google Scholar] [CrossRef]
- Park, S.H.; Kyeong, M.S.; Hwang, Y.; Ryu, S.Y.; Han, S.B.; Kim, Y. Inhibition of LPS Binding to MD-2 Co-receptor for Suppressing TLR4-mediated Expression of Inflammatory Cytokine by 1-Dehydro-10-gingerdione from Dietary Ginger. Biochem. Biophys. Res. Commun. 2012, 419, 735–740. [Google Scholar] [CrossRef]
- Brown, A.C.; Shah, C.; Liu, J.; Pham, J.T.; Zhang, J.G.; Jadus, M.R. Ginger’s (Zingiber officinale Roscoe) Inhibition of Rat Colonic Adenocarcinoma Cells Proliferation and Angiogenesis in vitro. Phytother. Res. 2009, 23, 640–645. [Google Scholar] [CrossRef]
- Lee, S.H.; Cekanova, M.; Baek, S.J. Multiple Mechanisms Are Involved in 6-Gingerol-induced Cell Growth Arrest and Apoptosis in Human Colorectal Cancer Cells. Mol. Carcinog. 2008, 47, 197–208. [Google Scholar] [CrossRef]
- Lin, C.B.; Lin, C.C.; Tsay, G.J. 6-Gingerol Inhibits Growth of Colon Cancer Cell LoVo via Induction of G2/M Arrest. Evid Based Complement Alternat Med. 2012, 2012, 326096. [Google Scholar] [CrossRef]
- Radhakrishnan, E.K.; Bava, S.V.; Narayanan, S.S.; Nath, L.R.; Thulasidasan, A.K.; Soniya, E.V.; Anto, R.J. [6]-Gingerol Induces Caspase-dependent Apoptosis and Prevents PMA-induced Proliferation in Colon Cancer Cells by Inhibiting MAPK/AP-1 Signaling. PLoS ONE. 2014, 9. [Google Scholar] [CrossRef]
- Jeong, C.H.; Bode, A.M.; Pugliese, A.; Cho, Y.Y.; Kim, H.G.; Shim, J.H.; Jeon, Y.J.; Li, H.; Jiang, H.; Dong, Z. [6]-Gingerol Suppresses Colon Cancer Growth by Targeting Leukotriene A4 Hydrolase. Cancer Res. 2009, 69, 5584–5591. [Google Scholar] [CrossRef]
- El-Naggar, M.H.; Mira, A.; Abdel Bar, F.M.; Shimizu, K.; Amer, M.M.; Badria, F.A. Synthesis, Docking, Cytotoxicity, and LTA4H Inhibitory Activity of New Gingerol Derivatives as Potential Colorectal Cancer. Therapy. Bioorg. Med. Chem. 2017, 25, 1277–1285. [Google Scholar] [CrossRef]
- Mansingh, D.P.; Sunanda, O.J.; Sali, V.K.; Vasanthi, H.R. [6]-Gingerol-induced Cell Cycle Arrest, Reactive Oxygen Species Generation, and Disruption of Mitochondrial Membrane Potential Are Associated with Apoptosis in Human Gastric Cancer (AGS) Cells. J. Biochem. Mol. Toxicol. 2018, 32. [Google Scholar] [CrossRef]
- Chen, C.Y.; Li, Y.W.; Kuo, S.Y. Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells. Molecules 2009, 14. [Google Scholar] [CrossRef]
- Kim, S.O.; Kim, M.R. [6]-Gingerol Prevents Disassembly of Cell Junctions and Activities of MMPs in Invasive Human Pancreas Cancer Cells through ERK/NF- κ B/Snail Signal Transduction Pathway. Evid. Based Complement. Alternat. Med. 2013, 2013, 761852. [Google Scholar] [CrossRef]
- Yagihashi, S.; Miura, Y.; Yagasaki, K. Inhibitory Effect of Gingerol on the Proliferation and Invasion of Hepatoma Cells in Culture. Cytotechnology 2008, 57, 129–136. [Google Scholar] [CrossRef]
- Li, X.H.; McGrath, K.C.; Tran, V.H.; Li, Y.M.; Duke, C.C.; Roufogalis, B.D.; Heather, A.K. Attenuation of Proinflammatory Responses by S-[6]-Gingerol via Inhibition of ROS/NF-Kappa B/COX2 Activation in HuH7 Cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 146142. [Google Scholar] [CrossRef]
- Lee, H.S.; Seo, E.Y.; Kang, N.E.; Kim, W.K. [6]-Gingerol Inhibits Metastasis of MDA-MB-231 Human Breast Cancer Cells. J. Nutr. Biochem. 2008, 19, 313–319. [Google Scholar] [CrossRef]
- Bernard, M.M.; McConnery, J.R.; Hoskin, D.W. [10]-Gingerol, a Major Phenolic Constituent of Ginger Root, Induces Cell Cycle Arrest and Apoptosis in Triple-negative Breast Cancer Cells. Exp. Mol. Pathol. 2017, 102, 370–376. [Google Scholar] [CrossRef]
- Fuzer, A.M.; Lee, S.Y.; Mott, J.D.; Cominetti, M.R. [10]-Gingerol Reverts Malignant Phenotype of Breast Cancer Cells in 3D Culture. J. Cell Biochem. 2017, 118, 2693–2699. [Google Scholar] [CrossRef] [PubMed]
- Fuzer, A.M.; Martin, A.C.B.M.; Becceneri, A.B.; da Silva, J.A.; Vieira, P.C.; Cominetti, M.R. [10]-Gingerol Affects Multiple Metastatic Processes and Induces Apoptosis in MDA-MB-231 Breast Tumor Cells. Anticancer Agents Med. Chem. 2019, 19, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.C.B.M.; Fuzer, A.M.; Becceneri, A.B.; da Silva, J.A.; Tomasin, R.; Denoyer, D.; Kim, S.H.; McIntyre, K.A.; Pearson, H.B.; Yeo, B.; et al. [10]-Gingerol Induces Apoptosis and Inhibits Metastatic Dissemination of Triple Negative Breast Cancer in vivo. Oncotarget 2017, 8, 72260–72271. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.H.; Hong, S.S.; Cho, Y.R.; Seo, D.W. 10-Gingerol Inhibits Proliferation and Invasion of MDA-MB-231 Breast Cancer Cells through Suppression of Akt and p38MAPK Activity. Oncol. Rep. 2016, 35, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.O.; Chun, K.S.; Kundu, J.K.; Surh, Y.J. Inhibitory Effects of [6]-Gingerol on PMA-induced COX-2 Expression and Activation of NF-kappaB and p38 MAPK in Mouse Skin. Biofactors 2004, 21, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.O.; Kundu, J.K.; Shin, Y.K.; Park, J.H.; Cho, M.H.; Kim, T.Y.; Surh, Y.J. [6]-Gingerol Inhibits COX-2 Expression by Blocking the Activation of p38 MAP Kinase and NF-kappaB in Phorbol Ester-Stimulated Mouse Skin. Oncogene 2005, 24, 2558–2567. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Chiu, S.H.; Chang, T.M. Inhibitory Effect of [6]-Gingerol on Melanogenesis in B16F10 Melanoma Cells and a Possible Mechanism of Action. Biosci. Biotechnol. Biochem. 2011, 75, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.C.; Min, J.K.; Kim, T.Y.; Lee, S.J.; Yang, H.O.; Han, S.; Kim, Y.M.; Kwon, Y.G. [6]-Gingerol, a Pungent Ingredient of Ginger, Inhibits Angiogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2005, 335, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Thakur, K.; Hu, F.; Zhang, J.G.; Wei, Z.J. 10-Gingerol, a Phytochemical Derivative from “Tongling White Ginger”, Inhibits Cervical Cancer: Insights into the Molecular Mechanism and Inhibitory Targets. J. Agric. Food Chem. 2017, 65, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.; Bishayee, K.; Ghosh, S.; Biswas, R.; Mandal, S.K.; Khuda-Bukhsh, A.R. [6]-Gingerol Induces Caspase 3 Dependent Apoptosis and Autophagy in Cancer Cells: Drug-DNA Interaction and Expression of Certain Signal Genes in HeLa Cells. Eur. J. Pharmacol. 2012, 694, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Haris, P.; Mary, V.; Sudarsanakumar, C. Probing the Interaction of the Phytochemical 6-Gingerol from the Spice Ginger with DNA. Int. J. Biol. Macromol. 2018, 113, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, N.; Duggal, S.; Singh, S.K.; Porwal, K.; Srivastava, V.K.; Maurya, R.; Bhatt, M.L.; Mishra, D.P. Proteasome Inhibition Mediates p53 Reactivation and Anti-cancer Activity of 6-Gingerol in Cervical Cancer Cells. Oncotarget. 2015, 6, 43310–43325. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, D.W.; Jung, C.H.; Lee, Y.J.; Park, D. Gingerol Sensitizes TRAIL-Induced Apoptotic Cell Death of Glioblastoma Cells. Toxicol. Appl. Pharmacol. 2014, 279, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Asami, A.; Shimada, T.; Mizuhara, Y.; Asano, T.; Takeda, S.; Aburada, T.; Miyamoto, K.; Aburada, M. Pharmacokinetics of [6]-Shogaol, a Pungent Ingredient of Zingiber officinale Roscoe (Part I). J. Nat. Med. 2010, 64, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lv, L.; Soroka, D.; Warin, R.F.; Parks, T.A.; Hu, Y.; Zhu, Y.; Chen, X.; Sang, S. Metabolism of [6]-Shogaol in Mice and in Cancer Cells. Drug Metab. Dispos. 2012, 40, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Chen, H.; Soroka, D.N.; Warin, R.F.; Sang, S. Cysteine-conjugated Metabolites of Ginger Components, Shogaols, Induce Apoptosis Through Oxidative Stress-mediated p53 Pathway in Human Colon Cancer Cells. J. Agric. Food Chem. 2014, 62, 4632–4642. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Warin, R.F.; Soroka, D.N.; Chen, H.; Sang, S. Metabolites of Ginger Component [6]-Shogaol Remain Bioactive in Cancer Cells and have Low Toxicity in Normal Cells: Chemical Synthesis and Biological Evaluation. PLoS ONE. 2013, 8, e54677. [Google Scholar] [CrossRef]
- Chen, H.; Soroka, D.N.; Zhu, Y.; Hu, Y.; Chen, X.; Sang, S. Cysteine-conjugated Metabolite of Ginger Component [6]-Shogaol Serves as a Carrier of [6]-Shogaol in Cancer Cells and in Mice. Chem. Res. Toxicol. 2013, 26, 976–985. [Google Scholar] [CrossRef]
- Warin, R.F.; Chen, H.; Soroka, D.N.; Zhu, Y.; Sang, S. Induction of Lung Cancer Cell Apoptosis Through a p53 Pathway by [6]-Shogaol and its Cysteine-conjugated Metabolite M2. J. Agric Food Chem. 2014, 62, 1352–1362. [Google Scholar] [CrossRef]
- Chen, H.; Fu, J.; Chen, H.; Hu, Y.; Soroka, D.N.; Prigge, J.R.; Schmidt, E.E.; Yan, F.; Major, M.B.; Chen, X.; et al. Ginger Compound [6]-Shogaol and its Cysteine-conjugated Metabolite (M2) Activate Nrf2 in Colon Epithelial Cells in vitro and in vivo. Chem. Res. Toxicol. 2014, 27, 1575–1585. [Google Scholar] [CrossRef]
- Wu, H.; Hsieh, M.C.; Lo, C.Y.; Liu, C.B.; Sang, S.; Ho, C.T.; Pan, M.H. 6-Shogaol is More Effective than 6-Gingerol and Curcumin in Inhibiting 12-O-Tetradecanoylphorbol 13-acetate-induced Tumor Promotion in Mice. Mol. Nutr. Food Res. 2010, 54, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Hsieh, M.C.; Hsu, P.C.; Ho, S.Y.; Lai, C.S.; Wu, H.; Sang, S.; Ho, C.T. 6-Shogaol Suppressed Lipopolysaccharide-induced Up-expression of iNOS and COX-2 in Murine Macrophages. Mol. Nutr. Food Res. 2008, 52, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Shieh, P.C.; Chen, Y.O.; Kuo, D.H.; Chen, F.A.; Tsai, M.L.; Chang, I.S.; Wu, H.; Sang, S.; Ho, C.T.; Pan, M.H. Induction of Apoptosis by [8]-Shogaol via Reactive Oxygen Species Generation, Glutathione Depletion, and Caspase Activation in Human Leukemia Cells. J. Agric. Food Chem. 2010, 58, 3847–3854. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.I.; Lee, J.K.; Youn, H.S. Inhibition of Homodimerization of Toll-like Receptor 4 by 6-Shogaol. Mol. Cells 2009, 27, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Peng, Y.B.; Zhou, P.; Qi, L.W.; Zhang, M.; Gao, N.; Liu, E.H.; Li, P. 6-Shogaol Induces Apoptosis in Human Leukemia Cells through a Process Involving Caspase-mediated Cleavage of eIF2α. Mol. Cancer 2013, 12, 135. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.W.; Zhang, Z.; Zhang, C.F.; Anderson, S.; Liu, Q.; Yuan, C.S.; Wang, C.Z. Anti-Colon Cancer Effects of 6-Shogaol Through G2/M Cell Cycle Arrest by p53/p21-cdc2/cdc25A Crosstalk. Am. J. Chin. Med. 2015, 43, 743–756. [Google Scholar] [CrossRef]
- Pan, M.H.; Hsieh, M.C.; Kuo, J.M.; Lai, C.S.; Wu, H.; Sang, S.; Ho, C.T. 6-Shogaol Induces Apoptosis in Human Colorectal Carcinoma Cells via ROS Production, Caspase Activation, and GADD 153 Expression. Mol. Nutr. Food Res. 2008, 52, 527–537. [Google Scholar] [CrossRef]
- Sang, S.; Hong, J.; Wu, H.; Liu, J.; Yang, C.S.; Pan, M.H.; Badmaev, V.; Ho, C.T. Increased Growth Inhibitory Effects on Human Cancer Cells and Anti-inflammatory Potency of Shogaols from Zingiber officinale Relative to Gingerols. J. Agric. Food Chem. 2009, 57, 10645–10650. [Google Scholar] [CrossRef]
- Gan, F.F.; Nagle, A.A.; Ang, X.; Ho, O.H.; Tan, S.H.; Yang, H.; Chui, W.K.; Chew, E.H. Shogaols at Proapoptotic Concentrations Induce G(2)/M Arrest and Aberrant Mitotic Cell Death Associated with Tubulin Aggregation. Apoptosis 2011, 16, 856–867. [Google Scholar] [CrossRef]
- Ishiguro, K.; Ando, T.; Watanabe, O.; Goto, H. Specific Reaction of α,β-unsaturated Carbonyl Compounds such as 6-Shogaol with Sulfhydryl Groups in Tubulin Leading to Microtubule Damage. FEBS Lett. 2008, 582, 3531–3536. [Google Scholar] [CrossRef]
- Chen, C.Y.; Liu, T.Z.; Liu, Y.W.; Tseng, W.C.; Liu, R.H.; Lu, F.J.; Lin, Y.S.; Kuo, S.H.; Chen, C.H. 6-Shogaol (Alkanone from Ginger) Induces Apoptotic Cell Death of Human Hepatoma p53 Mutant Mahlavu Subline via an Oxidative Stress-mediated Caspase-dependent Mechanism. J. Agric. Food Chem. 2007, 55, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhou, P.; Peng, Y.B.; Xu, X.; Ma, J.; Liu, Q.; Zhang, L.; Wen, X.D.; Qi, L.W.; Gao, N.; et al. 6-Shogaol Induces Apoptosis in Human Hepatocellular Carcinoma Cells and Exhibits Anti-tumor Activity in vivo Through Endoplasmic Reticulum Stress. PLoS ONE. 2012, 7. [Google Scholar] [CrossRef]
- Kim, J.K.; Jang, H.D. 6-Shogaol Attenuates H2O2-induced Oxidative Stress via Upregulation of Nrf2-mediated γ-Glutamylcysteine Synthetase and Heme Oxygenase Expression in HepG2 Cells. Food Sci. Biotechnol. 2016, 25, 319–327. [Google Scholar] [CrossRef]
- Nazim, U.M.; Park, S.Y. Attenuation of Autophagy Flux by 6-Shogaol Sensitizes Human Liver Cancer Cells to TRAIL-induced Apoptosis via p53 and ROS. Int. J. Mol. Med. 2019, 43, 701–708. [Google Scholar] [CrossRef]
- Weng, C.J.; Wu, C.F.; Huang, H.W.; Ho, C.T.; Yen, G.C. Anti-invasion Effects of 6-Shogaol and 6-Gingerol, Two Active Components in Ginger, on Human Hepatocarcinoma Cells. Mol. Nutr. Food Res. 2010, 54, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Chou, C.P.; Ho, C.T.; Yen, G.C. Molecular Mechanism Inhibiting Human Hepatocarcinoma Cell Invasion by 6-Shogaol and 6-Gingerol. Mol. Nutr. Food Res. 2012, 56, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Yang, H.; Tan, S.H.; Chui, W.K.; Chew, E.H. 6-Shogaol, an Active Constituent of Ginger, Inhibits Breast Cancer Cell Invasion by Reducing Matrix Metalloproteinase-9 Expression via Blockade of Nuclear Factor-κB Activation. Br. J. Pharmacol. 2010, 161, 1763–1777. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, C.; Bae, H.; Lee, J.H.; Baek, S.H.; Nam, D.; Chung, W.S.; Shim, B.S.; Lee, S.G.; Kim, S.H.; et al. 6-Shogaol Exerts Anti-proliferative and Pro-apoptotic Effects Through the Modulation of STAT3 and MAPKs Signaling Pathways. Mol. Carcinog. 2015, 54, 1132–1146. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Chen, C.Y.; Lin, I.P.; Tsai, E.M.; Kuo, P.L.; Hou, M.F. 4-Shogaol, an Active Constituent of Dietary Ginger, Inhibits Metastasis of MDA-MB-231 Human Breast Adenocarcinoma Cells by Decreasing the Repression of NF-κB/Snail on RKIP. J. Agric. Food Chem. 2012, 60, 852–861. [Google Scholar] [CrossRef]
- Hung, J.Y.; Hsu, Y.L.; Li, C.T.; Ko, Y.C.; Ni, W.C.; Huang, M.S.; Kuo, P.L. 6-Shogaol, an Active Constituent of Dietary Ginger, Induces Autophagy by Inhibiting the AKT/mTOR Pathway in Human Non-small Cell Lung Cancer A549 Cells. J. Agric. Food Chem. 2009, 57, 9809–9816. [Google Scholar] [CrossRef]
- Saha, A.; Blando, J.; Silver, E.; Beltran, L.; Sessler, J.; DiGiovanni, J. 6-Shogaol from Dried Ginger Inhibits Growth of Prostate Cancer Cells both in vitro and in vivo Through Inhibition of STAT3 and NF-κB Signaling. Cancer Prev. Res. 2014, 7, 627–638. [Google Scholar] [CrossRef]
- Liu, Q.; Peng, Y.B.; Qi, L.W.; Cheng, X.L.; Xu, X.J.; Liu, L.L.; Liu, E.H.; Li, P. The Cytotoxicity Mechanism of 6-Shogaol-Treated HeLa Human Cervical Cancer Cells Revealed by Label-Free Shotgun Proteomics and Bioinformatics Analysis. Evid. Based Complement. Alternat. Med. 2012. [Google Scholar] [CrossRef]
- Kim, M.O.; Lee, M.H.; Oi, N.; Kim, S.H.; Bae, K.B.; Huang, Z.; Kim, D.J.; Reddy, K.; Lee, S.Y.; Park, S.J.; et al. [6]-Shogaol Inhibits Growth and Induces Apoptosis of Non-small Cell Lung Cancer Cells by Directly Regulating Akt1/2. Carcinogenesis. 2014, 35, 683–691. [Google Scholar] [CrossRef]
- Lee, E.; Surh, Y.J. Induction of Apoptosis in HL-60 Cells by Pungent Vanilloids, [6]-Gingerol and [6]-Paradol. Cancer Lett. 1998, 134, 163–168. [Google Scholar] [CrossRef]
- Suresh, K.; Manoharan, S.; Vijayaanand, M.A.; Sugunadevi, G. Chemopreventive and Antioxidant Efficacy of (6)-Paradol in 7,12-Dimethylbenz(a)anthracene Induced Hamster Buccal Pouch Carcinogenesis. Pharmacol. Rep. 2010, 62, 1178–1185. [Google Scholar] [CrossRef]
- Mariadoss, A.V.; Kathiresan, S.; Muthusamy, R.; Kathiresan, S. Protective Effects of [6]-Paradol on Histological Lesions and Immunohistochemical Gene Expression in DMBA Induced Hamster Buccal Pouch Carcinogenesis. Asian. Pac. J. Cancer Prev. 2013, 14, 3123–3129. [Google Scholar] [CrossRef]
- Bode, A.M.; Ma, W.Y.; Surh, Y.J.; Dong, Z. Inhibition of Epidermal Growth Factor-induced Cell Transformation and Activator Protein 1 Activation by [6]-Gingerol. Cancer Res. 2001, 61, 850–853. [Google Scholar]
- Keum, Y.S.; Kim, J.; Lee, K.H.; Park, K.K.; Surh, Y.J.; Lee, J.M.; Lee, S.S.; Yoon, J.H.; Joo, S.Y.; Cha, I.H.; et al. Induction of Apoptosis and Caspase-3 Activation by Chemopreventive [6]-Paradol and Structurally Related Compounds in KB Cells. Cancer Lett. 2002, 177, 41–47. [Google Scholar] [CrossRef]
- Bae, W.Y.; Choi, J.S.; Kim, J.E.; Park, C.; Jeong, J.W. Zingerone Suppresses Angiogenesis via Inhibition of Matrix Metalloproteinases During Tumor Development. Oncotarget 2016, 7, 47232–47241. [Google Scholar] [CrossRef]
- Choi, J.S.; Ryu, J.; Bae, W.Y.; Park, A.; Nam, S.; Kim, J.E.; Jeong, J.W. Zingerone Suppresses Tumor Development through Decreasing Cyclin D1 Expression and Inducing Mitotic Arrest. Int. J. Mol. Sci. 2018, 19, 2832. [Google Scholar] [CrossRef]
- Ganaie, M.A.; Al Saeedan, A.; Madhkali, H.; Jan, B.L.; Khatlani, T.; Sheikh, I.A.; Rehman, M.U.; Wani, K. Chemopreventive Efficacy of Zingerone (4-[4-hydroxy-3-methylphenyl] butan-2-one) in Experimental Colon Carcinogenesis in Wistar Rats. Environ. Toxicol. 2019, 34, 610–625. [Google Scholar] [CrossRef]
- Marrelli, M.; Menichini, F.; Conforti, F. A Comparative Study of Zingiber officinale Roscoe Pulp and Peel: Phytochemical Composition and Evaluation of Antitumour Activity. Nat. Prod. Res. 2015, 29, 2045–2049. [Google Scholar] [CrossRef]
- Lee, Y. Cytotoxicity Evaluation of Essential Oil and its Component from Zingiber officinale Roscoe. Toxicol. Res. 2016, 32, 225–230. [Google Scholar] [CrossRef]
- Koga, A.Y.; Beltrame, F.L.; Pereira, A.V. Several Aspects of Zingiber zerumbet: A Review. Revista Brasileira de Farmacognosia. 2016, 26, 385–391. [Google Scholar] [CrossRef]
- Murakami, A.; Tanaka, T.; Lee, J.Y.; Surh, Y.J.; Kim, H.W.; Kawabata, K.; Nakamura, Y.; Jiwajinda, S.; Ohigashi, H. Zerumbone, a Sesquiterpene in Subtropical Ginger, Suppresses Skin Tumor Initiation and Promotion Stages in ICR Mice. Int. J. Cancer. 2004, 110, 481–490. [Google Scholar] [CrossRef]
- Tanaka, T.; Shimizu, M.; Kohno, H.; Yoshitani, S.; Tsukio, Y.; Murakami, A.; Safitri, R.; Takahashi, D.; Yamamoto, K.; Koshimizu, K.; et al. Chemoprevention of Azoxymethane-induced Rat Aberrant Crypt Foci by Dietary Zerumbone Isolated from Zingiber zerumbet. Biochem. Pharmacol. 2003, 66, 1253–1261. [Google Scholar] [CrossRef]
- Taha, M.M.; Abdul, A.B.; Abdullah, R.; Ibrahim, T.A.; Abdelwahab, S.I.; Mohan, S. Potential Chemoprevention of Diethylnitrosamine-initiated and 2-Acetylaminofluorene-promoted Hepatocarcinogenesis by Zerumbone from the Rhizomes of the Subtropical Ginger (Zingiber zerumbet). Chem. Biol. Interact. 2010, 186, 295–305. [Google Scholar] [CrossRef]
- Abdelwahab, S.I.; Abdul, A.B.; Devi, N.; Taha, M.M.; Al-zubairi, A.S.; Mohan, S.; Mariod, A.A. Regression of Cervical Intraepithelial Neoplasia by Zerumbone in Female Balb/c Mice Prenatally Exposed to Diethylstilboesterol: Involvement of Mitochondria-regulated Apoptosis. Exp. Toxicol. Pathol. 2010, 62, 461–469. [Google Scholar] [CrossRef]
- Kim, M.; Miyamoto, S.; Yasui, Y.; Oyama, T.; Murakami, A.; Tanaka, T. Zerumbone, a Tropical Ginger Sesquiterpene, Inhibits Colon and Lung Carcinogenesis in Mice. Int. J. Cancer. 2009, 124, 264–271. [Google Scholar] [CrossRef]
- Kim, M.J.; Yun, J.M. Molecular Mechanism of the Protective Effect of Zerumbone on Lipopolysaccharide-Induced Inflammation of THP-1 Cell-Derived Macrophages. J. Med. Food. 2019, 22, 62–73. [Google Scholar] [CrossRef]
- Haque, M.A.; Jantan, I.; Harikrishnan, H. Zerumbone Suppresses the Activation of Inflammatory Mediators in LPS-stimulated U937 Macrophages through MyD88-dependent NF-κB/MAPK/PI3K-Akt Signaling Pathways. Int. Immunopharmacol. 2018, 55, 312–322. [Google Scholar] [CrossRef]
- Murakami, A.; Takahashi, D.; Kinoshita, T.; Koshimizu, K.; Kim, H.W.; Yoshihiro, A.; Nakamura, Y.; Jiwajinda, S.; Terao, J.; Ohigashi, H. Zerumbone, a Southeast Asian Ginger Sesquiterpene, Markedly Suppresses Free Radical Generation, Proinflammatory Protein Production, and Cancer Cell Proliferation Accompanied by Apoptosis: The Alpha,Beta-unsaturated Carbonyl Group is a Prerequisite. Carcinogenesis 2002, 23, 795–802. [Google Scholar] [CrossRef]
- Rajan, I.; Jayasree, P.R.; Kumar, P.R. Zerumbone Induces Mitochondria-mediated Apoptosis via Increased Calcium, Generation of Reactive Oxygen Species and Upregulation of Soluble Histone H2AX in K562 Chronic Myelogenous Leukemia Cells. Tumour Biol. 2015, 36, 8479–8489. [Google Scholar] [CrossRef]
- Rahman, H.S.; Rasedee, A.; Chartrand, M.S.; Othman, H.H.; Yeap, S.K.; Namvar, F. Zerumbone Induces G2/M Cell Cycle Arrest and Apoptosis via Mitochondrial Pathway in Jurkat Cell Line. Nat. Prod. Commun. 2014, 9, 1237–1242. [Google Scholar] [CrossRef]
- Huang, G.C.; Chien, T.Y.; Chen, L.G.; Wang, C.C. Antitumor Effects of Zerumbone from Zingiber zerumbet in P-388D1 Cells in vitro and in vivo. Planta Med. 2005, 71, 219–224. [Google Scholar] [CrossRef]
- Xian, M.; Ito, K.; Nakazato, T.; Shimizu, T.; Chen, C.K.; Yamato, K.; Murakami, A.; Ohigashi, H.; Ikeda, Y.; Kizaki, M. Zerumbone, a Bioactive Sesquiterpene, Induces G2/M Cell Cycle Arrest and Apoptosis in leukemia Cells via a Fas- and Mitochondria-mediated Pathway. Cancer Sci. 2007, 98, 118–126. [Google Scholar] [CrossRef]
- Murakami, A.; Miyamoto, M.; Ohigashi, H. Zerumbone, an Anti-inflammatory Phytochemical, Induces Expression of Proinflammatory Cytokine Genes in Human Colon Adenocarcinoma Cell Lines. Biofactors 2004, 21, 95–101. [Google Scholar] [CrossRef]
- Yodkeeree, S.; Sung, B.; Limtrakul, P.; Aggarwal, B.B. Zerumbone Enhances TRAIL-induced Apoptosis through the Induction of Death Receptors in Human Colon Cancer Cells: Evidence for an Essential Role of Reactive Oxygen Species. Cancer Res. 2009, 69, 6581–6589. [Google Scholar] [CrossRef]
- Singh, S.P.; Nongalleima, K.; Singh, N.I.; Doley, P.; Singh, C.B.; Singh, T.R.; Sahoo, D. Zerumbone Reduces Proliferation of HCT116 Colon Cancer Cells by Inhibition of TNF-alpha. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Sidahmed, H.M.; Hashim, N.M.; Abdulla, M.A.; Ali, H.M.; Mohan, S.; Abdelwahab, S.I.; Taha, M.M.; Fai, L.M.; Vadivelu, J. Antisecretory, Gastroprotective, Antioxidant and anti-Helicobcter pylori Activity of Zerumbone from Zingiber zerumbet (L.) Smith. PLoS ONE. 2015, 10. [Google Scholar] [CrossRef]
- Tsuboi, K.; Matsuo, Y.; Shamoto, T.; Shibata, T.; Koide, S.; Morimoto, M.; Guha, S.; Sung, B.; Aggarwal, B.B.; Takahashi, H.; et al. Zerumbone Inhibits Tumor Angiogenesis via NF-κB in Gastric Cancer. Oncol. Rep. 2014, 31, 57–64. [Google Scholar] [CrossRef]
- Shamoto, T.; Matsuo, Y.; Shibata, T.; Tsuboi, K.; Nagasaki, T.; Takahashi, H.; Funahashi, H.; Okada, Y.; Takeyama, H. Zerumbone Inhibits Angiogenesis by Blocking NF-κB Activity in Pancreatic Cancer. Pancreas 2014, 43, 396–404. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Q.; Liu, Y.; Qiao, H.; Liu, Y. Zerumbone, a Southeast Asian Ginger Sesquiterpene, Induced Apoptosis of Pancreatic Carcinoma Cells through p53 Signaling Pathway. Evid. Based Complement. Alternat. Med. 2012. [Google Scholar] [CrossRef]
- Sakinah, S.A.; Handayani, S.T.; Hawariah, L.P. Zerumbone Induced Apoptosis in Liver Cancer Cells via Modulation of Bax/Bcl-2 Ratio. Cancer Cell Int. 2007, 7. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yoshida, C.; Murakami, A.; Ohigashi, H.; Osawa, T.; Uchida, K. Zerumbone, a Tropical Ginger Sesquiterpene, Activates Phase II Drug Metabolizing Enzymes. FEBS Lett. 2004, 572, 245–250. [Google Scholar] [CrossRef]
- Wani, N.A.; Zhang, B.; Teng, K.Y.; Barajas, J.M.; Motiwala, T.; Hu, P.; Yu, L.; Brüschweiler, R.; Ghoshal, K.; Jacob, S.T. Reprograming of Glucose Metabolism by Zerumbone Suppresses Hepatocarcinogenesis. Mol. Cancer Res. 2018, 16, 256–268. [Google Scholar] [CrossRef]
- Samad, N.; Abdul, A.B.; Rahman, H.S.; Rasedee, A.; Tengku Ibrahim, T.A.; Keon, Y.S. Zerumbone Suppresses Angiogenesis in HepG2 Cells through Inhibition of Matrix Metalloproteinase-9, Vascular Endothelial Growth Factor, and Vascular Endothelial Growth Factor Receptor Expressions. Pharmacogn. Mag. 2018, 13, 731–736. [Google Scholar]
- Lv, T.; Zhang, W.; Han, X. Zerumbone Suppresses the Potential of Growth and Metastasis in Hepatoma HepG2 Cells via the MAPK Signaling Pathway. Oncol. Lett. 2018, 15, 7603–7610. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Cui, P.; Zhao, Q.; Tan, B.B.; Zhang, Z.D.; Liu, Y.; Jia, N. Zerumbone Induces Gastric Cancer Cells Apoptosis: Involving Cyclophilin, A. Biomed. Pharmacother. 2016, 83, 740–745. [Google Scholar] [CrossRef]
- Han, J.; Bae, S.Y.; Oh, S.J.; Lee, J.; Lee, J.H.; Lee, H.C.; Lee, S.K.; Kil, W.H.; Kim, S.W.; Nam, S.J.; et al. Zerumbone Suppresses IL-1β-induced Cell Migration and Invasion by Inhibiting IL-8 and MMP-3 Expression in Human Triple-negative Breast Cancer Cells. Phytother. Res. 2014, 28, 1654–1660. [Google Scholar] [CrossRef]
- Jeon, M.; Han, J.; Nam, S.J.; Lee, J.E.; Kim, S. Elevated IL-1β Expression Induces Invasiveness of Triple Negative Breast Cancer Cells and is Suppressed by Zerumbone. Chem. Biol. Interact. 2016, 258, 126–133. [Google Scholar] [CrossRef]
- Fatima, A.; Abdul, A.B.; Abdullah, R.; Karjiban, R.A.; Lee, V.S. Binding Mode Analysis of Zerumbone to Key Signal Proteins in the Tumor Necrosis Factor Pathway. Int. J. Mol. Sci. 2015, 16, 2747–2766. [Google Scholar] [CrossRef]
- Sung, B.; Jhurani, S.; Ahn, K.S.; Mastuo, Y.; Yi, T.; Guha, S.; Liu, M.; Aggarwal, B.B. Zerumbone Down-regulates Chemokine Receptor CXCR4 Expression Leading to Inhibition of CXCL12-induced Invasion of Breast and Pancreatic Tumor Cells. Cancer Res. 2008, 68, 8938–8944. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sakao, K.; Singh, S.V. Notch2 Activation is Protective Against Anticancer Effects of Zerumbone in Human Breast Cancer Cells. Breast Cancer Res. Treat. 2014, 146, 543–555. [Google Scholar] [CrossRef]
- Kim, S.; Kil, W.H.; Lee, J.; Oh, S.J.; Han, J.; Jeon, M.; Jung, T.; Lee, S.K.; Bae, S.Y.; Lee, H.C.; et al. Zerumbone Suppresses EGF-induced CD44 Expression through the Inhibition of STAT3 in Breast Cancer Cells. Oncol. Rep. 2014, 32, 2666–2672. [Google Scholar] [CrossRef]
- Sehrawat, A.; Arlotti, J.A.; Murakami, A.; Singh, S.V. Zerumbone Causes Bax- and Bak-mediated Apoptosis in Human Breast Cancer Cells and Inhibits Orthotopic Xenograft Growth in vivo. Breast Cancer Res. Treat. 2012, 136, 429–441. [Google Scholar] [CrossRef]
- Shin, J.W.; Ohnishi, K.; Murakami, A.; Lee, J.S.; Kundu, J.K.; Na, H.K.; Ohigashi, H.; Surh, Y.J. Zerumbone Induces Heme Oxygenase-1 Expression in Mouse Skin and Cultured Murine Epidermal Cells through Activation of Nrf2. Cancer Prev. Res. 2011, 4, 860–870. [Google Scholar] [CrossRef]
- Yang, H.L.; Lee, C.L.; Korivi, M.; Liao, J.W.; Rajendran, P.; Wu, J.J.; Hseu, Y.C. Zerumbone Protects Human Skin Keratinocytes Against UVA-irradiated Damages through Nrf2 Induction. Biochem. Pharmacol. 2018, 148, 130–146. [Google Scholar] [CrossRef]
- Wang, S.D.; Wang, Z.H.; Yan, H.Q.; Ren, M.Y.; Gao, S.Q.; Zhang, G.Q. Chemotherapeutic Effect of Zerumbone on Melanoma Cells Through Mitochondria-mediated Pathways. Clin. Exp. Dermatol. 2016, 41, 858–863. [Google Scholar] [CrossRef]
- Yan, H.; Ren, M.Y.; Wang, Z.X.; Feng, S.J.; Li, S.; Cheng, Y.; Hu, C.X.; Gao, S.Q.; Zhang, G.Q. Zerumbone Inhibits Melanoma Cell Proliferation and Migration by Altering Mitochondrial Functions. Oncol. Lett. 2017, 13, 2397–2402. [Google Scholar] [CrossRef]
- Ashraf, S.M.; Sebastian, J.; Rathinasamy, K. Zerumbone, a Cyclic Sesquiterpene, Exerts Antimitotic Activity in HeLa Cells through Tubulin Binding and Exhibits Synergistic Activity with Vinblastine and Paclitaxel. Cell Prolif. 2019, 52. [Google Scholar] [CrossRef]
- Chan, M.L.; Liang, J.W.; Hsu, L.C.; Chang, W.L.; Lee, S.S.; Guh, J.H. Zerumbone, a Ginger Sesquiterpene, Induces Apoptosis and Autophagy in Human Hormone-refractory Prostate Cancers Through Tubulin Binding and Crosstalk Between Endoplasmic Reticulum Stress and Mitochondrial Insult. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 1223–1236. [Google Scholar] [CrossRef]
- Park, J.H.; Park, G.M.; Kim, J.K. Zerumbone, Sesquiterpene Photochemical from Ginger, Inhibits Angiogenesis. Korean J. Physiol. Pharmacol. 2015, 19, 335–340. [Google Scholar] [CrossRef]
- Kang, C.G.; Lee, H.J.; Kim, S.H.; Lee, E.O. Zerumbone Suppresses Osteopontin-Induced Cell Invasion Through Inhibiting the FAK/AKT/ROCK Pathway in Human Non-Small Cell Lung Cancer A549 Cells. J. Nat. Prod. 2016, 79, 156–160. [Google Scholar] [CrossRef]
- Abdelwahab, S.I.; Abdul, A.B.; Zain, Z.N.; Hadi, A.H. Zerumbone Inhibits Interleukin-6 and Induces Apoptosis and Cell Cycle Arrest in Ovarian and Cervical Cancer Cells. Int. Immunopharmacol. 2012, 12, 594–602. [Google Scholar] [CrossRef]
- Jorvig, J.E.; Chakraborty, A. Zerumbone Inhibits Growth of Hormone Refractory Prostate Cancer Cells by Inhibiting JAK2/STAT3 Pathway and Increases Paclitaxel Sensitivity. Anticancer Drugs. 2015, 26, 160–166. [Google Scholar] [CrossRef]
- Sun, Y.; Sheng, Q.; Cheng, Y.; Xu, Y.; Han, Y.; Wang, J.; Shi, L.; Zhao, H.; Du, C. Zerumbone Induces Apoptosis in Human Renal Cell Carcinoma via Gli-1/Bcl-2 Pathway. Pharmazie. 2013, 68, 141–145. [Google Scholar]
- Shanmugam, M.K.; Rajendran, P.; Li, F.; Kim, C.; Sikka, S.; Siveen, K.S.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Abrogation of STAT3 Signaling Cascade by Zerumbone Inhibits Proliferation and Induces Apoptosis in Renal Cell Carcinoma Xenograft Mouse Model. Mol. Carcinog. 2015, 54, 971–985. [Google Scholar] [CrossRef]
- Zainal, N.S.; Gan, C.P.; Lau, B.F.; Yee, P.S.; Tiong, K.H.; Abdul Rahman, Z.A.; Patel, V.; Cheong, S.C. Zerumbone Targets the CXCR4-RhoA and PI3K-mTOR Signaling Axis to Reduce Motility and Proliferation of Oral Cancer Cells. Phytomedicine 2018, 15, 33–41. [Google Scholar] [CrossRef]
- Ma, S.; Lei, Y.; Zhang, L.; Wang, J. Effects of Zerumbone on Proliferation and Apoptosis of Esophageal Cancer Cells and on P53 and Bcl-2 Expression Levels. Oncol. Lett. 2018, 16, 4379–4383. [Google Scholar] [CrossRef]
- Wang, M.; Niu, J.; Gao, L.; Gao, Y.; Gao, S. Zerumbone Inhibits Migration in ESCC via Promoting Rac1 Ubiquitination. Biomed. Pharmacother. 2019, 109, 2447–2455. [Google Scholar] [CrossRef]
- Hu, Z.; Zeng, Q.; Zhang, B.; Liu, H.; Wang, W. Promotion of p53 Expression and Reactive Oxidative Stress Production is Involved in Zerumbone-induced Cisplatin Sensitization of Non-small Cell Lung Cancer Cells. Biochimie 2014, 107, 257–262. [Google Scholar] [CrossRef]
- Weng, H.Y.; Hsu, M.J.; Wang, C.C.; Chen, B.C.; Hong, C.Y.; Chen, M.C.; Chiu, W.T.; Lin, C.H. Zerumbone Suppresses IKKα, Akt, and FOXO1 Activation, Resulting in Apoptosis of GBM 8401. Cells J. Biomed. Sci. 2012, 19. [Google Scholar] [CrossRef]
- Stoner, G.D. Ginger: Is it Ready for Prime Time? Cancer Prev. Res. 2013, 6, 257–262. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechner, J.F.; Stoner, G.D. Gingers and Their Purified Components as Cancer Chemopreventative Agents. Molecules 2019, 24, 2859. https://doi.org/10.3390/molecules24162859
Lechner JF, Stoner GD. Gingers and Their Purified Components as Cancer Chemopreventative Agents. Molecules. 2019; 24(16):2859. https://doi.org/10.3390/molecules24162859
Chicago/Turabian StyleLechner, John F., and Gary D. Stoner. 2019. "Gingers and Their Purified Components as Cancer Chemopreventative Agents" Molecules 24, no. 16: 2859. https://doi.org/10.3390/molecules24162859
APA StyleLechner, J. F., & Stoner, G. D. (2019). Gingers and Their Purified Components as Cancer Chemopreventative Agents. Molecules, 24(16), 2859. https://doi.org/10.3390/molecules24162859